

Транспортные атф-азы.
Исследования показали, что в биологических мембранах имеется несколько разновидностей ионных насосов – специальных систем интегральных белков – транспортных АТФаз. Перенос ионов транспортными АТФазами происходит вследствие сопряжения процессов транспорта с химическими реакциями, за счёт энергии метаболизма клеток. В настоящее время известны 3 типа электрогенных ионных насосов: Na, K-АТФаза, Ca-АТФаза и протонная помпа – Н-АТФаза (рис.9).
Первичный активный транспорт.
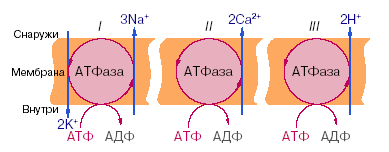
Рис.9. Активный перенос ионов транспортными АТФазами. I – схема K, Na насоса в клеточной плазматической мембране; II – схема кальциевого насоса в мембране саркоплазматического ретикулума; III – схема протонного насоса во внутренней мембране митохондрий
При работе Na, K-АТФазы за счёт энергии, освобождающейся при гидролизе каждой молекулы АТФ, в клетку переносятся 2 иона калия и одновременно из клетки выкачиваются 3 иона натрия. Таким образом, создаётся повышенная по сравнению с межклеточной средой концентрация в клетке ионов калия и пониженная – ионов натрия, что имеет огромное физиологическое значение. Ca-АТФаза обеспечивает активный перенос 2 ионов кальция, протонная помпа – 2-3 протонов на одну молекулу АТФ.
Признаки «бионасоса»:
Движение против градиента электрохимического потенциала.
поток вещества сопряжён с гидролизом АТФ (или другого источника энергии).
асимметрия транспортной машины.
насос in vitro способен гидролизовать АТФ только в присутствии тех ионов, которые он переносит in vivo.
при встраивании насоса в искусственную среду он способен сохранять селективность.
Вторичный (сопряжённый) активный транспорт.
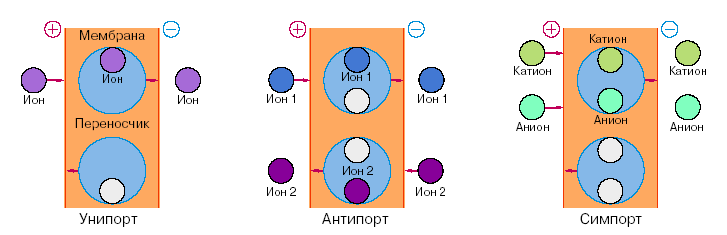
нипорт |
Антипорт |
Симпорт |
(пассивный) (К+ - валиномицин) |
Na+ и Н+ (бактерии) (нигерицин К+ и Н+) 2Na+ и Са++ |
(2Na+ и АК, ГЛ) |
Вторичный (или сопряжённый) активный транспорт ионов наблюдается в том случае, если в результате первичного транспорта была создана разность концентраций или потенциалов на мембране, которая затем приводит к перемещению рассматриваемых ионов. В настоящее время различают три схемы вторичного транспорта (рис.10). однонапрвленный перенос иона в комплексе со специфическим переносчиком, который получил название унипорта. При этом через мембрану переносится заряд либо комплексом, если молекула переносчика электронейтральна, либо пустым переносчиком, если перенос обеспечивается заряженным переносчиком. Движущей силой ионного транспорта служит электрическое поле, т.е. разность потенциалов на мембране, созданная в результате первичного транспорта ионов. Второй механизм сопряжённого переноса ионов – это антипорт, при котором перенос одного иона через мембрану происходит одновременно с переносом второго иона того же заряда в противоположном направлении. Совместный однонаправленный перенос ионов с участием «двухместного» переносчика называется симпортом. Считается, что по схеме симпорта происходит доставка и накопление клетками аминокислот. Из схемы симпорта следует, что этот процесс должен сопровождаться значительным увеличением осмотической активности, поскольку в одном цикле через мембрану переносятся две частицы в одном направлении.
При использовании АТФ образуются тетрамеры из 4 пар и субъединиц. При использовании ГТФ протомеры.
Характеристика Na, K-АТФазы. Na, K-АТФаза была одним из первых мембранных ферментов, выделенных из клеточных мембран в активном состоянии. Известно, что этот фермент состоит из двух полипептидных цепей. Большая из них (-субъединица) содержит -спиральные участки, которые как «колонны» пересекают мембрану 10 раз. Фрагменты полипептидной цепи, расположенные между 2-й и 3-й, а также между 4-й и 5-й -спиралями, являются достаточно протяжёнными. Первый из них содержит центр для переносимых ионов (ион-связывающий центр), а второй – центр, в котором осуществляется связывание и гидролиз АТФ (активный центр, обращённый в цитоплазму. Меньший по размерам полипептид (-субъединица), который встраивается в мембрану одновременно с -субъединицей, непосредственно не участвует в работе фермента, но обеспечивает правильную ориентацию его в мембране. (+)-комплекс представляет собой функциональную единицу Na,K-АТФазы (протомер), способную гидролизовать АТФ, связывать ионы и переносить их через мембрану. Таким образом, Na,K-АТФаза работает как молекулярная машина, перекачивающая через мембрану ионы натрия и калия, поэтому её называют Na,K-насосом.
Механизм действия.
Рассмотрим гидролитический цикл
фермента, начиная со стадии взаимодействия
белка с Na+. «Натриевая
конформация» (Na-конформер)
обозначается E1.
«Калиевая конформация» (К-конформер)
обозначается как E2.
взаимодействие Na-конформера
с АТФ приводит к образованию
фермент-субстратного комплекса, в
котором натрий активирует перенос
терминальной фосфатной группы АТФ на
карбоксил аспарагинового остатка белка,
приводящий к образованию фосфофермента,
обозначаемого
![]() ,
где
,
где
![]() – молекула белка-фермента, а Ф –
фосфатная группа. Гидролиз связи
– молекула белка-фермента, а Ф –
фосфатная группа. Гидролиз связи
![]() активируется K+.
Схематически весь процесс выглядит
следующим образом:
активируется K+.
Схематически весь процесс выглядит
следующим образом:

Основные стадии можно описать следующим образом:
Когда фермент находится в состоянии E1 он способен взаимодействовать с Na+ и АТФ с внутренней стороны мембраны. В результате фосфорелирования молекулы образуется , а АДФ высвобождается из активного центра и возвращается в цитоплазму.
Фосфорелированный белок переходит в состояние, при котором ионы натрия не способны высвобождаться ни с внутренней, ни с внешней стороны мембраны, они недоступны для обмена.
Переход фермнта в следующую стадию существенно активируется ионами Mg2+. Т.о., магний играет двойную роль в работе фермента – в комплексе с АТФ обеспечивает образование истинного субстрата реакции, а всвободном состоянии является активатором одной из стадий реакции. Хотя специальных центров связывания Mg2+ на молекуле фермента не обнаружено, его эффеккт очень важен – он заключается в ускорении перехода фосфорилированного фермента из E1 в E2. эта стадия отражает молекулярные перемещения отдельных частей белковой глобулы, связанные с непосредственным переносом Na+ через мембрану Na,K-насосом. Таким образом, перенос Na+ через мембрану осуществляется синхронно с конформационным переходом
 .
Вследствие этого перехода центр
связывания ионов становится более
гидрофобным, и ионы натрия диссоциируют
от фермента по другую сторону мембраны,
где с этим же центром связываются ионы
калия.
.
Вследствие этого перехода центр
связывания ионов становится более
гидрофобным, и ионы натрия диссоциируют
от фермента по другую сторону мембраны,
где с этим же центром связываются ионы
калия.K+ подвергается такой же окклюзии, что и Na+, в ходе которой осуществляется его перенос через мембрану.
Комплекс отличается тем, что Ф становится доступна для атаки молекулой воды. Происходит гидролиз и высвобождение неорганического фосфата во внутриклеточную среду.
После этого ионы K+ также диссоциируют от центров связывания, высвобождаясь в цитоплазму. Их место занимает Na+. Последняя стадия цикла одновременно подготавливает фермент для начала нового цикла.
Так происходит активный транспорт Na+ из клеток и K+ – в клетки, а энергия АТФ тратится на «оплату» перехода фермента из одной конформации в другую. Т.о., в ходе ферментативного процесса перенос ионов натрия и калия осуществляется одним и тем же ионным центром фермента, последовательно изменяющим своё сродство к переносимым ионам одновременно с изменением конформации Na, K-АТФазы. Центр связывания ионов расположен в петле между 2-й и 3-й -субъединицами фермента. Взаимодействие ионов с этими центрами обеспечивается координационными связями с атомами кислорода карбоксильных групп аспарагиновой и глутаминовой аминокислот. Кислород способен осуществлять координационные взаимодействия с лигандами, образуя решётку вдух типов. В одном случае образуется более рыхлая и доступная для молекул воды структура и ионный центр может связать 3 иона натрия, в другом реализуется более плотная упаковка и связываются два иона калия. Этим и обьясняется тот факт, что при гидролизе 1 молекулы АТФ фермент обменивает 3 иона натрия на два иона калия.
Комформационная перестройка, претерпеваемая белком при переходе , обеспечивает изменение конформации петли и связанную с этим перестройку ионного центра. Таким образом, перемещение петли (вместе со связанными ионами) внутрь мембраны обеспечивает сначала окклюзию переносимых монов, а затем их высвобождение по другую сторону мембраны.
Схема
рабочей части натрий-калиевого насоса:![]()

Энергетика цикла.
![]() ,
,
![]() ;
;
![]() ,
,
![]() ;
;
![]() ,
,
![]() .
.
Общие затраты энергии за один цикл: 2*20+3*130 = 430 мэВ.
Энергия гидролиза АТФ: 300-500 mV «КПД» 100%.
Схема кальциевого насоса:
