

Ответ на 1 вопрос.
Центральная нервная система (ЦНС) – это комплекс различных образований спинного и головного мозга, которые обеспечивают восприятие, переработку, хранение и воспроизведение информации, а также формирование адекватных реакций организма на изменения внешней и внутренней среды.
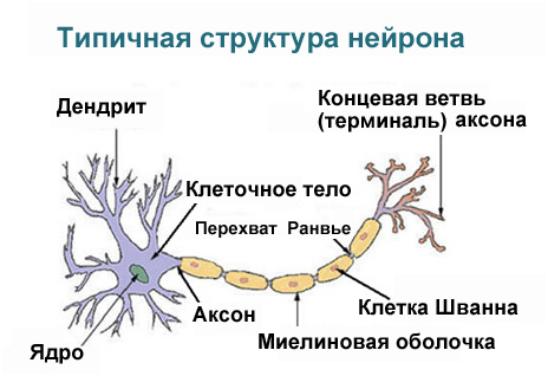
Структурным и функциональным элементом ЦНС являются нейроны. Это высокоспециализированные клетки организма, чрезвычайно различающиеся по своему строению и функциям. В ЦНС нет двух одинаковых нейронов. Мозг человека содержит 25 млрд. нейронов. В общем плане, все нейроны имеют тело – сому и отростки – дендриты и аксоны. Точной классификации нейронов нет. Но их условно разделяют по структуре и функциям на следующие группы:
1. По форме тела:
а. Многоугольные
б. Пирамидные
в. Круглые
г. Овальные
2. По количеству и характеру отростков:
а. Униполярные – имеющие один отросток
б. Псевдоуниполярные – от тела отходит один отросток, который затем делится на 2 ветви.
в. Биполярные – 2 отростка, один дендритоподобный, другой аксон.
г. Мультиполярные – имеют 1 аксон и много дендритов.
3. По медиатору, выделяемому нейроном в синапсе:
а. Холинергические
б. Адренергические
в. Серотонинергические
г. Пептидергические и т.д.
4. По функциям:
а. Афферентные, или чувствительные. Служат для восприятия сигналов из внешней и внутренней среды и передачи их в ЦНС.
б. Вставочные, или интернейроны, промежуточные. Обеспечивают переработку, хранение и передачу информации к эфферентным нейронам. Их в ЦНС большинство.
в. Эфферентные или двигательные. Формируют управляющие сигналы, и передают их к периферическим нейронам и исполнительным органам.
5. По физиологической роли:
а. Возбуждающие
б. Тормозные
Сома нейронов покрыта многослойной мембраной, обеспечивающей проведение ПД к начальному сегменту аксона – аксонному холмику. В соме расположено ядро, аппарат Гольджи, митохондрии, рибосомы. В рибосомах синтезируется тигроид, содержащий РНК, и необходимый для синтеза белков. Особую роль играют микротрубочки и тонкие нити – нейрофиламенты. Они имеются в соме и отростках. Обеспечивают транспорт веществ от сомы по отросткам и обратно. Кроме того, за счет нейрофиламентов происходит движение отростков. На дендритах имеются выступы для синапсов – шипики, через которые в нейрон поступает информация. По аксонам сигнал идет к другим нейронам или исполнительным органам.
Таким образом, общими функциями нейронов ЦНС являются прием, кодирование, хранение информации и выработка нейромедиатора. Нейроны, с помощью многочисленных синапсов, получают сигналы в виде постсинаптических потенциалов. Затем перерабатывают эту информацию и формируют определенную ответную реакцию. Следовательно, они выполняют и интегративную, т.е. объединительную функцию.
Ответ на второй вопрос.
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться.Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном.
