

ВВЕДЕНИЕ
Поджелудочная железа, по сути дела, представляет собой два разных органа, объединенных в единую морфологическую структуру. Ее ацинарная часть выполняет экзокринную функцию, секретируя в просвет двенадцатиперстной кишки ферменты и ионы, необходимые для процессов пищеварения. Эндокринная часть железы состоит из 1-2 млн. островков Лангерганса, на долю которых приходится 1—2% всей массы поджелудочной железы. Островки состоят из клеток разных типов.
Таблица 1. Типы клеток в островках Лангерганса
тип клетки |
относительное содержание, % |
образующийся гормон |
А |
25 |
Глюкагон |
В |
70 |
Инсулин |
D |
Мене 5 |
Соматостатин |
F |
следовые количества |
Панкреатический полипептид |
Островковый аппарат поджелудочной железы секретирует по крайней мере четые гормона: инсулин, глюкагон, соматостатин и панкреатический полипептид. Эти гормоны высвобождаются в панкреатическую вену, впадающую в воротную вену, что имеет очень важное значение, поскольку для инсулина и глюкагона печень служит главной мишенью. Основная роль этих двух гормонов сводится к регуляции углеводного обмена, однако они оказывают влияние и на многие другие процессы. Соматостатин впервые идентифицирован в гипоталамусе как гормон, подавляющий секрецию гормона роста. Однако в поджелудочной железе его концентрация выше, чем в гипоталамусе. Этот гормон участвует в локальной регуляции секреции инсулина и глюкагона. Панкреатический полипептид влияет на желудочно-кишечную секрецию.
ИНСУЛИН
Инсулин - гормон поджелудочной железы; усиливает проникновение глюкозы в клетки организме и способствует усвоению сахара клетками организма, снижая таким образом его содержание в крови. Инсулин теряет активность при температуре тела ниже 32*С. Инсулин способствует переходу калия и фосфора из внеклеточной жидкости внутрь клетки.
Инсулин оказывает на клетку плейотропное действие, охватывающее регуляторные эффекты, которые условно делят на две категории: метаболические и ростостимулирующие . Имеются данные, позволяющие полагать, что и другие пептиды инсулинового семейства также способны оказывать подобные влияния на клетку.
О том, сколь велика роль инсулина в углеводном , белковом и липидном обмене , яснее всего свидетельствуют последствия инсулиновой недостаточности у человека. Основным признаком сахарного диабета является гипергликемия , развивающаяся в результате 1)пониженного проникновения глюкозы в клетки, 2)снижения утилизации глюкозы различными тканями и 3)повышения образования глюкозы (глюконеогенеза) в печени.
В отсутствие инсулина снижается биосинтез белка , что отчасти объясняется уменьшением транспорта аминокислот в мышцы (аминокислоты служат субстратами для глюконеогенеза). Таким образом, инсулиновая недостаточность у человека сопровождается отрицательным азотным балансом . Характерное для этой ситуации отсутствие антилиполитического действия инсулина, равно как и его липогенного действия, приводит к тому, что содержание жирных кислот в плазме возрастает . Когда оно достигает уровня, превышающего способность печени окислять жирные кислоты до СO2, в крови накапливаются бета-гидроксимальная и ацетоуксусная кислоты (кетоз). Вначале организм компенсирует накопление этих органических кислот увеличением количества выдыхаемого СO2. Однако если развитие кетоза не сдерживается введением инсулина, то развивается тяжелый метаболический ацидоз и больной погибает от диабетической комы. Механизм инсулиновой недостаточности схематически представлен:
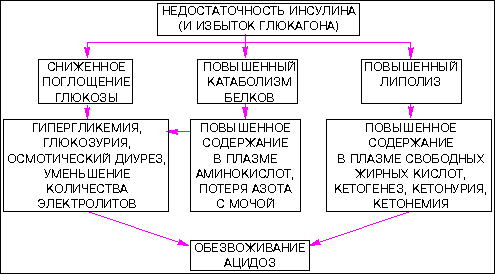
Инсулин синтезируется в поджелудочной железе B-клетками островков Лангерганса, и его поступление в кровь увеличивается при гипергликемии. Концентрация этого гормона в крови изменяется параллельно концентрации глюкозы; введение его быстро вызывает гипогликемию. К веществам, вызывающим секрецию инсулина, относятся аминокислоты, свободные жирные кислоты, кетоновые тела, глюкагон, секретин; адреналин и норадреналин, наоборот, блокируют его секрецию. Инсулин быстро вызывает увеличение поглощения глюкозы жировой тканью и мышцами за счет ускорения транспорта глюкозы через клеточные мембраны путем транслокации переносчиков глюкозы из цитоплазмы в плазматическую мембрану. Однако инсулин не оказывает прямого действия на проникновение глюкозы в клетки печени; это согласуется с данными о том, что скорость метаболизма глюкозы клетками печени не лимитируется скоростью ее прохождения через клеточные мембраны. Инсулин, однако, действует опосредованно, влияя на активность ферментов, участвующих в гликолизе и гликогенолизе.
