

Иннервация скелетной мышечной ткани
Скелетные мышцы обладают эфферентной (двигательной) и афферентной (чувствительной) иннервацией.
Эфферентная иннервация скелетных мышц обеспечивается нервными волокнами (аксонами а-мотонейронов), образующими на мышечных волокнах специализированные нервно-мышечные окончания (нервно-мышечные синапсы, или моторные бляшки), которые осуществляют передачу возбуждения с нервного волокна на мышечное. Один мотонейрон может иннервировать различное количество мышечных волокон.
Двигательная единица (ДЕ) представляет собой совокупность мотонейрона и иннервируемых им мышечных волокон. Количество мышечных волокон, входящих в одну ДЕ, минимально в мелких мышцах, осуществляющих точные и тонкие движения. В глазных мышцах человека ДЕ включает от 2-6 до 13-20 мышечных волокон, в мышцах кисти – от 10-25 до 100-300, в мышцах туловища – 1500-2000. Управление мышечной активностью обеспечивается как изменением частоты активации ДЕ, так и вовлечением различного их числа в процесс сокращения.
Мышечные волокна, образующие одну ДЕ, обладают одинаковыми механическими свойствами, гистохимическими характеристиками и относятся к одному типу, однако рассеяны по обширной территории мышцы.
Афферентная иннервация скелетных мышц обеспечивается нервно-мышечными веретенами - рецепторами растяжения волокон поперечнополосатых мышц, которые представляют собой сложные инкапсулированные нервные окончания, состоящие из веточек нервных волокон, оплетающих особые тонкие (интрафузальные) мышечные волокна, заключенные в тончайшую соединительнотканную капсулу. Остальные мышечные волокна назваются экстрафузальными.
Сердечная мышечная ткань
Сердечная мышечная ткань встречается только в мышечной оболочке сердца (миокарде) и устьях связанных с ним крупных сосудов. Ее клетки (сердечные миоциты, или кардиомиоциты) составляют лишь 30-40% общего числа клеток сердца, но образуют 70-90% его массы. Основным функциональным свойством сердечной мышечной ткани служит способность к спонтанным ритмическим сокращениям, на активность которых влияют гормоны и нервная система (симпатическая и парасимпатическая).
|
|
 А
А
 Б
Б
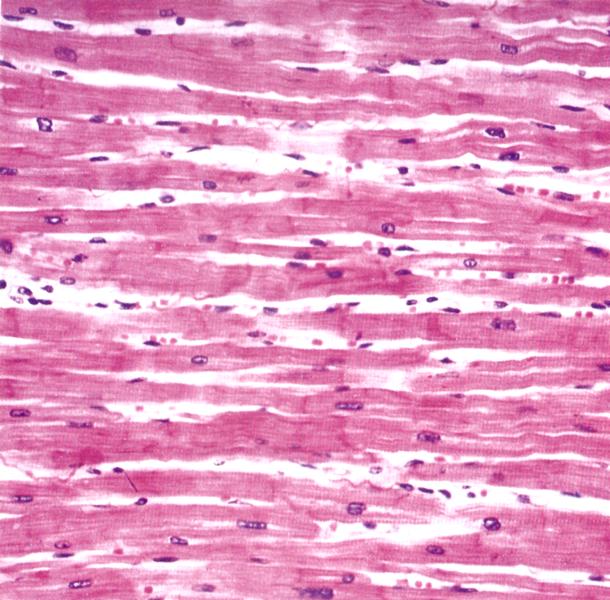
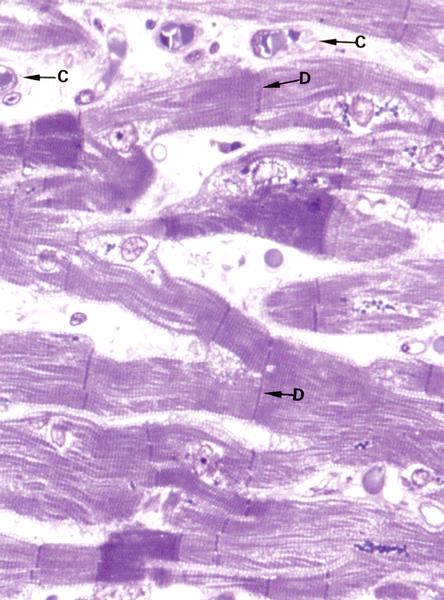
Рис. 7.12. На электронной микрофотографии изображен продольный срез миокарда А(х 198), Б(х 640). Ядра в миоцитах расположены центрально, синтициальное строение миокарда (образуется симпласт) – миоциты соединяются между собой с помощью вставочных дисков (D). Между миоцитами расположены прослойки соединительной ткани, в которой много капилляров (С).
