

6. Бластула.
У багатьох яйцеклітин ще на ранніх стадіях дроблення внутрішні кінці бластомерів розходяться і між ними виникає спочатку невелика, а потім все більша порожнина дроблення (бластоцель). У деяких типів яйцеклітин бластоцель може досягати значних розмірів. Зародок на цій стадії розвитку називається бластулою. У ході подальшого розвитку бластоцель перетворюється на первинну порожнину тіла, що є основною порожниною тіла в нижчих безхребетних. У вищих безхребетних і хребетних вона майже зовсім витісняється виникаючою пізніше вторинною порожниною тіла (целомом).
Бластоцель виконує дві функції:
- дає можливість клітинам при гаструляції мігрувати усередину зародка;
- існування бластоцелі запобігає взаємодії між клітинами, які перебувають вище і нижче неї.
Бластоцель - перший виникаючий по ходу розвитку відсік внутрішнього середовища організму, що відрізняється за іонним складом від зовнішнього середовища. Клітини стінок бластоцелі, які відгороджують її від зовнішнього середовища, утворюють між собою щільні контакти, що мають вибіркову проникливість для іонів.
На зовнішній, оберненій до зовнішнього середовища мембрані клітин розташовані іонні канали для натрію, хлору та інших іонів, що забезпечують їхній перенос по градієнтах концентрації, а на внутрішній, оберненій до бластоцелі - іонні насоси, що працюють проти градієнту. В результаті Nа+ та Cl¯ перекачуються в бластоцель, де виникає їхній надлишок, що створює в бластоцелі підвищений осмотичний тиск та забезпечує перенесення води, підвищуючи в ній тургорний тиск. Цей тиск розтягує поверхню зародка, що важливо для його наступного розвитку Надлишок натрію в бластоцелі може впливати на швидкість клітинних циклів і стимулювати експресію генів у клітинах бластули.
7. Типи бластул.
Є 5 основних типів бластул - целобластула; бластоциста; амфібластула; дискобластула; перибластула.
Целобластула (від грецького koilos – порожній) характерна для голкошкірих та ланцетника (мал. 12 А).
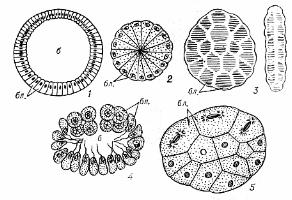
Мал. 12. Типи бластул (за Мануіловою, 1973; Токіним, 1987; Білоусовим, 1989):
1 – целобластула, поперечний розріз (морський огірок); 2 – стерробластула, поперечний розріз (ставромедуза); 3 – плакула (справа – вид збоку); 4 – стомобластула, поперечний розріз (вапняні губки); 5 – морула; бл. – бластомери.
Цей тип бластул відрізняється тим, що бластомери на анімальному та вегетативному полюсі абсолютно однакові за розміром, а тонку бластодерму формує один шар клітин. Порожнина всередині - бластоцель. Виділяють у целобластулі наступні зони: покрівля на анімальному полюсі, дно - на вегетативному, між ними знаходиться крайова або проміжна зона.
Бластула зі стінкою рівномірної товщини та дуже маленьким центрально розташованим бластоцелем (мал. 12 Б), що зустрічається в деяких кишковопорожнинних, молюсків і хробаків називається стерробластулою (від греч. sterros – твердий, щільний). Зародковий розвиток вапняних губок, деяких хробаків, асцидій приводить до утворення плакули. Вона має вигляд двошарової пластинки, утвореної однорідними клітинами. Між шарами розташовується порожнина дроблення (бластоцель). У деяких кишковопорожнинних, а також у ссавців на ранніх стадіях розвитку, бластоцель взагалі не виникає, і дроблення закінчується на стадії щільного скупчення клітин – морули (від лат. morum – шовковична ягода) (мал. 13 А).
Своєрідним типом бластули є стомобластула (від греч. stoma – рот), характерна для зародкового розвитку вапняних губок (мал. 12 В). Вона має порожнину в центрі та отвір (фіалопор) на анімальному полюсі. Джгутикоутворюючи полюси клітин обернені досередини. По закінченні дроблення стомобластула вивертається навиворіт через фіалопор (екскурвація), в результаті чого утворюється покрита джгутиками амфібластула.
Бластоциста характерна для ссавців (мал. 13 Б).
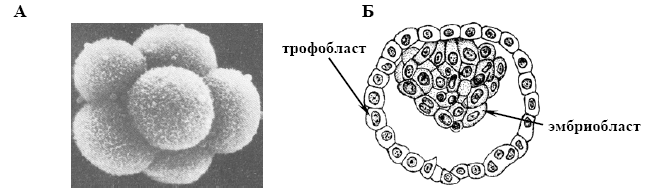
Мал. 13. Типи бластул (за Токіним, 1987 і Білоусовим, 1993):
А – морула, зовнішній вигляд (миша), Б – бластоциста, поперечний розріз (кажан).
Утворюється після проходження стадії морули. Вона представлена у вигляді пухирця (від грецьк. kystis – міхур). Причому в складі цього пухирця є дві частини. Стінку формує один шар дуже світлих клітин - трофобласт. Інша частина – темно зафарбовані клітини ембріобласту. Трофобласт забезпечує харчування, а ембріобласт йде на побудову тіла зародка.
Амфібластула характерна для амфібій і вапняних губок. На відміну від целобластули бластоцель в амфібластулі зміщена до анімального полюсу внаслідок більшої кількості жовтка на вегетативному полюсі. На анімальному полюсі знаходяться менші за розміром клітини - мікромери. В області вегетативного полюса розташовуються макромери. Як і у целобластулі, в амфібластулі розрізняють покрівлю, дно та проміжну (крайову) зону (мал. 14).
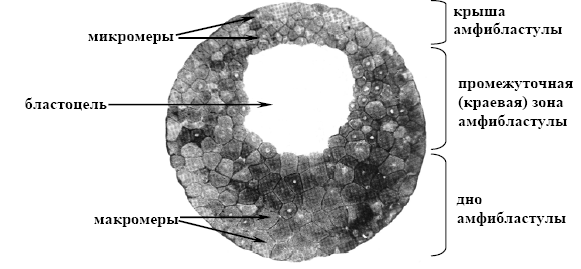
Мал. 14. Амфібластула (за Фаліним, 1957):
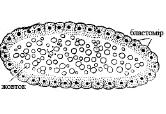
Дискобластула характерна для костистих риб, рептилій і птахів. В результаті дискоїдального дроблення утворюється одношарова бластодерма (бластодиск), з якого потім виникає 3-х або 4-х клітинний шар (мал. 15).

Мал. 15. Дискобластула: бл. – бластоміри; б – бластоціль; ж – жовток.
Бластодерма дискобластули трохи вигинається над жовтком і між ними формується підзародкова порожнина. На цій стадії чітко помітні дві області бластодиску:
світле поле (area pellucida) – відповідає тій його частині, що розташовується над порожниною;
темне поле (area opaca) – складається із клітин, розташованих по краю бластодиска та прилеглих до жовтка (мал. 16, А).
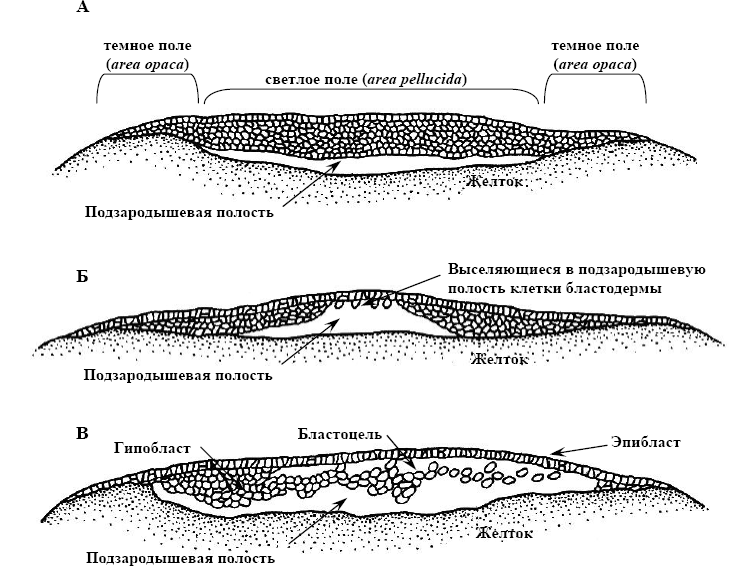
Мал. 16. Дискобластула - утворення гіпобласту в яйці птаха (за Гілбертом, 1993):
А – бластодерма, перетворена на трьох- або чотирьох шаровий клітинний пласт;
Б – початок виселення клітин бластодерми в підзародкову порожнину та формування гіпобласту; В – завершення утворення дискобластули: формування епі- і гіпобласту, блатоцелі.
До моменту відкладення яйця деякі із клітин бластодерми виселяються у підзародкову порожнину, де утворюють другий шар (мал. 16, Б). Таким чином, на даній стадії зародок складається з епібласту (зовнішній шар клітин) і розташованого під ним гіпобласту. Між ними знаходиться бластоцель (мал. 16, В).
Перібластула (мал. 17) характерна для комах і являє собою заключну стадію поверхневого дроблення. Енергіди (ядра з оточуючими їх острівцями цитоплазми) мігрують на поверхню яйця, де вони оточуються новими плазматичними мембранами. Першими зміни відбуваються у ядрах, що мігрували до заднього полюса яйця та утворили полярні клітини зародка, які дають початок статевим клітинам дорослого організму. Клітини бластодерми, що утворюється, утворюють скупчення на вентральній поверхні, позначаючи місце, де виникнуть зародкові листки (зародкова смужка). Ця область дає початок всім клітинам комахи. Інші клітини бластодерми утворюють зовнішньозародкові оболонки.
Мал. 17. Перібластула
Таким чином, дроблення являє собою сукупність процесів, результатом яких є перетворення заплідненого яйця на багатоклітинну структуру. Характерні особливості кожного типу дроблення залежать від еволюційного шляху виду та від здатності яєць забезпечити потреби зародка в поживних речовинах. Деякі тварини потребують великої кількості жовтка для того, щоб підтримати розвиток зародка до його переходу на активне харчування, тоді як інші тварини проходять у своєму розвитку стадію личинки, що харчується, і для них не треба присутності великої кількості жовтка у цитоплазмі яйця. У ссавців в результаті своєрідного типу дроблення виникають елементи зародку плаценти., що буде постачати поживними речовинами. За незвичайною різноманітністю типів дроблення ховається, однак, спільність функцій та механізмів. В усіх випадках каріокінез та цитокінез повинні бути координовані та яйце поділено на клітинні області. В процесі дроблення відновлюється характерне для соматичних клітин ядерно-плазменне співвідношення і важлива для розвитку інформація розподіляється між різними клітинними областями.
Контрольні питання:
1. Біологічна роль дроблення.
2. Відмінності дроблення від мітозу соматичних клітин.
3. Механізми дроблення.
4. Просторова організація бластомерів. Правила Гертвіга-Сакса.
5. Борозни дроблення, їхнє розташування й хід.
6. Типи дроблення. Принципи, покладені в основу їхньої класифікації.
7. Голобластичне дроблення: радіальне й спіральне.
8. Голобластичне дроблення: білатеральне й анархічне.
9. Меробластичне дроблення.
10. Бластула. Функції бластоцелю.
11. Будова целобластули (стерробластули, плакули), морули (бластоцисти), стомобластули. Для яких тварин характерні ці типи дроблення.
12. Будова амфібластули, дискобластули та перібластули. Для яких тварин характерні ці типи дроблення.
У результаті запліднення утворюється зигота - початкова стадія розвитку нового організму. Стадія зиготи триває від кількох хвилин до кількох годин. У деяких видів тварин вже в зиготі здійснюється інтенсивний синтез білка, матрицею якого на початкових стадіях є власна ІРНК яйцеклітини. У цей же час з'являється яскраво виражена двобічна симетрія. У жаби, наприклад, точка, в якій спермій проник в яйцеклітину, якраз і визначає площину симетрії зародка і положення майбутньої дорсальної губи бластопора. Встановлено, що в зиготі ссавців і людини до початку наступної стадії ембріогенезу також відбувається диференціювання і переміщення ділянок цитоплазми, що призводить до двобічної симетрії.
Наступна стадія ембріогенезу – дроблення. Дробленням називають ряд мітотичних поділів зиготи, між якими немає типової інтерфази: пресинтетичний період G1 — відсутній повністю, а cинтетичний S-період дуже короткий і починається ще у телофазі попереднього мітозу. В результаті цього дочірні клітини зиготи (бластомери) не набувають розмірів материнських клітин і з кожним поділом стають все меншими і меншими і, кінець кінцем, результат дроблення - бластула (морула) майже не відрізняється за розміром від зиготи (рис. 1.154).
Дроблення залежно від типу яйцеклітини може бути повним і неповним; рівномірним і нерівномірним; синхронним і асинхронним, але в будь-якому випадку обов'язково закінчується утворенням бластули (від лат. Ьіазіоз - зачаток, проросток).
У залежності від типу дроблення розрізняють целобластули, амфібластули, дискобластули, стер-робластули. Будова їх завжди однакова. Вони мають стінку (бластодерму), побудовану із бластомерів, і порожнину всередині, яка називається бла-стоцелем, або первинною порожниною. Стадію бла-стули проходять зародки всіх типів тварин. Дроблення зиготи людини повне, нерівномірне й асинхронне закінчується утворенням бластоцисти (стер-робластули) (рис. 1.155).
По закінченню періоду дроблення у багатоклітинних тварин настає період утворення зародкових листків - гаструляція. Гаструляція - процес розвитку з одношарового зародка багатоклітинних тварин (бластули) двошарового (гаструли), а в більшості з них - згодом і тришарового.
Гаструляція починається з утворення в бластулі круглого отвору - бластопора (рис. 1.156). При цьому бластоцель зникає, а утворюється нова порожнина - порожнина первинної кишки. Клітини зародка переміщуються, розташовуються у вигляді трьох окремих зародкових листків, або шарів, утворюючи гаструлу. Утворення зародкових листків відбувається у два етапи. Спочатку утворюється рання гаструла, яка має два зародкових листки (ектодерму й ентодерму), а потім пізня гаструла, коли формується третій зародковий листок - мезодерма (рис. 1.155). На першому етапі можливі чотири способи: інвагінація (впинання), як у ланцетника; імміграція (виселення клітин), як у кишковопорожнинних (рис. 1.157); епіболія (обростання), як у жаби, і деламінація (розщеплення), як у деяких кишковопорожнинних.
Другий етап гаструляції передбачає два варіанти: телобластичний і ентероцельний.
Обґрунтована і доведена тотожність зародкових листків у всіх систематичних групах вищих тварин. У людини перший етап гаструляції проходить переважно за типом деламінації, а другий етап - шляхом виселення клітин. На стадії гаструляції зародок імплантується (проникає) у слизову оболонку матки (рис. 1.158).
Подальші клітинні поділи, переміщення, ріст та диференціювання зародкових листків призводять до гістогенезу - утворення тканин. При цьому спостерігаються закономірності, притаманні конкретним листкам і тканинам (рис. 1.159).
Процес формування органів називається органогенезом. Гістогенез і органогенез ідуть паралельно і завершуються в основному наприкінці ембріонального періоду.
Основоположником експериментальної ембріології став наприкінці XIX століття німецький зоолог В. Ру. Він спробував з'ясувати, на якій стадії розвитку зародка визначається доля його частин. Чи може, наприклад, кожен із двох перших бластомерів розвинутись у цілий організм? З цією метою В. Ру один із двох бластомерів жаби руйнував розпеченою голкою.
Із незруйнованого (цілого) бластомера розвинувся неповноцінний зародок. Це дало привід В. Ру дійти невірного висновку про те, що вже на ранніх стадіях ембріонального розвитку доля окремих бластомерів повністю визначена. Тут потрібно відзначити, що в ході експерименту вчений залишав ушкоджений бластомер поряд з цілим (неушкодженим). Пізніше інші науковці-експериментатори провели експеримент інакше. Вони забирали ушкоджений бластомер від цілого і тоді спостерігали, як з останнього розвивався повноцінний зародок. Учень В. Ру -Г. Дріш показав: якщо яйця морського їжака помістити в морську воду, позбавлену кальцію, то у зиготі, що почала ділитися, бластомери розпадаються і потім кожен із них, за сприятливих умов, здатен сформувати повноцінний зародок. Таке явище, коли ізольований бластомер може розвинутися в повноцінний організм, отримало назву тотіпотентність {рівноспадковість), а такі бластомери назвали тотіпотентними (рис. 1.160). У подальшому, знову ж таки експериментальним шляхом, було встановлено, що у тритона зберігається така тотіпотентність до стадії 16 бластомерів, у кроля -до стадії 4 бластомерів. Про існування подібного явища у бластомерів людини вказують випадки народження двох, трьох, чотирьох і навіть семи монозиготних близнюків.
Після утворення бластули зародкові клітини втрачають тотіпотентність. Починається диференціація, тобто формування різноманітних структур і частин тіла (а згодом - тканин і органів) із відносно однорідного матеріалу зародка. Але з'ясувалося, що, незважаючи на втрату тотіпотентності і диференціації, клітини повністю зберігають генетичну інформацію. Це показали в своїх експериментах американський ботанік Стюард на моркві та англійський зоолог Д. Гердон (1964—1966), який пересаджував ядра із клітин епітелію шкіри і кишок пуголовків у яйцеклітини, з яких їх власні ядра видалялися. Багато з таких клітин розвинулись, утворивши пуголовків і навіть статевозрілих жаб (рис. 1.161).
Таким чином, можна дійти висновку, що будь-яка соматична клітина, з одного боку, являє собою диференційовану частину організму і виконує вузькоспеціалізовані функції, а з іншого боку, утримує в собі генотип всього організму. Тоді випливає питання: якщо генотип у всіх клітин однаковий, а різні клітини мають неоднакову будову і входять до складу органів, що відрізняються один від одного за морфологією і за функціями, то чим зумовлене диференціювання під час розвитку?
Щоб розібратись у цьому питанні, повернемося знову до експерименту В. Ру з двома бластомерами зародка жаби. Ру вбивав один з бластомерів, але залишав його поряд з цілим, неушкодженим, і спостерігав неповноцінний його розвиток. Тобто тотіпотентність не проявлялася. Чому? А тому, що ушкоджений бластомер впливав і перешкоджав нормальному розвитку другого цілого бластомера. Так майже 100 років тому була виявлена і доведена взаємодія частин організму в процесі розвитку. Шзшше це явище взаемного впливу одних час-тин оргашзму, що розвиваеться, на iirtni отримало назву ембрюналъно'1 шдукцп.
Першим, хто переконливо дов1в взаемодш час-тин зародка на приклад1 формування ока, був на початку XX столптя шмецький ембрюлог Г. Шпе-ман (1864-1941). Вш автор орипнально!' теорй 1нди-вщуального розвитку - теори оргашзацшних центр1в.
Рання закладка ока являе собою випинання тка-нини пром1жного мозку в так званий очний Mixyp. BiH росте у напрямку до ширноУ ектодерми, з яко'1 на Micni i'x зближення утворюеться кришталик. В експеримент1 було показано, що якщо на одному бощ зародка видалити закладку очного Mixypa, то на цьому бощ кришталик не формуеться. I, навпаки, якщо закладку очного Mixypa пересадити шд пиару в шшому Micui голови або тулуба, то тут на меяа ектодерми виникае кришталик. BiH у свою чергу, вплине на формування очноУ западини. Таким чином, розвиток одного органа зумовлюе формування наступних.
Подалыпими дослщженнями було розкрито HOBi ланки впливу м1ж частинами зародка. Г. Шпема-ном було доведено, що при нормальному розвитку спинна ектодерма гаструли перетворюеться на зачаток нервовоТ трубки, але якщо и пересадити, на-приклад, на черевну частину зародка, то з не! утво-риться шира живота. I, навпаки, якщо шюрну екто-дерму живота пересадити на спинну частину зародка, то з не\' утвориться нервова трубка (рис. 1.162).
Це свщчить про те, що на стадн ранньо'1 гаструли доля зачатюв ще не повнютю визначена i мож-ливе перевизначення подалыпого шляху розвитку. Ця стад1я ембр1онального розвитку отримала назву стадй лабшьного диференц1ювання.
Чому ж ектодерма спинно!' частини зародка дае початок нервовш трубц13i вЫма и похщними? Ек-спериментально було встановлено, що якщо на раннш стадп видалити дшянку верхньо1 "губи" бла-стопора, яка i прилягае до ектодерми на спинному бощ зародка, то нервова трубка зовс1м не утворюеться. I, навпаки, якщо верхню "губу" бласто-пора пересадити шд шюру в будь-якш дшянщ гаструли, то з навколишшх кл1тин формуеться нервова трубка. I нав1ть якщо пересадити додаткову верхню "губу" бластопора, то утвориться додат-кова нервова трубка. Так, Г. Шпеман дійшов висновку, що верхня "губа" бластопора впливає на клітини ектодерми, спрямовуючи їх розвиток на формування нервової трубки. Згодом були виявлені інші ділянки тіла зародка з аналогічним впливом на його частини. Ці ділянки були названі організаційними центрами, а їх індукований вплив - організаторами (або індукторами).
Реалізація індукції можлива тільки за умов, що клітини здатні сприймати вплив, тобто є компетентними. Компетенція клітин може змінюватися в часі (виникати й зникати). На пізніших стадіях розвитку, коли диференціація уже почалася, пересадка цієї ділянки в іншу частину зародка не змінює його розвиток, бо він вже повністю визначений. Це явище незалежної, стабільної диференціації. У розвитку органів спостерігається перехід від залежної до незалежної диференціації.
Подальші дослідження проблеми ембріональної індукції йшли в напрямку визначення природи індукторів.
Ще в 30-ті роки XX ст. американський вчений Ч. Чайлд висунув припущення про градієнт фізіологічної активності організму зародка. Було показано, що інтенсивність окиснювальних та інших метаболічних процесів поступово гальмується в напрямку від головного відділу до хвостового. Згідно з цими даними, розвиток органів у тій чи іншій частині зародка визначається інтенсивністю метаболізму. Дані узгоджуються з положеннями школи Шпемана: дійсно, найбільш інтенсивний обмін речовин у гаструли відбувається в ділянці верхньої "губи" бластопора, тобто в місці утворення нервової трубки.
X. Тидеман у 1970 році провів дослідження, які показали роль взаємовідносин органів зародка на молекулярному рівні. Він виділив із зародків курчат індуктори білкової природи. Один з них індукує структури нервової системи й органів чуття, інший -м'язову систему. Вчений довів, що білки-індуктори є вже у незаплідненому яйці, але вони знаходяться в неактивному стані через присутність інгібіторів, які стримують їх дію. Під час ембріонального розвитку змінюється іонний склад цитоплазми і зовнішнього середовища, яке оточує клітину. Це призводить до виходу інгібіторів із клітини і цим інактивація з індукторів знімається. В результаті цього індуктори входять в ядро і активують відповідні гени, що, у свою чергу, сприяє диференціюванню клітин і формуванню органів і систем. Механізми росту та морфогенезу
Морфогенез — це процес виникнення нових структур і зміна їх форми в процесі онтогенезу. Морфогенез на клітинному рівні розпочинається з гаструляції. У хордових після гаструляції закладаються осьові органи (рис. 1.163). В цей період морфологічні перебудови охоплюють весь зародок. Потім відбувається закладка і розвиток окремих органів (органогенез) (рис. 1.164). Це вже місцеві процеси, всередині кожного з яких відбувається дискретизація на окремі зачатки. Так послідовно в часі і в просторі перебігає індивідуальний розвиток, що призводить до формування організму із складною будовою і значно багатшою інформацією, ніж генетична інформація зиготи.
Морфогенез зв'язаний з багатьма процесами, починаючи з прогенезу. Поляризація яйцеклітини, овоплазматична сегрегація після запліднення, закономірно орієнтовані поділи дробіння, рух клітинних мас в процесі гаструляції і закладок різних органів, зміна пропорцій тіла - все це процеси, які мають велике значення для морфогенезу.
Таким чином, морфогенез - багаторівневий динамічний процес. Сьогодні вже багато відомо про ті перебудови, які відбуваються на внутрішньоклітинному і міжклітинному рівнях і які перетворюють хімічну енергію клітин на механічну, тобто про елементарні рушійні сили морфогенезу.
Ріст - це збільшення загальної маси в процесі розвитку, яка призводить до постійного збільшення розмірів організму.
Ріст забезпечується наступними механізмами:
1) збільшенням розмірів клітин;
2) зростанням числа клітин;
3) накопиченням неклітинної речовини, продуктів життєдіяльності клітин.
Ріст відбувається на клітинному, тканинному, органному та організмовому рівнях. Збільшення маси цілого організму віддзеркалює ріст його органів, тканин і клітин.
Розрізняють два типи росту: обмежений і необмежений. Необмежений ріст триває впродовж усього періоду онтогенезу аж до смерті (наприклад, риби). При обмеженому рості - ріст відбуваєтьсялише в певні періоди онтогенезу, а згодом уповільнюється і гальмується.
Однією з важливих характеристик росту є його диференційованість, тобто швидкість росту неоднакова в різних ділянках організму і на різних стадіях розвитку.
Швидкість загального росту людського організму також залежить від стадії розвитку (рис. 1.165). Максимальна швидкість характерна для перших чотирьох місяців ембріонального розвитку, що пояснюється інтенсивним поділом клітин. У міру росту плода кількість мітозів зменшується у всіх тканинах і після шести місяців внутрішньоутробного розвитку майже не утворюються нові м'язові і нервові клітини.
О Генетичний контроль розвитку
Одним із центральних питань медичної біології є генетичний контроль розвитку. В цілому генетичний контроль онтогенезу є очевидним, однак його механізми ще не повністю з'ясовані. Для того, щобвивчити генетику індивідуального розвитку, вчені широко використовують метод експериментальної мутації. Виявивши мутації, які змінюють онтогенез, дослідник порівнює фенотипи мутантних особин з нормальними. Це дозволяє зрозуміти, як даний ген впливає на нормальний розвиток. За допомогою складних методів намагаються визначити час і місце дії гена.
