

Оспа натуральная
Оспа натуральная – острое, часто смертельное, вирусное заболевание. Для него характерны выраженная интоксикация, высокая температура, обильные пузырьковые высыпания на коже и слизистых оболочках, после исчезновения которых остаются втянутые рубцы – оспины.
|
Этиология. Возбудитель оспы относится к вирусам семейства Poxviridae, подсемейства Chordopoxviridae, рода Orthopoxvirus; содержит РНК, имеет крупные размеры 200-350 ммк, размножается в цитоплазме клетки (рис.3). Вирус натуральной оспы имеет антигенное родство с эритроцитами группы А крови человека, что обусловливает слабый иммунитет, высокую заболеваемость и смертность. |
Рис. 3. Вирус оспы (uk.wikipedia.org) |
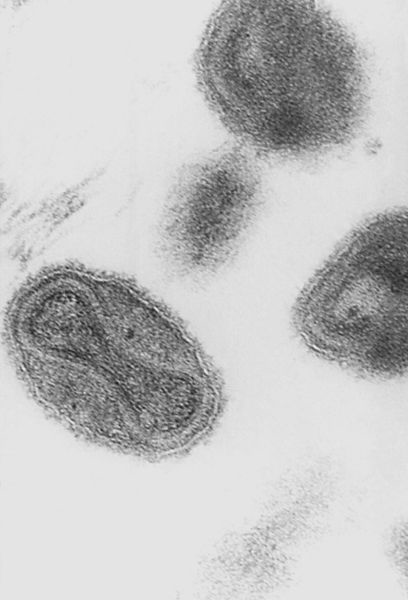
Он устойчив к воздействию внешней среды, особенно к высушиванию и низким температурам.
Эпидемиология. Инфекция строго антропонозная. Возбудитель инфекции передается путем непосредственных контактов, воздушно-капельным способом, от здоровых носителей и животных, и может сохранять жизнеспособность на одежде и постельном белье. Все невакцинированные люди восприимчивы к заражению; естественного иммунитета к натуральной оспе нет. Заболевание возможно в любом возрасте, но дети до четырех лет особенно уязвимы.
Патогенез и клинческая картина. Инкубационный период при натуральной оспе продолжается от 8 до 14 дней. Начальный период болезни характеризуется быстрым подъемом температуры, головной болью, болями в мышцах и пояснице, ознобом и часто рвотой. До появления сыпи на четвертый день болезни установить диагноз натуральной оспы трудно. Обильная сыпь на голове и конечностях сначала имеет вид пятен розового цвета, которые быстро превращаются в везикулы, наполненные прозрачной жидкостью, а затем гноем, напоминая множественные фурункулы. Пузырьки вскрываются и, подсыхая, покрываются корочкой, постепенно исчезая в течение трех недель, в тяжелых случаях оставляя пожизненные следы – оспины. Заболевание варьирует от тяжелейших форм геморрагической, или «черной», оспы с наполненными кровью и гноем пустулами и выраженным общим токсикозом, до легких форм у вакцинированных лиц без сыпи и без повышения температуры.
Диагностика, профилактика, лечение. Натуральная оспа как нозологическая форма ликвидирована в глобальном масштабе. Однако, говорить о полном исчезновении вируса с лица планеты пока рано.
ГРУППА 3. ФАГИ
Бактериофагия – явление лизиса живых бактерий, инфицированных бактериофагом. Вирусы микроорганизмов (актиномицетов, бактерий) получили название фагов (актинофаги и бактериофаги) от греч. «phagos» – пожиратель).
В настоящее время появились сведения о вирофагах – вирусов, пожирателей вирусов. Размеры их колеблются от 40 до 150 нм (рис.4). Фаги неспособны самостоятельно синтезировать нуклеиновые кислоты и белки, поэтому являются облигатными внутриклеточными паразитами. Фаги существуют в двух формах: внутриклеточной (профаг, ДНК) и внеклеточной (вирион).
Фаги, как и другие вирусы, обладают антигенные свойствами и содержат группоспецифические (по ним делятся на серотипы) и типоспецифические антигены. Сыворотки, содержащие антитела к этим антигенам (антифаговые сыворотки) нейтрализуют литическую активность фагов. Взаимодействие бактериофага с клеткой происходит в соответствии с основными типами взаимодействия, характерными для всех вирусов, – продуктивная (литическая), абортивная вирусная и латентная (лизогения, вирогения) инфекция, а также вирус-индуцированная трансформация.
По характеру взаимодействия фага с клеткой бактериофаги делятся на:
вирулентные (литические), вызывающие продуктивную инфекцию и лизис бактериальной клетки,
умеренные, вызывающие латентную инфекцию и ассоциацию генома вируса с бактериальной хромосомой. Умеренные фаги, в отличие от вирулентности, не вызывают гибель бактериальных клеток, и при взаимодействии с ней переходят в неинфекционную форму фага, называемую профагом.
профаг – геном фага, ассоциированный с бактериальной хромосомой. Профаг, ставший частью хромосомы клетки, при ее размножении реплицируется синхронно с геномом бактерии, не вызывая ее лизиса, и передается по наследству от клетки к клетке в неограниченном числе поколений.
