

Глава 10
Кровь и перенос газов
129
Мы видели, что при акклиматизации к высоте концентрация гемоглобина превышает это значение. Очевидно, особые обстоятельства приводят к нарушению оптимальной концентрации гемоглобина. И, по-видимому, не случайно у самого мелкого млекопитающего — землеройки — концентрация гемоглобина в крови выше, чем у других млекопитающих, — около 170 г на 1 л крови (Ulrich, Bartels, 1963). У мелких летучих мышей концентрация гемоглобина даже еще выше: отмечены значения до 244 г гемоглобина на 1 л крови (Jurgens et al., 1981).
Таким образом, мы пришли к выводу, что концентрация гемоглобина в крови млекопитающих не зависит от их размеров, однако могут возникать особые физиологические требования, которые приводят у некоторых животных к превышению оптимальной концентрации.
Объем крови
Общий объем крови у большинства млекопитающих составляет от 60 до 70 см3 на 1 кг массы тела; таким образом, относительный объем крови не зависит от размеров тела. Уравнение для объема крови (VK, мл) относительно массы тела (Мт, кг) имеет следующий вид: Ук = 65,6 MTl>02 (Stahl, 1967). Показатель степени при массе почти не отличается от 1,0; т. е. кровь млекопитающих составляет в общем постоянную часть массы тела.
Следует отметить, что из этого общего правила имеются исключения, и эти исключения весьма значительны. В частности, у некоторых ныряющих млекопитающих относительный объем крови гораздо больше. Очевидно, это объясняется тем, что кровь представляет собой наиболее важное хранилище кислорода, которым снабжается мозг во время ныряния.
Поскольку у наземных млекопитающих объем крови составляет постоянную долю массы тела, а концентрация гемоглобина в крови не зависит от размеров тела, общее количество гемоглобина в теле животного (произведение объема крови на концентрацию гемоглобина) будет постоянной долей массы тела (опять-таки за исключением ныряющих млекопитающих).
Размеры эритроцитов
Эритроциты млекопитающих по форме напоминают круглые двояковогнутые диски. Они легко деформируются и могут проходить через капилляры, диаметр которых меньше диаметра эритроцита, тем не менее диаметр этих клеток дает неплохое приближение для величины необходимого просвета капилляров. Если мы сравним размеры эритроцитов различных млекопитающих, то обнаружим, возможно, неожиданный факт — их диа-
метр оказывается довольно однообразным и не зависит от размеров тела (табл. 10.1).
Таблица 10.1, Диаметр эритроцитов разных млекопитающих; более чем у 100 видов дополнительно изученных млекопитающих эритроциты по диаметру не выходят за пределы, указанные в таблице. (Altaian, Dittmer, 1961.)
Вид |
Диаметр эритроцита, мкм |
Вид |
Диаметр эритроците, ним |
Вид |
Диаметр эритроцита, мкм |
Землеройка Мышь Крыса Собака |
7,5 6,6 6,8 7,1 |
Овца Человек Корова |
4,8 7,5 5,9 |
Лошадь Слон Горбатый кит |
5,5 9,2 8,2 |
Мы располагаем обширнейшей информацией относительно эритроцитов (Altman, Ditmer, 1961). В списке эритроцитов от 115 видов млекопитающих большая часть значений (свыше 100) укладывалась в границы от 5 до 8 мкм без какой-либо корреляции с размерами тела. У самого мелкого млекопитающего — землеройки — и одного из самых крупных —■ горбатого кита — размеры эритроцитов практически одинаковы — 7,5 и 8,2 мкм соответственно. Ни у одного из млекопитающих диаметр эритроцитов не превышает 10 мкм, и только у нескольких (овца, коза, олень) диаметр меньше 5 мкм.
Выводы, которые можно сделать из приведенных данных, таковы: 1) капилляры у самых мелких и самых крупных млекопитающих имеют фактически одинаковый диаметр и 2) высокая интенсивность метаболизма у самых мелких млекопитающих, очевидно, не требует какой-либо специфической адаптации эритроцитов (т. е. расстояние диффузии внутри эритроцита не оказывает влияния на интенсивность метаболизма).
У других позвоночных эритроциты овальные, а не дискообразные. У птиц короткий диаметр овального эритроцита почти равен диаметру эритроцита млекопитающих — от 6 до 7 мкм, а длинный — почти в два раза больше. Эритроциты рептилий также овальны и несколько больше, чем у птиц, — короткий и длинный диаметры составляют около 12 и 20 мкм. Эритроциты рыб имеют приблизительно такие же размеры, но в сравнении с ними эритроциты амфибий кажутся гигантскими. У бесхвостых амфибий эритроциты достигают 20—25 мкм длины, тогда как у хвостатых — 35 мкм, а у некоторых видов даже превышают 50 мкм (Szarski, Czopek, 1966). Мы пока еще не понимаем, каково функциональное значение более крупных по сравнению с другими позвоночными эритроцитов амфибий, поэтому и не будем обсуждать этот вопрос более подробно.
