

Учение о биологическом прогрессе и регрессе, их критерии.
Идея о направленном характере эволюции, идущей по пути прогресса, т.е. по пути совершенствования организмов от простого к сложному, от низших форм жизни к высшим, общепринята. Однако не следует думать, что прогресс - единственный путь эволюционных изменений, в действительности это лишь одно из возможных направлений эволюции.
Современные представления о направлениях эволюции основываются на работах отечественных биологов А.Н. Северцова и И.И. Шмальгаузена и американского палеонтолога Дж.Г. Симпсона. А.Н. Северцов выделил понятия биологического прогресса и регресса.
Биологический прогресс означает победу вида или другой систематической группы в борьбе за существование. Признаками биологического прогресса являются увеличение численности особей данной систематической группы, расширение ее ареала и распадение на подчиненные систематические группы. Все три признака биологического прогресса связаны друг с другом. Увеличение численности особей заставляет вид (или любую другую систематическую группу) расширять границы ареала, заселять новые места обитания, что приводит к образованию новых популяций, подвидов, видов.
Биологическому прогрессу противостоит биологический регресс . Он характеризуется обратными признаками: снижением численности особей, сужением ареала, постепенным или быстрым уменьшением популяционного и видового многообразия группы. Биологический регресс может привести вид к вымиранию. Общая причина биологического регресса - отставание в темпах эволюции группы от скорости изменений внешней среды. Быстрое изменение окружающей среды, вызванное деятельностью человека, ведет к увеличению числа видов переходящих в состояние биологического регресса и обреченных на вымирание (если не сохранится приемлемая для них среда).
Движущие силы эволюции действуют непрерывно, в результате чего совершенствуются приспособления. Однако под влиянием хозяйственной деятельности человека среда обитания живых существ изменяется значительно быстрее, чем формируются приспособления. Это приводит к сокращению численности, сужению ареалов и угрозе вымирания ряда видов. Небывалое по скорости наступление человека на природу ведет к столь быстрым изменениям среды, что виды, еще несколько десятков лет назад находившиеся в состоянии биологического прогресса, не поспевая за изменениями, претерпевают биологический регресс. Так, распашка степей ведет к резкому снижению численности целинных видов: сусликов, сурков, степных пищух, журавлей, степных орлов. Минимальные шансы на выживание имеют крупные млекопитающие (особенно хищники), представленные небольшим числом особей, которые живут на небольшой территории, медленно размножаются и не мирятся с соседством человека. Таковы, например, тигр, гепард, леопард, барс, белый медведь. Напротив, биологического прогресса достигают мелкие плодовитые животные, широко распространенные и легко уживающиеся с человеком. К ним из млекопитающих в первую очередь относится типично городское животное - крыса пасюк, из птиц - ворона, из насекомых - рыжий таракан.
Человек на строго научной основе должен активно заботиться о восстановлении численности, расширении ареалов редких и исчезающих видов. Только биологический прогресс гарантирует видам будущее.
ВОПРОС №3 Свиной цепень.
тип Plathelminthes (Плоские черви)
кл. Cestoidae (Ленточн)
отр. Cyclophyllidae (Цепни)
cem. Taenidae
в. Taenia solhim
Морфология.
Взрослая особь свиного цепня представляет собой червя с лентовидным телом белого цвета, размерами от 1,5 до 6 м. Головка цепня микроскопических размеров (2-3 мм), имеет венчик крючьев и 4 присоски. За головкой расположена шейка, а за ней — членики, имеющие квадратную форму.
Цикл развития.
В кишечнике свиньи из яйца развивается личинка, названная онкосферой, представляющая собой шарик из множества клеток с шестью хитиновыми крючками на нем.
Активно работая этими крючками, личинки проникают в кровь и переносятся в мышцы и другие органы.
Там яйца преобразуются в личиночные пузырьки — цистицерки (финны). Это особая стадия развития всех ленточных червей, ее еще называют пузырчатой глистой. Внешне финна свиного цепня выглядит как пузырек размером с горошину, наполненный жидкостью.