Функциональная морфология сердечной мышечной ткани
Сердечная мышечная ткань образована клетками - кардиомиоцитами (сердечными миоцитами), связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих функциональных волокон (рис. 7.12).
Кардиомиоциты - цилиндрические или ветвящиеся клетки, более крупные в желудочках, где их длина составляет 100-150 мкм, а диаметр - 10-20 мкм. В предсердиях они обычно имеют неправильную форму и меньшие размеры (длина - 40-70 мкм, диаметр - 5-6 мкм). Кардиомиоциты содержат одно или два ядра и саркоплазму, покрыты сарколеммой, которая снаружи окружена базальной мембраной.
Саркоплазма кардиомиоцитов содержит органеллы и включения, которые образуют следующие аппараты: 1) сократительный, 2) передачи возбуждения (с сарколеммы на сократительный аппарат), 3) опорный, 4) энергетический, 5) синтетический, 6) лизосомальный (аппарат внутриклеточного переваривания).
Сократительный аппарат сильно развит в сократительных (рабочих) кардиомиоцитах (в особенности, в желудочковых), которых он занимает до 50-70% объема клетки. Фагоструктура сходна с таковой поперечно-полосатой мышечной ткани. Механизм мышечного сокращения этих видов мышечной ткани также сходны.
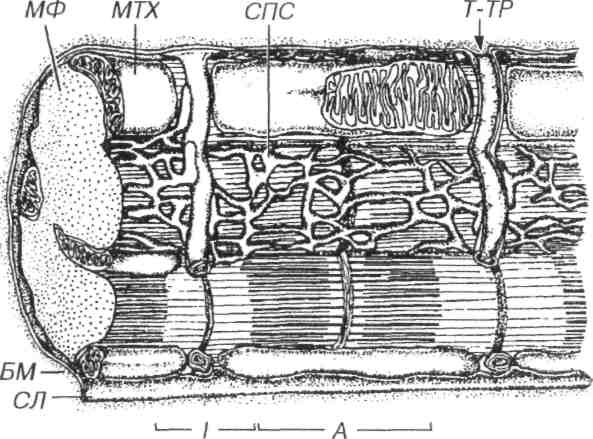
Рис. 7.13. Саркотубулярная система кардиомиоцита. Саркотубулярная система представлена саркоплазматической сетью (СПС), не образующей терминальных цистерн, и широкими Т-трубочками (Т-ТР).
Аппарат передачи возбуждения (саркотубулярная система) в кардиомиоцитах в целом сходен с таковым в скелетных мышечных волоконах, однако он обладает рядом особенностей (рис. 7.13).
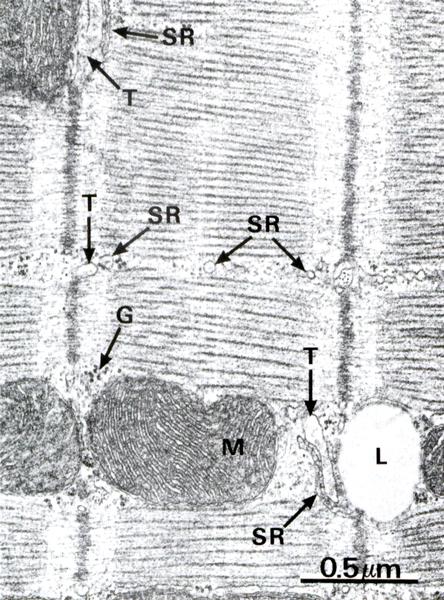
Саркоплазматическая сеть развита слабее, чем в скелетном мышечном волокне, менее активно накапливает Са2+, не образует терминальных цистерн. Во время расслабления она выделяет ионы Са2+ в саркоплазму с низкой скоростью, что обеспечивает автоматизм кардиомиоцитов (рис. 7.14).
|
|
 А
А
 Б
Б
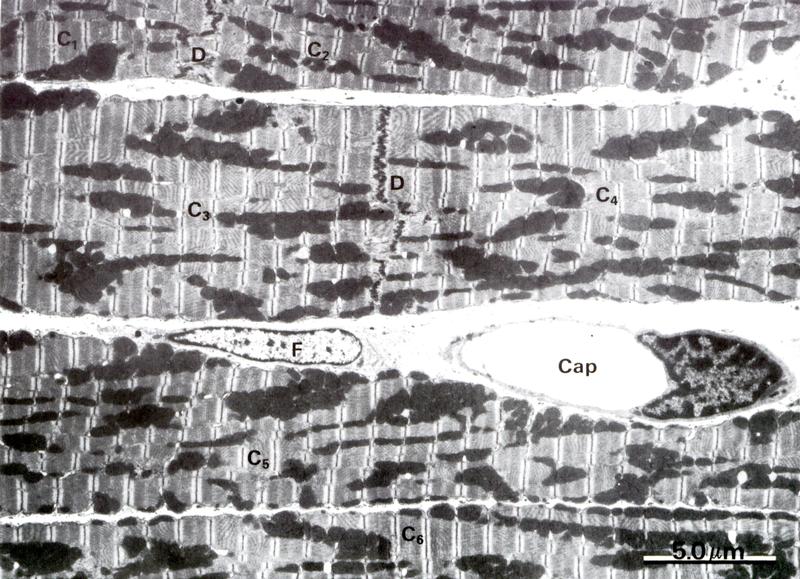
Рис. 7.14. На электронной микрофотографии изображен: А (х5.000) – участок миокарда состоящий из 6 миоцитов (С1-С6). Эти клетки соединяются между собой вставочным диском D. В поле зрения в миоцитах нет ядер. Между миоцитами – рыхлая соединительная ткань, в которой много капилляров (Сар), фибробластов (F); Б (х38.000) ультраструктура саркомера миокарда схожа с таковой поперечно-полосатой мышечной тканью. Обратить внимание на строение А и J дисков СПР (SR), тубулярной системы (Т), расположение митохондрий. В миоците много липидных капель (L) и гранул гликогена (G).
Поперечные (Т-) трубочки - широкие, содержат компоненты базальной мембраны, вместе с элементами саркоплазматической сети образуют диады (включают одну Т-трубочку и одну цистерну сети), которые располагаются в области Z-линий (рис. 7.14).
Опорный аппарат кардиомиоцитов представлен элементами цитоскелета, обеспечивающими упорядоченное расположение миофиламентов и миофибрилл внутри волокна, а также базальной мембраной и сарколеммой. Его структурная и биохимическая организация сходна с таковой в волокнах скелетной мышечной ткани. В кардиомиоцитах его особенностью служит то, что элементы цитоскелета связаны с особыми межклеточными соединениями – вставочными дисками.
Вставочные диски осуществляют связь кардиомиоцитов друг с другом. Под световым микроскопом они имеют вид поперечных прямых или зигзагообразных полосок, пересекающих функциональное волокно сердечной мышечной ткани (рис. 7.15).
Энергетический аппарат кардиомиоцитов представлен митохондриями и включениями, расщепление которых обеспечивает получение энергии. Митохондрии кардиомиоцитов обладают также свойством накапливать кальций в высоких концентрациях.
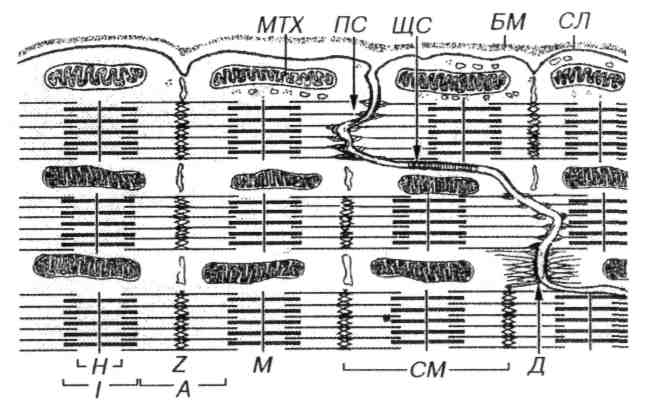
Рис. 7.15. Ультраструктурная организация области вставочного диска кардиомиоцитов. В поперечных участках вставочного диска соседние кардиомиоциты образуют многочисленные интердигитации, связанные контактами типа десмосом (Д) актиновые филаменты прикрепляются к поперечным участкам сарколеммы вставочного диска в участке полоски слипания (ПС). На сарколемме продольных участков вставочного диска располагаются щелевые соединения (ЩС). БМ – базальная мембрана, СЛ – сарколемма, МТХ – митохондрия. Обозначения компонентов саркомера СМ) – те же, что на рис. 4.
Синтетический аппарат кардиомиоцитов Синтетический аппарат включает свободные рибосомы и полисомы, цистерны грЭПС. и элементы комплекса Гольджи, которые располагаются в саркоплазме преимущественно у полюсов ядра.
Лизосомальный аппарат (аппарат внутриклеточного переваривания) кардиомиоцитов хорошо развит, что отражает высокую скорость обновления их структурных компонентов. Он включает эндосомы, лизосомы и остаточные тельца. Собственно лизосомы располагаются у полюсов ядра и занимают до 10% объема саркоплазмы. Остаточные тельца в виде липофусциновых гранул очень многочисленны (при старении могут составлять до 20% сухой массы миокарда).