

39. Нейронные сети. Однонаправленные многослойные нейронные сети
Многослойные однонаправленные сети
Такие сети называются также сетями прямого распространения, или многослойными персептронами.
Сети этого типа состоят из нескольких слоев нейронов: входного слоя, выходного и нескольких «скрытых слоев». На рис. изображена сеть, у которой К слоев. Нейроны каждого слоя не связаны между собой. Выходной сигнал с каждого нейрона поступает на входы всех нейронов следующего слоя. Нейроны входного слоя не осуществляют преобразования входных сигналов, их функция заключается в распределении этих сигналов между нейронами первого скрытого слоя.
Функционирование многослойной сети осуществляется следующим образом: входной сигнал, подаваемый на сеть, поступает на нейроны входного слоя, проходит по очереди через все слои и снимается с выходов нейронов выходного слоя. По мере распространения сигнала по сети он претерпевает ряд преобразований, которые зависят от его начального значения, от преобразующих функций и величин весов связей.
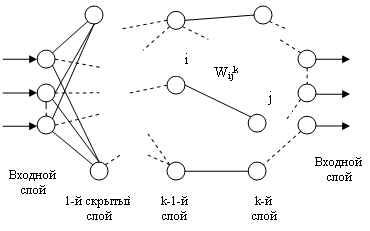
Многослойный перцептрон представляет собой классический пример однонаправленной нейронной сети, т. е. нейронной сети, в которой отсутствуют как обратные воздействия сигналов выходных нейронов на вход сети, так и межсоединения между нейронами одного и того же слоя.
48 Генетический алгоритм. Мутация, ее роль в алгоритме. Механизмы реализации процесса мутации.
Мутация
Принято считать что репродукция в генетическом алгоритме является основны поисковым механизмом, а мутация позмоляет лишь внести в процесс некоторую случайность, что понижает вероятность застревания алгоритма в локальных решениях. Однако кроме этой метафоры существует множество других. Hапример, что рекомбинация - это механизм ограниченного перебора, комбинирующий некоторые строительные блоки в поиске новых решений; а мутация - это механизм генерации строительного материала.
Как бы то нибыло, с уверенностью можно сказать лишь одно - опыты показывают необходимость присутствия обоих механизмов в алгоритме. При этом В большенстве экспериментов оператор мутации применяется к потомку с заданной вероятностью, причем величина вероятности выбирается давольно малой - порядка 0,01.
Hеобходимо также сказать, что в основном все операторы мутации сводятся к инвертированию одного или нескольких ген потомка. Это справедливо только для бинарного кодирования хромосом. В случае других способов кодирования мутация гена требует либо случайного выбора из заданного множества вариантов (случай алфавитного кодирования) либо специальных алгоритмов.
Одноточечная мутация
В этом варианте оператора мутации в потомке случайно выбирается один ген и мутируется. По своему опыту могу сказать что это крайне не эффективный подход, т.к. во многих задачах изменение одного гена не позволяет выйти решению из локального оптимума. Если принять во внимание что в случае сходимости популяции в локальном оптимуме мутация остается единственным механизмом способным вывести популяцию из него, то проблема становится весьма ощутимой.
Плотность мутации
Стратегия мутации с использованием понятия "плотности" заключается в мутировании каждого гена потомка с заданной вероятностью. Таким образом, кроме вероятности применения мутации к самому потомку используется еще вероятность применения мутации к каждому его гену, величину которой выбирают с таким расчетом, чтобы в среднем мутировало от 1 до 10 процентов ген.
Инцест
В нескольких своих работах я предложил использовать стратегию инцеста (хотя возможно не я первый её придумал :) как механизм самоадаптации оператора мутации. Она заключается в том, что "плотность мутации" (вероятность мутации каждого гена) определяется для каждого потомка на основании генетической близости его родителей. Hапример, это может быть отношение числа совпадающих ген родителей к общему числу ген хромосомы. Это приводит к очень интересному эффекту - при высоком разнообразии генофонда популяции (первые шаги ГА) последствия мутации будут минимальными, что позволяет репродукции работать без стороннего вмешательства. В случае же понижения разнообразия, что возникает в основном при застревании алгоритма в локальном оптимуме, последствия мутации становятся более ощутимыми, а при полном схождении популяции алгоритм просто становится стахостическим, что увеличивает вероятность выхода популяции из локального оптимума.