При внимательном рассмотрении можно заметить на финне небольшую ямку, внутри которой спрятаны неразвившиеся присоски и крючья. Это будущая головка червя (сколекс), втянутая внутрь финны.
В этот момент рост и развитие финны приостанавливаются. Для дальнейшего преобразования финне необходимо попасть в кишечник человека.
Если мясо зараженного животного не подвергнется достаточной тепловой обработке и попадет в организм человека, личинки активизируются, и это станет началом развития взрослой особи гельминта.
В кишечнике человека финна выворачивает наружу втянутую до этого головку. Присосками и крючьями она крепко цепляется к стенкам кишечника, и солитер начинает наращивать от головки новые членики.
Бывает, что в организме человека паразитирует не один цепень. Известен случай, когда при проведении курса лечения у одного пациента отошло 104 головки свиных цепней и множество члеников общей длиной 128 м.
Патогенное действие.
Симптомы заболевания бывают самыми разными: тошнота, рвота, понос, отсутствие аппетита.
Лабораторная диагностика. При диагностике недостаточно обнаружить яйца гельминта в фекалиях, так как онкосферы бычьего и свиного цепней нельзя отличить друг от друга. Необходимо обнаружить и исследовать зрелые членики.
Профилактика.
Цистицеркоз. Пути заражения. Профилактика.
Человек может служить и промежуточным хозяином для свиного цепня, когда заражение происходит от другого человека. Цистицерки, образовавшиеся в органах человека, вызывают цистицеркоз, который может проявляться в судорожных припадках и слепоте.
БИЛЕТ №12
Реализация биологической информации в клетке.
Транскрипция - первая стадия реализации генетической информации в клетке. В ходе процесса образуются молекулы мРНК, служащие матрицей для синтеза белков, а также транспортные, рибосомальные и другие виды молекул РНК, выполняющие структурные, адапторные и каталитические функции
Транскрипция у эукариотов происходит в ядре. В основе механизма транскрипции лежит тот же структурный .принцип комплементарного спаривания оснований в молекуле РНК (G ≡ C, A=U и Т=А). ДНК служит только матрицей и в ходе транскрипции не изменяется. Рибонукле-озидтрифосфаты (ЦТФ, ГТФ, АТФ, УТФ) -субстраты и источники энергии, необходимые для протекания полимеразной реакции, образования 3',5'-фосфодиэфирной связи между рибонуклеозидмонофосфатами.
Синтез молекул РНК начинается в определённых последовательностях (сайтах) ДНК, которые называют промоторы, и завершается в терминирующих участках (сайты терминации). Участок ДНК, ограниченный промотором и сайтом терминации, представляет собой единицу транскрипции - транскриптон. У эукариотов в состав транскриптона, как правило, входит один ген у прокариотов несколько. В каждом транскриптоне присутствует неинформативная зона; она содержит специфические последовательности нуклеотидов, с которыми взаимодействуют регуляторные транскрипционные факторы.
Транскрипционые факторы - белки, взаимодействующие с определёнными регуляторными сайтами и ускоряющие или замедляющие процесс транскрипции. Соотношение информативной и неинформативной частей в транскриптонах эукариотов составляет в среднем 1:9 (у прокариотов 9:1).
Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками ДНК. Разделение ДНК на множество транскриптонов позволяет осуществлять с разной активностью индивидуальное считывание (транскрипцию) разных генов.
В каждом транскриптоне транскрибируется только одна из двух цепей ДНК, которая называется матричной, вторая, комплементарная ей цепь, называется кодирующей. Синтез цепи РНК идёт от 5'- к З'-концу, при этом матричная цепь ДНК всегда антипараллельна синтезируемой нуклеиновой кислоте (рис. 4-28).
Транскрипция не связана с фазами клеточного цикла; она может ускоряться и замедляться в зависимости от потребности клетки или организма в определённом белке.