Бактериальные клетки, содержащие в своей хромосоме профаг, называются лизогенными. Профаг в лизогенных бактериях самопроизвольно или под влиянием различных индуцированных агентов может переходить в вегетативный фаг. В результате такого превращения бактериальная клетка лизируется и продуцирует новые фаги.
|
В ходе лизогенизации бактериальные клетки могут дополнительно приобретать новые признаки, детерминируемые геномом вируса. Такое явление – изменение свойств микроорганизмов под влиянием профага, называется фаговой, или лизогенной конверсией (проявление вирус-индуцированной трансформации). |
Рис. 4. Бактериофаг (www.byllan.ru) |

Умеренные фаги, неспособные ни при каких условиях переходить из профага в вегетативный фаг (образовывать зрелые фаговые частицы), называются дефектными, чаще это происходит в результате нарушения стадии сборки вирусных частиц. Некоторые умеренные фаги называются трансдуцирующими, поскольку с их помощью осуществляется один из механизмов генетической рекомбинации у бактерий – трансдукции. Такие фаги могут использоваться, в частности в генной инженерии в качестве векторов для получения рекомбинантных ДНК и/или приготовлении рекомбинантных (генно-инженерных) вакцин.
Специфичность фагов послужила основанием для их наименования по видовым и родовым названиям чувствительных к ним бактерий. Так, например, фаги, лизирующие стрептококки, называются стрептококковыми, лизирующие холерный вибрион – холерные, стафилококки – стафилококковыми.
По признаку специфичности выделяют поливалентные бактериофаги, лизирующие культуры одного семейства или рода бактерий, моновалентные (монофаги) – лизирующие культуры только одного вида бактерий, а также отличающиеся наиболее высокой специфичностью – типовые бактериофаги, способные вызывать лизис только определенных типов (вариантов) бактериальной культуры внутри вида бактерий.
Наборы таких типоспецифических фагов используются для дифференцировки бактерий внутри вида – это метод фаготипирования бактерий. С помощью этого метода можно установить источник и пути передачи инфекционного заболевания, т.е. провести его эпидемиологический анализ, поскольку он позволяет сравнивать фаготипы (фаговары) чистых культур бактерий, выделенных в ходе бактериологического исследования от больного и от окружающих его лиц, возможных бактерионосителей.
Мимивирус – паразит амебы – род вирусов, включающий в себя единственный опознанный вид Acanthamoeba polyphaga mimivirus (APMV). Данный вирус отличается наибольшим диаметром капсида из всех известных вирусов, а также, по сравнению с другими вирусами, более объёмным и сложноструктурированным геномом. Возможно, мимивирус представляет собой недостающее звено между вирусами и бактериями или же представляет собой принципиально новую форму жизни, не относящуюся к вирусам или бактериям.
Мимивирус не был до сих пор помещён в какое-либо семейство, но, на основании полученных данных он входит в группу вирусов, содержащих двуцепочечную ДНК и не имеющих РНК-стадии, и отнесен к семейству Mimiviridae. В эту группу входят такие семейства вирусов как иридовирусы, поксвирусы, и другие. Все эти вирусы отличаются крупными размерами, схожими молекулярными характеристиками и сложными геномами.
Геном мимивируса кодирует значительное количество белков, напоминающих эукариотические и бактериальные. По-видимому, эти гены были приобретены мимивирусом вторично и происходят из геномов хозяев вируса и их паразитов. Мимивирус обладает многими свойствами, которые помещают его на границу живого и неживого. По своим размерам он превосходит некоторых бактерий, таких как Rickettsia conorii, содержит геном, сопоставимый по размеру с геномом многих бактерий, и имеет гены, не найденные у других вирусов, в том числе кодирующие ферменты синтеза нуклеотидов и аминокислот, которые отсутствуют даже у некоторых мелких бактерий-внутриклеточных паразитов. Это означает независимость мимивируса от генома клетки-хозяина, кодирующего основные метаболические пути. Однако мимивирус не имеет генов синтеза рибосомальных белков, из-за чего он испытывает необходимость в рибосомах хозяина. Сочетание этих свойств вызвало в научной среде споры, является ли мимивирус особой формой жизни, доменом, наряду с эукариотами, бактериями и археями.
Тем не менее, мимивирус не обладает гомеостазом, не отвечает на раздражители, не растёт и не размножается самостоятельно (вместо этого синтезируется клеткой и самособирается в ней из отдельных компонентов), что типично для вирусов.
Гены, свойственные мимивирусу (в том числе кодирующие белки капсида), сохраняются во множестве вирусов, поражающих организмы всех трёх доменов. На основании этого факта делается предположение, что мимивирус связан с ДНК-содержащими вирусами, которые появились одновременно с наиболее древними организмами, имеющими клеточное строение, и занимают ключевое положение в происхождении жизни на Земле
Вирофаг Спутник (Sputnik virophage) – субвирусный агент, размножающийся в амёбах, заражённых мимивирусом, активно паразитирует на мимивирусе. Имеет икосаэндрическую форму и диаметр около 50 нм. Для размножения вирофаг использует структуры (вирусные фабрики), созданные мимивирусом для собственного воспроизводства. Поэтому вирофаг не может сам по себе размножаться в здоровых амёбах. Фактически вирофаг косвенно паразитирует на мимивирусе. В амёбах, зараженных одновременно мимивирусом и вирофагом, значительно увеличивается доля дефектных вирионов мимивируса. Производство жизнеспособных вирионов мимивируса уменьшается примерно на 70%. Тем самым это примерно втрое уменьшает смертность среди заражённых амёб.