Инсулин: структура
Инсулин - двухцепочечный полипептид, - состоит из 51 аминокислотного остатка и имеет молекулярную массу около 6000. Его короткая цепь А состоит из 21 остатка аминокислот, а более длинная цепь В - из 30 остатков.
Обе цепи связаны между собой двумя дисульфидными мостиками (7-7 и 20-19).
Цепи инсулина кодируются одним геном, который обусловливает синтез первичного крупного одноцепочечного белка-предшественника инсулина, из которого затем выщепляются обе цепи инсулина.
Первичная химическая структура инсулина была расшифрована Сэнгером (1953), а пространственная структура инсулина - Д.Ходжкин (1972).
Анализ функциональной роли структурных элементов его молекулы показал, что каждая цепь в отдельности биологической активностью не обладает. Для проявления биологической активности необходима жесткая пространственная конфигурация его двухцепочечной молекулы, закрепленная двумя дисульфидными мостиками, которые соединяют цепи А и В, а также внутренним дисульфидным мостиком 6-11 цепи А.
С-концы цепей А и В находятся вблизи друг от друга, образуя гидрофобный центр молекулы инсулина, богатый ароматическими аминокислотами.
Этот центр имеет первостепенное значение для проявления биологической активности инсулина.
Предполагается, что для связывания клеточными рецепторами специфическое значение имеет 24-29-я С-концевая часть цепи В.
Инсулин: влияние на транспорт глюкозы через мембрану
Внутриклеточная концентрация свободной глюкозы значительно ниже ее внеклеточной концентрации. Большинство имеющихся данных свидетельствует о том, что скорость транспорта глюкозы через плазматическую мембрану мышечных и жировых клеток определяет интенсивность фосфорилирования глюкозы и ее дальнейший метаболизм. D-глюкоза и другие сахара с аналогичной конфигурацией по С1-С3 (галактоза, D-ксилоза и L-арабиноза) проникают в клетки путем облегченной диффузии, опосредованной переносчиком. Во многих клетках инсулин усиливает этот процесс, что обусловливается увеличением числа переносчиков (Мmax-эффект), а не повышением сродства связывания (Км-эффект). Согласно имеющимся данным, в жировых клетках это происходит путем мобилизации переносчиков глюкозы из неактивного их пула в аппарате Гольджи с дальнейшим направлением их к активному участку плазматической мембраны. Такая транслокация переносчиков - процесс, зависимый от температуры и энергии и независимый от синтеза белков.
Печеночные клетки представляют собой важное исключение из этой схемы. Инсулин не стимулирует облегченной диффузии глюкозы в гепатоциты, но усиливает ее приток косвенным путем, индуцируя глюкокиназу - фермент, превращающий глюкозу в глюкозо-6-фосфат. В результате быстро протекающего фосфорилирования концентрация свободной глюкозы в гепатоцитах поддерживается на очень низком уровне, что способствует проникновению глюкозы в клетки путем простой диффузии по градиенту концентрации.
Инсулин способствует также проникновению в клетки аминокислот (особенно в мышечные клетки) и стимулирует перемещение К+, Са2+, нуклеозидов и органического фосфата. Эти эффекты зависят от влияния инсулина на поступление в клетку глюкозы.