РНК-полимеразы
Биосинтез РНК осуществляется ДНК-зависимыми РНК-полимеразами. В ядрах эукариотов обнаружены 3 специализированные РНК-полимеразы: РНК-полимераза I, синтезирующая пре-рРНК; РНК-полимераза II, ответственная за синтез пре-мРНК; РНК-полимераза III, синтезирующая пре-тРНК. РНК-полимеразы - олигомерные ферменты, состоящие из нескольких субъединиц - 2α, β, β', σ. Субъединица о (сигма) выполняет регуляторную функцию, это один из факторов инициации транскрипции, РНК-полимеразы I, II, III, узнающие разные промоторы, содержат разные по строению субъединицы σ.
А. Стадии транскрипции
В процессе транскрипции различают 3 стадии: инициацию, элонгацию и терминацию.
Инициация
Активация промотора происходит с помощью большого белка - ТАТА-фактора, называемого так потому, что он взаимодействует со специфической последовательностью нуклеотидов промотора - ТАТААА- (ТАТА-бокс) (рис. 4-29).
Присоединение ТАТА-фактора облегчает взаимодействие промотора с РНК-полимеразой. Факторы инициации вызывают изменение кон-формации РНК-полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, т.е. образуется транскрипционная вилка, в которой матрица доступна для инициации синтеза цепи РНК (рис. 4-30).
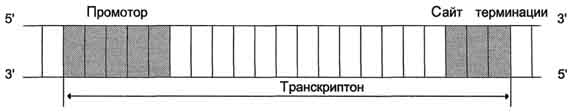
Рис. 4-27. Строение транскриптона.

Рис. 4-28. Транскрипция РНК на матричный цепи ДНК. Синтез РНК всегда происходит в направлении 5' → 3'.
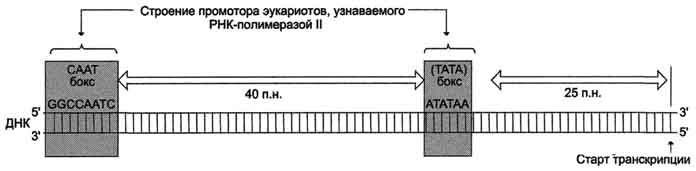
Рис. 4-29. Строение промотора эукариотов. Промоторные элементы - специфические последовательности нуклеотидов, характерные для любого промотора, связывающего РНК-полимеразу. Первый промоторный элемент - последовательность АТАТАА- (ТАТА-бокс) отделён от сайта начала транскрипции приблизительно на 25 пар нуклеотидов (п.н.). На расстоянии примерно 40 (иногда до 120) п.н. от него располагается последовательность GGCCAATC- (СААТ-бокс).
После того как синтезирован олигонуклеотид из 8-10 нуклеотидных остатков, σ-субъединица отделяется от РНК-полимеразы, а вместо неё к молекуле фермента присоединяются несколько факторов элонгации.
Элонгация
Факторы элонгации повышают активность РНК-полимеразы и облегчают расхождение цепей ДНК. Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. На стадии элонгации, в области транскрипционной
вилки, одновременно разделены примерно 18 нуклеотидных пар ДНК. Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади - восстановление двойной спирали ДНК.
Терминация
Раскручивание двойной спирали ДНК в области сайта терминации делает его доступным для фактора терминации. Завершается синтез РНК в
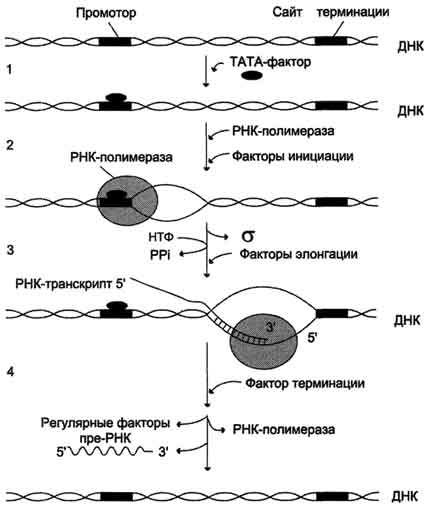
Рис. 4-30. Стадии транскрипции. 1 - присоединение ТАТА-фактора к промотору. Чтобы промотор был узнан РНК-полимера-зой, необходимо образование транскрипционного комплекса ТАТА-фактор/ТАТА-бокс (промотор). ТАТА-фактор остаётся связанным с ТАТА-боксом во время транскрипции, это облегчает использование промотора многими молекулами РНК-полимеразы; 2 - образование транскрипционной вилки; 3 - элонгация; 4.- терминация.
строго определенных участках матрицы - терминаторах (сайты терминации). Фактор терминации облегчает отделение первичного транскрипта (пре-мРНК), комплементарного матрице, и РНК-полимеразы от матрицы. РНК-полимераза может вступить в следующий цикл транскрипции после присоединения субъединицы σ.
Все образовавшиеся РНК непосредственно после трансляции не способны, функционировать, так как они синтезируются в виде молекул-предшественников: пре-р, пре-т и пре-м РНК. Чтобы начать работать, пре-РНК должны подвергнуться процессингу (созреванию). Под процессингом понимают совокупность биохимических реакций, при которых пре-РНК укорачиваются, подвергаются химическим модификациям, в результате которых образуются зрелые РНК. Процессинг т- и рРНК осуществляется по одинаковому плану. В основном вырезаются лишние фрагменты с 3'- и 5'-концов и химически модифицируются азотистые основания.
Процессинг мРНК гораздо сложнее. Интересно, что мРНК прокариот (бактерий) процессингу не подвергаются вообще — это единственные РНК, способные работать сразу после синтеза. У эукариот Процессинг мРНК осуществляется многоступенчато.
С появлением методов, позволяющих изучать первичную структуру молекул мРНК в цитоплазме и последовательность нуклеотидов кодирующей её геномной ДНК, было установлено, что они не комплементарны, а длина гена в несколько раз больше "зрелой" мРНК. Последовательности нуклеотидов, присутствующие в ДНК, но не входящие в состав зрелой мРНК, были названы некодирующими, или интроны, а последовательности, присутствующие в мРНК, - кодирующими, или экзоны. Таким образом, первичный транскрипт - строго комплементарная матрице нуклеиновая кислота (пре-мРНК), содержащая как экзоны, так и интроны. Длина интронов варьирует от 80 до 1000 нуклеотидов. Последовательности интронов "вырезаются" из первичного транскрипта, концы экзонов соединяются друг с другом. Такую модификацию РНК называют "сплайсинг" (от англ, to splice - сращивать). Сплайсинг происходит в ядре, в цитоплазму поступает уже "зрелая" мРНК.
Гены эукариотов содержат больше интронов, чем экзонов, поэтому очень длинные молекулы пре-мРНК (около 5000 нуклеотидов) после сплайсинга превращаются в более короткие молекулы цитоплазматической мРНК (от 500 до 3000 нуклеотидов).
Процесс "вырезания" интронов протекает при участии малых ядерных рибонуклеопротеинов (мяРНП). В состав мяРНП входит малая ядерная РНК (мяРНК), нуклеотидная цепь которой связана с белковым остовом, состоящим из нескольких протомеров. В сплайсинге принимают участие различные мяРНП (рис. 4-32).
166
Нуклеотидные последовательности нитронов функционально неактивны. Но на 5'- и З'-концах они имеют высокоспецифические последовательности - AGGU- и GAGG- соответственно (сайты сплайсинга), которые обеспечивают их удаление из молекулы пре-мРНК. Изменение структуры этих последовательностей влияет на процесс сплайсинга.
На первой стадии процесса мяРНП связываются со специфическими последовательностями первичного транскрипта (сайты сплайсинга), далее к ним присоединяются другие мяРНП. При формировании структуры сплайсосомы 3'-конец одного экзона сближается с 5'-концом следующего экзона. Сплайсосома катализирует реакцию расщепления 3',5'-фосфодиэфирной связи на границе экзона с интроном. Последовательность интрона удаляется, а два экзона соединяются. Образование 3',5'-фосфодиэфирной связи между двумя экзонами катализируют мяРНК (малые ядерные РНК), входящие в структуру сплайсосомы. В результате сплайсинга из первичных транскриптов мРНК образуются молекулы "зрелой" мРНК.