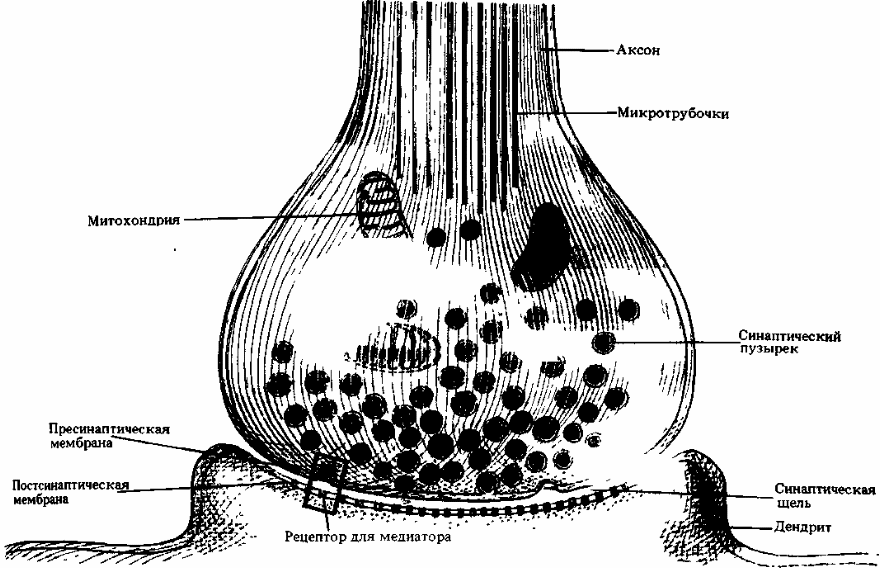
Структура синапса.Типичный синапс — аксо-дендритический химический. Такой синапс состоит из двух частей: пресинаптической, образованной булавовидным расширением окончанием аксона передающей клетки и постсинаптической, представленной контактирующим участком цитолеммы воспринимающей клетки (в данном случае — участком дендрита). Синапс представляет собой пространство, разделяющее мембраны контактирующих клеток, к которым подходят нервные окончания. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём посредством прохождения ионов из одной клетки в другую.
Между обеими частями имеется синаптическая щель — промежуток шириной 10—50 нм между постсинаптической и пресинаптической мембранами, края которой укреплены межклеточными контактами.
Часть аксолеммы булавовидного расширения, прилежащая к синаптической щели, называется пресинаптической мембраной. Участок цитолеммы воспринимающей клетки, ограничивающий синаптическую щель с противоположной стороны, называется постсинаптической мембраной, в химических синапсах она рельефна и содержит многочисленные рецепторы.
В синаптическом расширении имеются мелкие везикулы, так называемые синаптические пузырьки, содержащие либо медиатор (вещество-посредник в передаче возбуждения), либо фермент, разрушающий этот медиатор. На постсинаптической, а часто и на пресинаптической мембранах присутствуют рецепторы к тому или иному медиатору.
Классификации синапсов
В зависимости от механизма передачи нервного импульса различают
химические;
электрические — клетки соединяются высокопроницаемыми контактами с помощью особых коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе — 3,5 нм (обычное межклеточное — 20 нм)
Так как сопротивление внеклеточной жидкости мало(в данном случае), импульсы проходят не задерживаясь через синапс. Электрические синапсы обычно бывают возбуждающими.
Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
смешанные синапсы: Пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Наиболее распространены химические синапсы.
Химические синапсы можно классифицировать по их местоположению и принадлежности соответствующим структурам:
периферические
нервно-мышечные
нейросекреторные (аксо-вазальные)
рецепторно-нейрональные
центральные
аксо-дендритические — с дендритами, в т. ч.
аксо-шипиковые — с дендритными шипиками, выростами на дендритах;
аксо-соматические — с телами нейронов;
аксо-аксональные — между аксонами;
дендро-дендритические — между дендритами;
В зависимости от медиатора синапсы разделяются на
аминергические, содержащие биогенные амины (например, серотонин, дофамин;)
в том числе адренергические, содержащие адреналин или норадреналин;
холинергические, содержащие ацетилхолин;
пуринергические, содержащие пурины;
пептидергические, содержащие пептиды.
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По знаку действия:
возбуждающие
тормозные.
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях.), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор — глицин) и ГАМК-ергические синапсы (медиатор — гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов: 1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала; 2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение. Синапс холинергический (s. cholinergica) — синапс, медиатором в котором является ацетилхолин.
В некоторых синапсах присутствует постсинаптическое уплотнение — электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические — симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы.
Механизм функционирования химического синапса
При деполяризации пресинаптической терминали открываются потенциал-чувствительные кальциевые каналы, ионы кальция входят в пресинаптическую терминаль и запускают механизм слияния синаптических пузырьков с мембраной. В результате медиатор выходит в синаптическую щель и присоединяется к белкам-рецепторам постсинаптической мембраны, которые делятся на метаботропные и ионотропные. Первые связаны с G-белком и запускают каскад реакций внутриклеточной передачи сигнала. Вторые связаны с ионными каналами, которые открываются при связывании с ними нейромедиатора, что приводит к изменению мембранного потенциала. Медиатор действует в течение очень короткого времени, после чего разрушается специфическим ферментом. Например, в холинэргических синапсах фермент, разрушающий медиатор в синаптической щели — ацетилхолинэстераза. Одновременно часть медиатора может перемещаться с помощью белков-переносчиков через постсинаптическую мембрану (прямой захват) и в обратном направлении через пресинаптическую мембрану (обратный захват). В ряде случаев медиатор также поглощается соседними клетками нейроглии.
Открыты два механизма высвобождения: с полным слиянием везикулы с плазмалеммой и так называемый «поцеловал и убежал» (англ. kiss-and-run), когда везикула соединяется с мембраной, и из неё в синаптическую щель выходят небольшие молекулы, а крупные остаются в везикуле. Второй механизм, предположительно, быстрее первого, с помощью него происходит синаптическая передача при высоком содержании ионов кальция в синаптической бляшке.
Следствием такой структуры синапса является одностороннее проведение нервного импульса. Существует так называемая синаптическая задержка — время, нужное для передачи нервного импульса. Её длительность составляет около — 0,5 мс.
Так называемый «принцип Дейла» (один нейрон — один медиатор) признан ошибочным. Или, как иногда считают, он уточнён: из одного окончания клетки может выделяться не один, а несколько медиаторов, причём их набор постоянен для данной клетки.
Ответ на 3 вопрос.
Pефлекс (от лат. "рефлексус" - отражение) - реакция организма на изменения внешней или внутренней среды, осуществляемая при посредстве центральной нервной системы в ответ на раздражение рецепторов.
Рефлексы проявляются в возникновении или прекращении какой-либо деятельности организма: в сокращении или расслаблении мышц, в секреции или прекращении секреции желез, в сужении или расширении сосудов и т. п.
Благодаря рефлекторной деятельности организм способен быстро реагировать на различные изменения внешней среды или своего внутреннего состояния и приспособляться к этим изменениям. У позвоночных животных значение рефлекторной функции центральной нервной системы настолько велико, что даже частичное выпадение ее (при оперативном удалении отдельных участков нервной системы или при заболеваниях ее) часто ведет к глубокой инвалидности и невозможности осуществлять необходимые жизненные функции без постоянного тщательного ухода.
Значение рефлекторной деятельности центральной нервной системы в полной мере было раскрыто классическими трудами И. М. Сеченова и И. П. Павлова. И. М. Сеченов еще в 1862 г. в своем составившем эпоху труде "Рефлексы головного мозга" утверждал: "Все акты сознательной и бессознательной жизни по способу происхождения суть рефлексы".
Виды рефлексов
Все рефлекторные акты целостного организма разделяют на безусловные и условные рефлексы
Безусловные рефлексы передаются по наследству, они присущи каждому биологическому виду; их дуги формиру.ются к моменту рождения и в норме сохраняются в течение всей жизни. Однако они могут изменяться под влиянием болезни.