Аналіз генетичного контролю ускладнюється рядом причин, насамперед, тим, що роль генів неоднакова. Частина геному складається із генів, які визначають життєво важливі функції або відповідають за синтез речовин, без яких неможливе функціонування жодної клітини (наприклад, гени, що відповідають за синтез т-РНК, або ДНК-поліме-рази). Інші гени безпосередньо беруть участь у детермінації, диференціюванні і морфогенезі, тобто функція їх більш специфічна. Для аналізу генетичного контролю необхідно також знати місце первинної дії гена. Наприклад, при вторинній (відносній) плейотропії існує одне первинне місце дії мутантного гена - гемоглобін в еритроцитах, а всі інші симптоми, які спостерігаються при цій патології (серцева недостатність, збільшення селезінки та ін.) розвиваються як наслідок дії аномального гемоглобіну.
Аналіз мутацій ускладнюється також тим, що розвиток кожного органа і системи органів контролюється сукупною координованою дією сотень генів. У людини відомо понад 120 форм спадкової глухоти, які виникають у результаті експресії мутантних генів, що відповідають за формування слухового аналізатора. Описано близько 250 спадкових уражень зорового аналізатора, 150 спадкових аномалій скелета та ін. Про значення генетичного контролю онтогенезу також свідчать численні хвороби, пов'язані з геномними та хромосомними мутаціями.
Крім експериментальних мутацій, існує цілий ряд інших методів, які дозволяють вивчити роль генів в індивідуальному розвитку. Це методи генної інженерії, клонування, біохімічного аналізу, імунологічні та ін.
Дослідження генетичного контролю онтогенезу має важливе значення для медицини, оскільки це дозволить у майбутньому суттєво знизити рівень спадкової патології. ■ Диференціювання клітин, зародкових листків, тканин
Диференціювання - це процес, внаслідок якого клітина стає більш спеціалізованою, тобто набуває морфологічних і функціональних особливостей. Це зміни, які відбуваються у клітині впродовж одного, нерідко термінального, клітинного циклу. Прикладом може бути диференціювання клітин епідермісу шкіри людини, при якому у клітинах, що переміщуються з базального шару до вищих шарів, відбувається накопичення кератогіаліну, який перетворюєтьс і у клітинах блискучого шару в елеїдин, а потім у роговому шарі - в кератин. При цьому змінюється форма клітин, будова клітинних мембран і набір орга-нел. Насправді диференціюється не одна клітина, а група подібних клітин. Прикладів можна навести багато, оскільки в організмі людини налічується близько 100 видів різних типів клітин. Фібробласти синтезують колаген, міобласти - міозин, клітини епітелію травного тракту - пепсин і трипсин.
У більш широкому розумінні під диференціюванням розуміють поступове наростання відмінностей і спеціалізації між клітинами, які походять від однорідних клітин. Перші хімічні та морфогенетичні відмінності між клітинами відбуваються в період гаструляції.
Зародкові листки та їхні похідні є прикладом раннього диференціювання, яке призводить до обмеження потенцій клітин зародка.
Можна виділити цілий ряд ознак, які характеризують ступінь диференціації клітин. Так, для недиферен-ційованого стану характерні відносно велике ядро і високе ядерно-цитоплазматичне співвідношення, диспергований хроматин і добре виражене ядерце, численні рибосоми та інтенсивний синтез РНК, висока мітотична активність і неспецифічний метаболізм. '
Всі ці ознаки змінюються в процесі диференцію- ^ вання, характеризуючи клітинну спеціалізацію. Про- 1 цес, внаслідок якого окремі тканини в ході дифе- ( ренціювання набувають характерного для них ви- * гляду, називають гістогенезом. 1
Диференціювання клітин під час онтогенезу є ре- І зультатом послідовних реципрокних (взаємних) впливів цитоплазми і змінених продуктів активності ядерних с генів. Таким чином, диференціальна експресія генів є г основним механізмом цитодиференціювання. ІУ ряді випадків остаточне диференціювання пов'язане з "добудовою" молекул ферментів або гормонів. Це вже посттрансляційні події. Наприклад, фермент тирозиназа з'являється у зародків амфібій ще в ранньому ембріогенезі, але переходить в активну форму лише після їх вилуплення.
