

10 Вопрос. Функциональная классификация таламических ядер.
Таламус (thalamus opticus), или зрительный бугор, – крупное эллипсоидное тело (рис. 3.22). Снизу он сливается с подбугорной областью, от которой отделен гипоталамической бороздой.Латерально таламус граничит с базальными ганглиями больших полушарий, эти структуры разделяет пограничная борозда (рис. 3.30). Медиальная поверхность зрительных бугров образует боковую стенку III желудочка. Эта поверхность отделена от верхней мозговой полоской, которая расширяется в каудальной части, образуя треугольник поводка. Медиальные поверхности двух зрительных бугров соединены межталамическим сращением. На верхней поверхности таламуса в передней его части отчетливо виден передний бугорок, а в основании – утолщение, называемое подушкой (pulvinar). Вентральнее подушки расположены медиальное и латеральное коленчатые тела (рис. 3.27).
Основная пластинка мозговой трубки
эмбриона в среднем мозге заканчивается
и зрительный бугор (вместе со всем
передним мозгом) оказывается
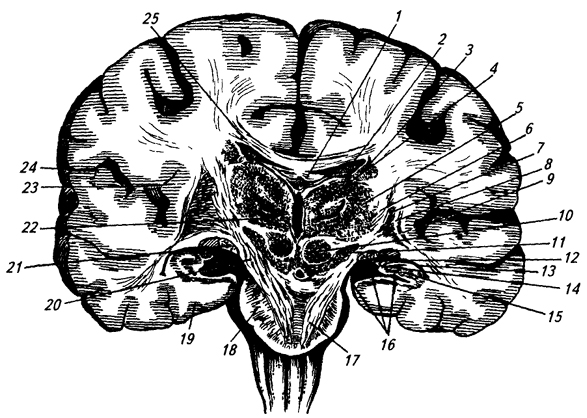 Рис.
3.30. Фронтальный разрез головного мозга
в области ножек мозга:
Рис.
3.30. Фронтальный разрез головного мозга
в области ножек мозга:
1 – свод; 2 – сосудистое сплетение в средней части бокового желудочка; 3 – хвост хвостатого ядра; 4 – медиальное и 5 – латеральное ядра таламуса; 6 – внутренняя капсула; 7 – чечевицеобразное ядро; 8 – наружная капсула; 9 –ограда; 10 – красное ядро; 11 – черная субстанция; 12 – сосудистое сплетение нижнего рога бокового желудочка (13); 14 – бахромка свода; 15 – гиппокамп; 16– гиппокампова борозда; 17 – кортико-спинальный путь; 18 – мост; 19 – парагиппокампова извилина; 20 – основание ножки мозга; 21 – зрительный путь; 22 – вентральное ядро таламуса; 23 – островок; 24 – боковая борозда; 25– мозолистое тело
производным крыльнои пластинки, где нет двигательных нейронов.
Таламус состоит из серого вещества, группирующегося в ядра. Различают пять основных групп ядер таламуса: передние, интраламинарные (внутрипластинчатые), срединные (медиальные), латеральные и задние (см. Атл.). В каждую из этих групп входит по 5–6 и более ядер. Группы ядер разделены V-образной медуллярной (мозговой) пластинкой (laminae medullares), внутри которой также лежат скопления нейронов, называемые интраламинарными ядрами. В переднем ядреоканчивается мамиллоталамический путь, приносящий многокомпонентные импульсы из гипоталамуса. На каудальном полюсе таламуса, в так называемой подушке, оканчивается часть волокон зрительного тракта (см. Атл.).
Наряду с морфологической (по месту локализации) существует функциональная классификация ядер таламуса. Ядра таламуса, нейроны которых направляют аксоны в кору больших полушарий, относят к проекционным или релейным (специфическим). На их нейронах оканчиваются аксоны нейронов восходящих сенсорных систем, кроме обонятельной. Например, в нижней части таламуса лежат основные ядра системы кожной чувствительности и чувствительности опорно-двигательного аппарата – вентролатеральное и вентро-медиальное ядра, в которых оканчиваются спинно-таламический путь, медиальная петля, тройничная петля, волокна верхних ножек мозжечка и другие пути, проводящие импульсы от кожи и проприорецепторов. К ядрам этой группы могут быть отнесены латеральное и медиальное коленчатые тела. В них происходит переключение импульсов соответственно зрительной и слуховой сенсорных систем. Разрушение проекционных ядер приводит к полному выпадению соответствующего вида чувствительности, что указывает на отсутствие иного пути для сенсорной информации в кору.