9 Шмидт-Ниельсен
130
Кровь и перенос
газов
Поглощение кислорода и его перенос
Обратимое связывание кислорода с гемоглобином имеет значение для двух процессов: 1) поглощения кислорода в органах дыхания и 2) высвобождения кислорода в метаболически активных тканях. Высокая степень сродства гемоглобина к кислороду благоприятствует активному поглощению кислорода дыхательной поверхностью. Вместе с тем высвобождению кислорода в тканях благоприятствует низкая степень сродства, так что кислород легко высвобождается из связи с гемоглобином. Таким образом, степень сродства гемоглобина к кислороду должна быть в некотором роде компромиссной между этими двумя требованиями. Можно ли найти какую-либо связь между сродством гемоглобина к кислороду и размерами тела, которая бы соответствовала высокой интенсивности метаболизма у мелких млекопитающих?
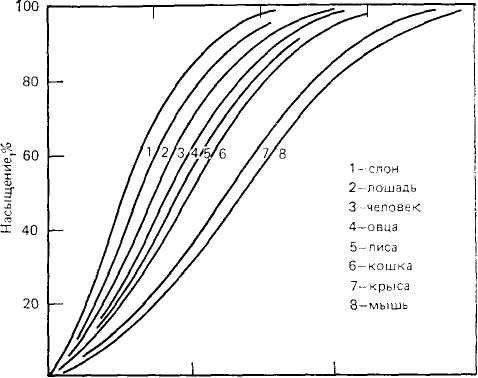
Сродство гемоглобина к кислороду. Обычный и очень удобный способ описания связи между гемоглобином и кислородом— это так называемая кривая кислородной диссоциации. Эта кривая показывает для любой конкретной пробы крови степень насыщения гемоглобина кислородом при любой данной его концентрации. Ряд типичных кривых кислородной диссоциации для крови млекопитающих приведен на рис. 10.2.
Форма этих кривых почти одинакова, но их расположение в обычной системе координат до некоторой степени различно. Если кривая диссоциации расположена левее, то гемоглобин связывает больше кислорода при любом данном давлении кислорода. Иными словами, кривая, расположенная левее, указывает на более высокое сродство к кислороду. Если кривая расположена правее, то при любом данном давлении кислорода его связывается меньше, т. е. сродство гемоглобина к кислороду низкое, и он легче высвобождается. Таким образом, расположение кривой диссоциации левее или правее указывает соответственно на высокое или низкое сродство к кислороду.
В гл. 9 было показано, что интенсивность вентиляции легких тесно связана с потребностью в кислороде, причем парциальное давление кислорода в легких не зависит от размеров. У всех млекопитающих при нормальных условиях парциальное давление кислорода в альвеолярном воздухе очень близко к 100 мм рт. ст. (при нормальном атмосферном давлении). Взглянув на кривые диссоциации, приведенные на рис. 10.2, мы видим, что при Ро2, равном 100 мм рт. ст., кровь всех млекопитающих насыщена фактически на 100%, т. е. гемоглобин принял полную нагрузку кислорода и больше связывать его не в состоянии, даже если давление кислорода повысится.
13
Парциальное давление кислорода, кПа 4 ' 8 12
131
0 40 80 120
Парциальное давление кислорода, мм рт.ст.
Рис. 10.2. Кривые кислородной диссоциации для крови млекопитающих разных видов. У мелких млекопитающих сродство гемоглобина к кислороду ниже. Это облегчает отдачу кислорода в тканях для поддержания высокого уровня метаболизма у мелких животных. (Из Schmidt-Nielsen, 1972.)
Мы можем заключить, что, хотя кровь млекопитающих и различается по сродству к кислороду, это не оказывает заметного влияния на количество кислорода, которое она может забрать при нормальном парциальном давлении кислорода в альвеолах.