Іншим прикладом є диференціювання клітин, при якому вони набувають здатності реагувати на хімічні речовини не відразу після синтезу відповідного рецептора, а лише в певний момент. Показано, що м'язові волокна у своїй мембрані мають рецептори до медіаторної речовини ацетилхоліну. Цікаво, що ці хо-лінрецептори виявляли всередині цитоплазми клітин-міобластів до утворення ними м'язових волокон, а чутливість до ацетилхоліну виникала лише з моменту вбудовування рецепторів у плазматичну мембрану під час утворення м'язових трубочок і м'язових волокон. Цей приклад показує, що експресія генів і тканинне диференціювання можуть регулюватися після трансляції у процесі міжклітинних взаємодій.
Таким чином, диференціювання клітин не зводиться лише до синтезу специфічних білків, тому стосовно багатоклітинного організму ця проблема пов'язана з просторово-часовими аспектами і більш високими рівнями її регуляції, ніж біосинтез білка на клітинному рівні.
1.3.3.7
жатальнии період розвитку
людини
• особливості
• критичні періоди
Експериментальне вивчення розвитку тварин показало, що впродовж як онтогенезу взагалі, так і ембріогенезу зокрема існують періоди, коли зародок найбільш чутливий до пошкодження різноманітними факторами, які можуть порушувати нормальний розвиток. Ці періоди отримали назву критичних, тобто -це періоди найменшої стійкості (резистентності) зародків до факторів зовнішнього середовища.
Російський ембріолог П. Г. Свєтлов у 1960 р. сформулював теорію критичних періодів розвитку і перевірив її експериментально. Суть цієї теорії полягає в утвердженні загального положення, що кож-ний етап розвитку зародка в цілому і його окремих органів починається відносно коротким періодом якісно нової перебудови. Ця перебудова супроводжується детермінацією, проліферацією і диференціюванням клітин. У критичні періоди у зародків активується метаболізм, різко посилюється дихання, змінюється вміст РНК, виявляються нові білки й одночасно гальмується темп росту. В цей час ембріон найбільш чутливий і сприйнятливий до пошкоджуючої дії різної природи. Так, у розвитку риб виявлено три критичні періоди: перший - на початку або посередині дроблення; другий - на початку гаструляції і третій - на стадії формування осьових органів. Ікринки на цих стадіях розвитку особливо чутливі до нестачі кисню, високої або низької температур, механічних впливів тощо.
Всебічне вивчення критичних періодів показує, що вони співпадають з активним морфологічним диференціюванням, з переходом від одного періоду розвитку до іншого, зі зміною умов існування зародка. Так, перехід зиготи до дроблення створює для зародка інші умови взаємозв'язку в організмі. Те ж саме відбувається з настанням гаструляції. У ссавців імплантація бластоцисти у стінку матки теж є критичним періодом - переходом до нових умов живлення і газообміну (рис. 1.166). Розвиток плаценти й перехід до плацентарного живлення і газообміну являють собою новий переломний етап у житті зародка і потребують нових пристосувань.
Є підстави вважати, що різні гени починають функціонувати на різних стадіях ембріогенезу, які співпадають з критичними періодами. Такого висновку дійшли на підставі того, що під впливом по-шкоджувальних факторів фізичної й хімічної природи виникають порушення нормального розвитку, які нагадують собою мутації. Російський дослідник Й. А. Рапопорт дією різноманітних хімічних речовин на личинки дрозофіли в різні періоди розвитку домігся модифікаційних змін, які імітують мутації. Так, у дослідах з солями срібла у дрозофіли отримали великий відсоток особин з жовтим тілом, таких самих, як при відповідній мутації.
Що стосується людини, то в її ембріональному розвитку П. Г. Свєтлов розрізняє такі критичні періоди: прогенез (а саме стадія мейозу), запліднення, імплантація, під час якої проходить гаструляція, диференціювання зародкових листків і закладка осьо-вих органів, плацентація, органогенез і період пологів. З останнім критичним періодом в організмі новонародженого пов'язані різкі зміни умов існування і перебудова діяльності всіх систем організму (змінюється характер кровообігу, газообміну, живлення тощо). Крім того, відмічені критичні періоди розвитку окремих органів на різних етапах життя ембріона людини. Вивчення критичних періодів в ембріогенезі показує необхідність захисту материнського організму від шкідливих факторів, особливо на ранніх стадіях вагітності. Умови існування зародка в цей час позначаються на його ембріональному розвитку, а, отже, на всьому наступному періоді життя.