Проекционные ядра имеют топическую организацию. Это означает, что каждая точка рецептивного (чувствительного) поля данной сенсорной системы проецируется в определенную группу нейронов этих ядер. Чем больше плотность рецепторных окончаний на данном участке поверхности, тем большее число нейронов в ядрах таламуса получает импульсы от этих рецепторов. Поверхность, менее насыщенная рецепторными окончаниями, имеет меньшее представительство. Та же картина повторяется и в коре больших полушарий. Наряду с передачей чувствительности в кору эти ядра выполняют сложную переработку сенсорной информации.
Среди проекционных ядер выделяются и такие, которые получают афферентные импульсы от мозжечка и базальных ганглиев, и передают эфферентные сигналы в моторные области коры. Кроме того, афферентация может приходить к этим ядрам от других подкорковых структур, а проецироваться в лимбические области коры.
Другие ядра, как, например, латеральные и большая часть подушки, передают возбуждение на межанализаторные или ассоциативные области коры; эти ядра относят к ассоциативным.Для них характерно наличие множественных источников афферентации. Информация к этим ядрам поступает от коленчатых тел, других ядер таламуса, миндалевидного комплекса и т.д. Нарушение нейронов ассоциативных ядер и их связей с теменно-затылочными областями коры (таламо-париетальная система) приводит к нарушению распознавания речи, расстройству внимания. Эти ядра образуют также проекции в лобные области коры (таламо-фронтальная
система), благодаря которым осуществляется контроль эмоционального состояния, воспринимается пространство, время и т.д. Ассоциативные ядра таламуса считаются филогенетически молодыми, так как возникают и дифференцируются они в связи с формированием ассоциативных областей новой коры.
Как и в стволовой части мозга, в таламусе есть образования, сходные по функции с ретикулярной формацией. К ним относятся срединные и интраламинарные ядра, а также некоторые ядра латеральной группы. Они оказывают на кору больших полушарий неспецифическое влияние, вызывая не только возбуждающий, но и отчетливый тормозной эффект, и потому получили название неспецифических. Подобно ретикулярной формации ствола неспецифические ядра таламуса не несут каких-либо особых высших интегративных функций, но в известной мере участвуют в регуляции различных афферентных влияний, идущих по восходящим путям сенсорных систем. Эта группа ядер получает афферентацию от большого числа подкорковых структур (красного ядра, черной субстанции, ядер мозжечка, гиппокампа, миндалины и других подкорковых ядер), они не имеют четко организованных проекций в кору. Эти ядра участвуют в организации реакций сон–бодрствование, эмоциональных состояний и т.д.
Отростки нейронов таламуса образуют таламическую лучистость (лучистый венец). Волокна ее направляются к большим полушариям, где они заканчиваются главным образом в коре, а также на клетках базальных ганглиев.
Латеральное коленчатое тело (corpus geniculatum laterale)(рис. 3.27) расположено вблизи подушки таламуса и зрительного тракта, волокна которого в нем оканчиваются. Состоит латеральное коленчатое тело из дорсального и вентрального ядер. Дорсальное ядро имеет слоистое строение, в него поступают афферентные волокна латерального корешка зрительного тракта и ручек переднего двухолмия. Эфферентные волокна идут к 17, 18, 19 полям затылочной доли коры. Вентральное ядро получает афференты от переднего двухолмия, мозжечка и зрительного тракта, эфферентные волокна проходят к интраламинарным ядрам таламуса, переднему двухолмию, покрышке ножек мозга и ретикулярной формации среднего мозга. Прямых выходов на кору это ядро не имеет.
Медиальное коленчатое тело (corpus geniculatum mediale)лежит на уровне поперечной бороздки четверохолмия (рис. 3.27). Оно образовано несколькими ядрами. Афферентные волокна приходят в составе латеральной петли и ручек нижнего двухолмия. Эфферентные волокна в составе слуховой лучистости направляются к коре больших полушарий в 41 и 42 поля.