

Режимы мышечных сокращений
При сокращении мышцы в ней могут изменяться два параметра: длина и мышечное напряжение (тонус). В зависимости от того, какое из этих изменений преобладает, различают три режима мышечных сокращений: изотонический, изометрический и ауксотонический. В изотоническом режиме мышцы работают, перемещая какой-либо постоянный груз, как это происходит, например, при сгибании руки в локтевом суставе. В ней может находиться какой-нибудь предмет, хотя это и не обязательно, поскольку и сама рука является «грузом». Так как груз постоянен, постоянно и мышечное напряжение (изотонично), тогда как длина мышцы изменяется (в приведённом примере уменьшается длина сгибателей).
В качестве примера работы мышц в изометрическом режиме можно рассмотреть попытку поднять, например, ведро с водой, Для того, чтобы оторвать его от пола, мышцы должны наращивать напряжение, длина же их остаётся постоянной (изометричной) до тех пор, пока не изменится угловое положение сустава. После того, как груз придёт в движение, мышцы переходят в изотонический режим работы. Если зафиксировано постоянное угловое положение сустава (поэтому длина мышц не изменяется) и происходит мышечное сокращение (для того, например, чтобы поднять или сдвинуть тяжёлый неподвижный предмет или удерживать какой-нибудь груз), то режим работы является изометрическим. При ауксотононическом режиме меняется и длина и тонус мышцы. Если при сокращении длина мышцы уменьшается в связи с увеличением развиваемой силы, такой режим называют ауксотоническим. Он является переходным между изотоническим и изометрическим режимами, поскольку включает две существующие фазы сокращения – динамическую и статическую.
Механизм мышечного сокращения
Специфическим свойством мышц является сократимость. Инициатором мышечного сокращения является ПД, который активирует выделение ацетилхолина в нервно-мышечном синапсе. К мышечному сокращению приводит повышение концентрации ионов кальция в саркоплазме и их присоединение к тропонину. Вследствие этого молекула тропонина изменяет свою конформацию и потому сдвигает тропомиозин в глубину желобка актиновой нити. Так на актиновой нити освобождаются участки, служащие местом прикрепления головок миозина. Внутриклеточный кальций хранится в саркоплазматическом ретикулуме, представляющем систему продольных трубочек и поперечных цистерн. Когда возбуждение передастся через нервно-мышечный синапс на сарколемму, потенциалы действия распространяются по ней к Т-трубочкам, а в связи с этим деполяризуется соприкасающаяся с ними мембрана цистерн саркоплазматического ретикулума. Это приводит к открытию в ней каналов для ионов кальция. Кальций выходит по концентрационному градиенту и присоединяется к тропонину. Весь механизм преобразования процесса возбуждения в мышечное сокращение называется электромеханическим сопряжением. Ключевая роль в нём принадлежит ионам кальция (рис.11).
Рис.11. Электромеханическая сопряжённость
процессов возбуждения в мышечном
волокне:
А – состояние покоя, Б – начало
сокращения, В - расслабление
Сокращение волокна происходит благодаря скольжению актиновых нитей вдоль миозиновых - этому способствует замыкание поперечных мостиков между ними. Длина актиновых и миозиновых нитей при этом не изменяется, но вследствие скольжения уменьшается длина саркомеров, а значит и всего мышечного волокна (рис.12).
Рис.12.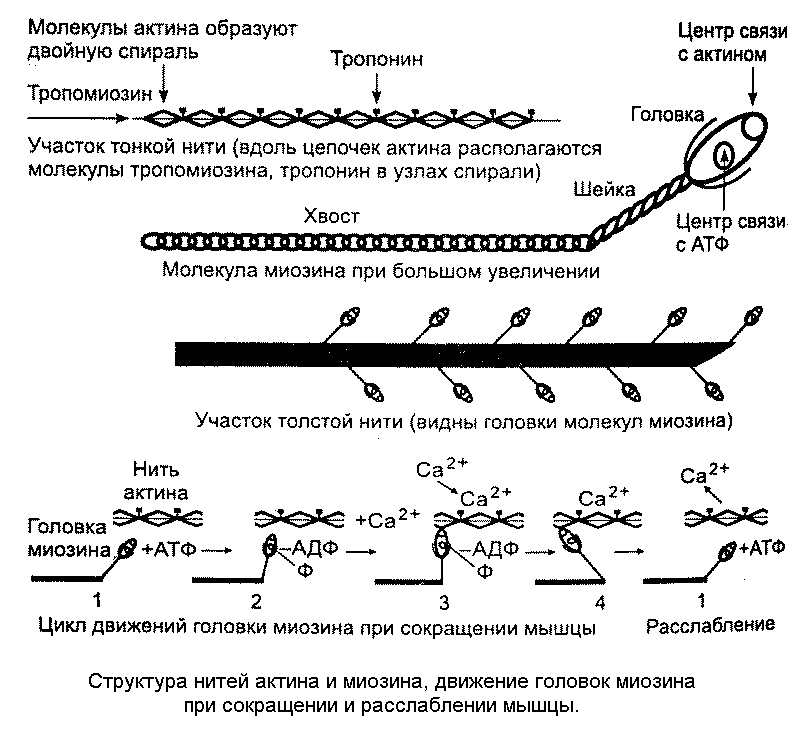
В образовании поперечных мостиков участвуют головки миозина, периодически прикрепляющиеся к соседним нитям актина и создающие усилие, подтягивающее нити актина. Поскольку миозиновые головки одновременно прикрепляются к актиновым нитям, соединённым с двумя соседними Z-мембранами, тянущее усилие сближает Z-мембраны, уменьшая тем самым длину саркомера.
Многочисленные головки миозина прикрепляются к актиновым нитям поочерёдно, а затем, после осуществления тянущего усилия - «гребка», отделяются от этого участка актина, чтобы тотчас прикрепиться к следующему. Эту деятельность можно сравнить с вытягиванием верёвки, которую человек перебирает обеими руками. Для ритмичных прикреплений и отделений миозиновых головок расходуется энергия АТФ, молекулы которой расщепляются под действием каталитически активных центров миозина, который приобретают АТФ-азную активность при взамодействии с актином. Освободившаяся энергия идёт на отделение миозиновой головки, которая тотчас же прикрепляется к соседнему участку актина. Существуют основания полагать, что при мышечном сокращении за одну секунду может происходить от 50 до 100 «гребков».
Высокая концентрация ионов кальция в саркоплазме активирует кальциевый насос, закачивающий эти ионы обратно в саркоплазматический ретикулум. При уменьшении концентрации кальция в саркоплазме тропонин освобождается от него и восстанавливает прежнюю конформацию молекулы. В связи с этим возвращается на прежнее место тропомиозин и уже не позволяет большинству головок миозина прикрепляться к актиновой нити. Когда большинство головок миозина начнёт отделяться от актиновых нитей, наступает расслабление мышечного волокна. Оно происходит пассивно, без затраты энергии: нити актина и миозина скользят друг относительно друга так, что саркомеры увеличивают свою длину (рис.13).
Рис.13.
Таким образом ,последовательность этапов мышечного сокращения состоит из:
Возбуждение нервно-мышечного синапса;
Возбуждение сарколеммы;
Высвобождение ионов кальция из саркоплазматического ретикулума;
Повышение концентрации кальция в саркоплазме;
Связывание кальция с тропонином;
Образование акто-миозинового комплекса;
Укорочение саркомеров за счёт работы поперечных мостиков и скольжении актина по
поверхности миозина.