Условные рефлексы возникают при индивидуальном развитии и накоплении новых навыков. Выработка новых временных связей зависит от изменяющихся условий среды. Условные рефлексы формируются на основе безусловных и с участием высших отделов головного мозга.
Безусловные и условные рефлексы можно классифицировать на различные группы по ряду признаков.
По биологическому значению
пищевые
оборонительные
половые
ориентировочные
позно-тонические (рефлексы положения тела в пространстве)
локомоторные (рефлексы передвижения тела в пространстве)
По расположению рецепторов, раздражение которых вызывает данный рефлекторный акт
экстерорецептивный рефлекс - раздражение рецепторов внешней поверхноcти тела
висцеро- или интерорецептивный рефлекс - возникающий при раздражении рецепторов внутренних органов и сосудов
проприорецептивный (миотатический) рефлекс - раздражение рецепторов скелетных мышц, суставов, сухожилий
По месту расположения нейронов, участвующих в рефлексе
спинальные рефлексы - нейроны расположены в спинном мозге
бульбарные рефлексы - осуществляемые при обязательном участии нейронов продолговатого мозга
мезэнцефальные рефлексы - осуществляемые при участии нейронов среднего мозга
диэнцефальные рефлексы - участвуют нейроны промежуточного мозга
кортикальные рефлексы - осуществляемые при участии нейронов коры больших полушарий головного мозга
NB! (Nota bene - обрати внимание!)
В рефлекторных актах, осуществляемых при участии нейронов, расположенных в высших отделах центральной нервной системы, всегда участвуют и нейроны, находящиеся в низших отделах - в промежуточном, среднем, продолговатом и спинном мозгу. С другой стороны, при рефлексах, которые осуществляются спинным или продолговатым, средним или промежуточным мозгом, нервные импульсы доходят до высших отделов центральной нервной системы. Таким образом, эта классификация рефлекторных актов до некоторой степени условна.
По характеру ответной реакции, в зависимости от того, какие органы в ней участвуют
моторные, или двигательные рефлексы - исполнительным органом служат мышцы;
секреторные рефлексы - заканчиваются секрецией желез;
сосудодвигателъные рефлексы - проявляющиеся в сужении или расширении кровеносных сосудов.
NB! Эта классификация приемлема к более или менее простым рефлексам, направленным на объединение функций внутри организма. При сложных же рефлексах, в которых участвуют нейроны, находящиеся в высших отделах центральной нервной системы, как правило, в осуществление рефлекторной реакции вовлекаются различные исполнительные органы, в результетате чего происходит изменение соотношения организма с внешней средой, изменение поведения организма.Примеры некоторых относительно простых рефлексов, наиболее часто исследуемых в условиях лабораторного эксперимента на животном или в клинике при заболеваниях нервной системы человека.
Как уже отмечалось выше, подобная классификация рефлексов условна: если какой-либо рефлекс может быть получен при сохранности того или иного отдела центральной нервной системы и разрушении вышележащих отделов, то это не означает, что данный рефлекс осуществляется в нормальном организме только при участии этого отдела: в каждом рефлексе участвуют в той или иной мере все отделы центральной нервной системы.
Любой рефлекс в организме осуществляется при помощи рефлекторной дуги.
Рефлекторная дуга - это путь, по которому раздражение (сигнал) от рецептора проходит к исполнительному органу. Структурную основу рефлекторной дуги образуют нейронные цепи, состоящие из рецепторных, вставочных и эффекторных нейронов. Именно эти нейроны и их отростки образуют путь, по которому нервные импульсы от рецептора передаются исполнительному органу при осуществлении любого рефлекса.
В периферической нервной системе различают рефлекторные дуги (нейронные цепи)соматической нервной системы, иннервирующие скелетную иускулатуру
вегетативной нервной системы, иннервирующие внутренние органы: сердце, желудок, кишечник, почки, печень и т.д. Рефлекторная дуга состоит из пяти отделов:
рецепторов, воспринимающих раздражение и отвечающих на него возбуждением. Рецепторами могут быть окончания длинных отростков центростремительных нервов или различной формы микроскопические тельца из эпителиальных клеток, на которых оканчиваются отростки нейронов. Рецепторы расположены в коже, во всех внутренних органах, скопления рецепторов образуют органы чувств (глаз, ухо и т. д.).
чувствительного (центростремительного, афферентного) нервного волокна, передающего возбуждение к центру; нейрон, имеющий данное волокно, также называется чувствительным. Тела чувствительных нейронов находятся за пределами центральной нервной системы - в нервных узлах вдоль спинного мозга и возле головного мозга.
нервного центра, где происходит переключение возбуждения с чувствительных нейронов на двигательные; Центры большинства двигательных рефлексов находятся в спинном мозге. В головном мозге расположены центры сложных рефлексов, таких, как защитный, пищевой, ориентировочный и т. д. В нервном центре происходит синаптическое соединение чувствительного и двигательного нейрона.
двигательного (центробежного, эфферентного) нервного волокна, несущего возбуждение от центральной нервной системы к рабочему органу; Центробежное волокно - длинный отросток двигательного нейрона. Двигательным называется нейрон, отросток которого подходит к рабочему органу и передает ему сигнал из центра.
эффектора - рабочего органа, который осуществляет эффект, реакцию в ответ на раздражение рецептора. Эффекторами могут быть мышцы, сокращающиеся при поступлении к ним возбуждения из центра, клетки железы, которые выделяют сок под влиянием нервного возбуждения, или другие органы.
Простейшую рефлекторную дугу можно схематически представить как образованную всего двумя нейронами: рецепторным и эффекторным, между которыми имеется один синапс. Такую рефлекторную дугу называют двунейронной и моносинаптической. Моносинаптические рефлекторные дуги встречаются весьма редко. Примером их может служить дуга миотатического рефлекса.
В большинстве случаев рефлекторные дуги включают не два, а большее число нейронов: рецепторный, один или несколько вставочных и эффекторный. Такие рефлекторные дуги называют многонейронными и полисинаптическими. Примером полисинаптической рефлекторной дуги является рефлекс отдергивания конечности в ответ на болевое раздражение.
Рефлекторная дуга соматической нервной системы на пути от ЦНС к скелетной мышце нигде не прерывается в отличии от рефлекторной дуги вегетативной нервной системы, которая на пути от ЦНС к иннервируемому органу обязательно прерывается с образованием синапса - вегетативного ганглия.
Вегетативные ганглии, в зависимости от локализации, могут быть разделены на три группы:
позвоночные (вертебральные) ганглии - относятся к симпатической нервной системе. Они расположены по обе стороны позвоночника, образуя два пограничных ствола (их еще называют симпатическими цепочками)
предпозвоночные (превертебральные) ганглии располагаются на большем расстояни от позвоночника, вместе с тем они находятся в некотором отдалении и от иннервируемых ими органов. К числу превертебральных ганглиев относят ресничный узел, верхний и средний шейный симпатические узлы, солнечное сплетение, верхний и нижний брыжеечные узлы.
внутриорганные ганглии расположены во внутренних органах: в мышечных стенках сердца, бронхов, средней и нижней трети пищевода, желудка, кишечника, желчного пузыря, мочевого пузыря, а также в железах внешней и внутренней секреции. На клетках этих ганглий прерываются парасимпатические волокна.
Такое различие соматической и вегетативной рефлекторной дуги обусловлено анатомическим строением нервных волокон, составляющих нейронную цепь, и скоростью проведения по ним нервного импульса.
Для осуществления любого рефлекса необходима целостность всех звеньев рефлекторной дуги. Нарушение хотя бы одного из них ведет к исчезновению рефлекса.
Схема реализации рефлекса
В ответ на раздражение рецептора нервная ткань приходит в состояние возбуждения, которое представляет собой нервный процесс, вызывающий или усиливающий деятельность органа. В основе возбуждения лежит изменение концентрации анионов и катионов по обе стороны мембраны отростков нервной клетки, что приводит к изменению электрического потенциала на мембране клетки.
В двухнейронной рефлекторной дуге (первый нейрон - клетка спинно-мозгового ганглия, второй нейрон - двигательный нейрон [мотонейрон] переднего рога спинного мозга) дендрит клетки спинно-мозгового ганглия имеет значительную длину, он следует на периферию в составе чувствительных волокон нервных стволов. Заканчивается дендрит особым приспособлением для восприятия раздражения - рецептором.
Возбуждение от рецептора по нервному волокну центростремительно (центрипетально) передается в спинно-мозговой ганглий. Аксон нейрона спинномозгового ганглия входит в состав заднего (чувствительного) корешка; это волокно доходит до мотонейрона переднего рога и с помощью синапса, в котором передача сигнала происходит при помощи химического вещества - медиатора, устанавливает контакт с телом мотонейрона или с одним из ее дендритов. Аксон этого мотонейрона входит в состав переднего (двигательного) корешка, по которому центробежно (центрифугально) сигнал поступает к исполнительному органу, где соответствующий двигательный нерв заканчивается двигательной бляшкой в мышце. В результате происходит сокращение мышцы.
Возбуждение проводится по нервным волокнам со скоростью от 0,5 до 100 м/с, изолированно и не переходит с одного волокна на другое, чему препятствуют оболочки, покрывающие нервные волокна.
Процесс торможения противоположен возбуждению: он прекращает деятельность, ослабляет или препятствует ее возникновению. Возбуждение в одних центрах нервной системы сопровождается торможением в других: нервные импульсы, поступающие в центральную нервную систему, могут задерживать те или иные рефлексы.
Оба процесса - возбуждение и торможение - взаимосвязаны, что обеспечивает согласованную деятельность органов и всего организма в целом. Например, во время ходьбы чередуется сокращение мышц сгибателей и разгибателей: при возбуждении центра сгибания импульсы следуют к мышцам-сгибателям, одновременно с этим центр разгибания тормозится и не посылает импульсы к мышцам-разгибателям, вследствие чего последние расслабляются, и наоборот.
Взаимосвязь, определяющая процессы возбуждения и торможения, т.е. саморегуляции функций организма, осуществляется при помощи прямых и обратных связей между центральной нервной системой и исполнительным органом. Обратная связь ("обратная афферентация" по П.К.Анохину), т.е. связь между исполнительным органом и центральной нервной системой, подразумевает передачу сигналов с рабочего органа в центральную нервную систему о результатах его работы в каждый данный момент.
Согласно обратной афферентации, после получения исполнительным органом эфферентного импульса и выполнения рабочего эффекта, исполнительный орган сигнализирует центральной нервной системе о выполнении приказа на периферии.
Так, при взятии рукой предмета глаза непрерывно измеряют расстояние между рукой и целью и свою информацию посылают в виде афферентных сигналов в мозг. В мозгу происходит замыкание на эфферентные нейроны, которые передают двигательные импульсы в мышцы руки, производящие необходимые для взятия ею предмета действия. Мышцы одновременно воздействуют на находящиеся в них рецепторы, беспрерывно посылающие мозгу чувствительные сигналы, информирующие о положении руки в каждый данный момент. Такая двусторонняя сигнализация по цепям рефлексов продолжается до тех пор, пока расстояние между кистью руки и предметом не будет равно нулю, т.е. пока рука не возьмет предмет. Следовательно, все время совершается самопроверка работы органа, возможная благодаря механизму "обратной афферентации", который имеет характер замкнутого круга.
Существование такой замкнутой кольцевой, или круговой, цепи рефлексов центральной нервной системы и обеспечивает все сложнейшие коррекции протекающих в организме процессов при любых изменениях внутренних и внешних условий (В.Д. Моисеев, 1960). Без механизмов обратной связи живые организмы не смогли бы разумно приспособиться к окружающей среде.
Следовательно, вместо прежнего представления о том, что в основе строения и функции нервной системы лежит разомкнутая рефлекторная дуга, теория информации и обратной связи ("обратной афферентации") дает новое представление о замкнутой кольцевой цепи рефлексов, о круговой системе эфферентно-афферентной сигнализации. Не разомкнутая дуга, а сомкнутый круг - таково новейшее представление о строении и функции нервной системы.
Ответ на 4 и 5 вопрос.
Нервный центр — совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт. Представление о структурно-функциональной основе нервного центра обусловлено историей развития учения о локализации функций в центральной нервной системе. На смену старым теориям об узкой локализации, или эквипотенциальности, высших отделов головного мозга, в частности коры большого мозга, пришло современное представление о динамической локализации функций, основанное на признании существования четко локализованных ядерных структур нервных центров и менее определенных рассеянных элементов анализаторных систем мозга. При этом с цефализацией нервной системы растут удельный вес и значимость рассеянных элементов нервного центра, внося существенные различия в анатомических и физиологических границах нервного центра. В результате функциональный нервный центр может быть локализован в разных анатомических структурах. Например, дыхательный центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, в коре большого мозга.
Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований.
1. Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры,
процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
2. Иррадиация возбуждения. Особенности структурной организации центральных нейронов, огромное
число межнейронных соединений в нервных центрах существенно модифицируют (изменяют) направление распространения процесса возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов. Значительное увеличение силы раздражителя приводит к расширению области вовлекаемых в процесс возбуждения центральных нейронов — иррадиации возбуждения.
3. Суммация возбуждения. В работе нервных центров значительное место занимают процессы пространственной и временной суммации возбуждения, основным нервным субстратом которой является постсинаптическая мембрана. Процесс пространственной суммации афферентных потоков возбуждения облегчается наличием на мембране нервной клетки сотен и тысяч синаптических контактов. Процессы временной суммации обусловлены суммацией ВПСП на постсинаптической мембране.
4. Наличие синаптической задержки. Время рефлекторной реакции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка). В нервных клетках высших животных и человека одна синаптическая задержка примерно равна 1 мс. Если учесть, что в реальных рефлекторных дугах
имеются десятки последовательных синаптических контактов, становится понятной длительность большинства рефлекторных реакций — десятки миллисекунд.
Высокая утомляемость. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением. Этот процесс связан с деятельностью синапсов — в последних наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, происходит адаптация постсинаптического рецептора к медиатору.
6. Тонус. Тонус, или наличие определенной фоновой активности нервного центра, определяется тем, что в покое в отсутствие специальных внешних раздражений определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки. Даже во сне в высших отделах мозга остается некоторое количество фоновоактивных нервных клеток, формирующих «сторожевые пункты» и определяющих некоторый тонус соответствующего нервного центра.
7. Пластичность. Функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
8. Конвергенция. Нервные центры высших отделов мозга являются мощными коллекторами, собирающими разнородную афферентную информацию. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10:1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны. На это указывают прямые исследования центральных нейронов: в нервном центре имеется значительное количество поливалентных, полисенсорных нервных клеток, реагирующих на разномодальные стимулы (свет, звук, механические раздражения и т. д.). Конвергенция на клетках нервного центра разных афферентных входов предопределяет важные интегративные, перерабатывающие информацию функции центральных нейронов, т. е. высокий уровень интеграционных функций. Конвергенция нервных сигналов на уровне эфферентного звена рефлекторной дуги определяет физиологический механизм принципа «общего конечного пути» по Ч. Шеррингтону.
9. Интеграция в нервных центрах. Важные интегративные функции клеток нервных центров ассоциируются с интегративными процессами на системном уровне в плане образования функциональных объединений отдельных нервных центров в целях осуществления сложных координированных приспособительных целостных реакций организма (сложные адаптивные поведенческие акты).