Таким образом, кажется, что у млекопитающих сродство гемоглобина к кислороду не имеет существенного значения для скорости его поглощения в легких. Это, однако, неверно для животных, у которых обычно Рой в альвеолярном воздухе не достигает 100 мм рт. ст. Так, у животных, которые обитают на больших высотах, например у южноамериканской ламы, сродство крови к кислороду гораздо выше величины, обычной для млекопитающих, т. е. кривая кислородной диссоциации для них расположена значительно левее границ, обычных для млекопитающих (Hall el at., 1936). Таким образом, высокое сродство крови к кислороду у ламы облегчает поглощение кислорода при низком атмосферном давлении на больших высотах. Это высокое сродство к кислороду и его влияние на степень насыщения кислородом артериальной крови ламы подтверждено достаточно полно (например, Banchero et al., 1971). Сдвиг влево характерен для небольшого числа животных, обитающих на большой вы-
132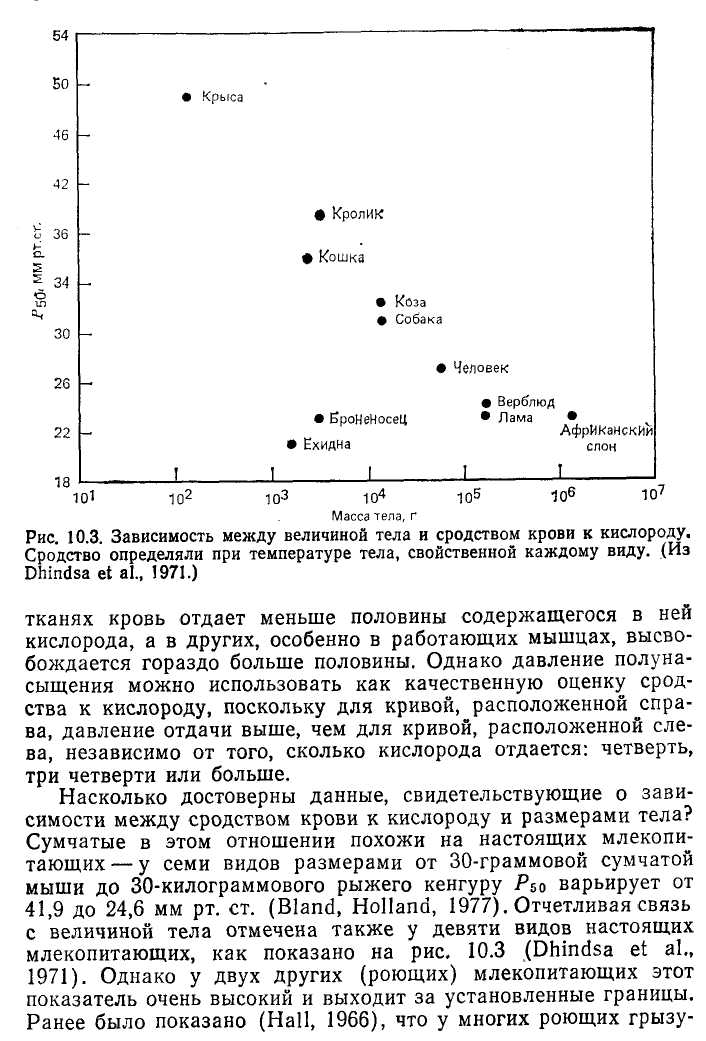
соте. Когда равнинные животные, в том числе человек, акклиматизируются к высоте, их кривые диссоциации по существу не меняются или даже могут слегка сдвигаться в противоположную сторону (Lenfant, Sullivan, 1971).
У некоторых роющих грызунов кровь характеризуется исключительно высоким сродством к кислороду (кривая диссоциации расположена слева). Среди них у луговых собачек (грызуны) сродство крови к кислороду особенно высокое, фактически такое же высокое, как у ламы. Луговые собачки роют норы глубиной до 5 м (Sheets et al., 1971), и, по-видимому, иногда этим животным приходится дышать в своих плохо вентилируемых норах воздухом с низким парциальным давлением кислорода; при этом высокое сродство крови к кислороду облегчает его поглощение. И действительно, из 17 изученных видов грызунов лишь собачки способны выжить при самом низком парциальном давлении кислорода (Hall, 1966).
В заключение еще раз отметим, что у огромного большинства млекопитающих расположение кривой кислородной диссоциации (сродство к кислороду) не имеет большого значения для поглощения кислорода в легких при обычном парциальном давлении кислорода. Однако из этого правила имеются исключения, которые можно объяснить особенностями местообитаний животных, такими, как большая высота или подземный образ жизни. В этих случаях поглощение кислорода обеспечивается более высоким, чем обычно, сродством крови к кислороду.
Высвобождение кислорода в тканях. Серия кривых диссоциации на рис. 10.2 показывает связь между сродством к кислороду и размерами тела млекопитающих. Кривые кислородной диссоциации для мелких животных расположены правее, что указывает на более низкое сродство крови к кислороду и более легкое его высвобождение, тогда как у более крупных животных кривые расположены левее, т. е. сродство крови к кислороду у них выше.
Мы видели, что расположение кривой диссоциации, вероятно, не связано с поглощением кислорода в легких, за исключением некоторых особых случаев (высокогорные и роющие животные). В таком случае существует ли более значимая связь между легкостью высвобождения кислорода и размерами тела?
С этой точки зрения в расположении кривых диссоциации можно найти больше смысла. Однако нам необходим удобный способ, чтобы выразить положение кривой диссоциации и сродство к кислороду. Обычно эти переменные выражаются через величину давления полунасыщения (Р50), которое иногда называют «давлением отдачи» для кислорода. Это давление (Pso) не является точным давлением отдачи О2, так как в некоторых
134