10. Свойство доминанты. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в центральной нервной системе. По А.А.Ухтомскому, доминантный нервный очаг характеризуется такими свойствами, как повышенная возбудимость, стойкость и инертность возбуждения, способность к суммированию возбуждения.
В доминантном очаге устанавливается определенный уровень стационарного возбуждения, способствующий суммированию ранее подпороговых возбуждений и переводу на оптимальный для данныхусловий ритм работы, когда этот очаг становится наиболее чувствительным. Доминирующее значение такого очага (нервного центра) определяет его угнетающее влияние на другие соседние очаги возбуждения. Доминантный очаг возбуждения «притягивает» к себе возбуждение других возбужденных зон (нервных центров). Принцип доминанты определяет формирование главенствующего (активирующего) возбужденного нервного центра в тесном соответствии с ведущими мотивами, потребностями организма в конкретный момент времени.
11. Цефализация нервной системы. Основная тенденция в эволюционном развитии нервной системы проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС. Этот процесс называется цефализацией управляющей функции нервной системы. При всей сложности складывающихся отношений между старыми, древними и эволюционно новыми нервными образованиями стволовой части мозга общая схема взаимных влияний может быть представлена следующим образом: восходящие влияния (от нижележащих «старых» нервных структур к вышележащим «новым» образованиям) преимущественно носят возбуждающий стимулирующий характер, нисходящие (от вышележащих «новых» нервных образований к нижележащим «старым» нервным структурам) носят угнетающий тормозной характер. Эта схема согласуется с представлением о росте в процессе эволюции роли и значения тормозных процессов в осуществлении сложных интегративных рефлекторных реакций.
6. Торможение – местный нервный процесс, приводящий к угнетению или устранению возбуждения. В отличие от возбуждения не распространяется по нервным структурам, как ПД
Торможение в ЦНС выполняет две основные функции:
Во-первых, оно координирует функции, т.е. оно направляет возбуждение по определенным путям к определенным нервным центрам, при этом включая те пути и нейроны, активность которых в данный момент не нужна для получения конкретного приспособительного результата. (Пример важности этой функции процесса торможения для функционирования организма можно наблюдать в эксперименте с введением животному стрихнина). Стрихнин блокирует тормозные синапсы в ЦНС (в основном, глицинэргические) и, тем самым, устраняет основу для формирования процесса торможения. В этих условиях раздражение животного вызывает некоординированную реакцию, в основе которой лежит диффузная (генерализованная) иррадиация возбуждения.
Во-вторых, торможение выполняет охранительную или защитную функцию, предохраняя нервные клетки от перевозбуждения и истощения при действии сверхсильных и длительных раздражителей.
В ходе эволюции одновременно с процессом возбуждения формировались ограничивающие и прерывающие его механизмы торможения.
Классификация центрального торможения
По различным признакам:
По электрическому состоянию мембраны –деполяризационное и гиперполяризационное;
По отношению к синапсу –пресинаптическое и постсинаптическое;
По нейрональной организации –поступательное, латеральное (боковое), возвратное и реципрокное.
Постсинаптическое торможение:
Развивается в условиях, когда медиатор, выделяемый нервным окончанием, изменяет свойства постсинаптической мембраны таким образом, что способность нервной клетки генерировать процессы возбуждения подавляется.
Постсинаптическое торможение может быть деполяризационным, если в его основе лежит процесс длительной деполяризации, и гиперполяризационным, если -гиперполяризации.
Поступательное торможение
обусловлено включением тормозных нейронов на пути следования возбуждения
Торможение возвратное (син. антидромное) - процесс регуляции нервными клетками интенсивности поступающих к ним сигналов по способу обратной связи. Он заключается в том, что коллатерали аксона нервной клетки устанавливают синаптические контакты со специальными вставочными нейронами, которые воздействуют на первый нейрон тормозным синапсом (клетка Реншоу по отношению к мотонейрону спинного мозга).
Это механизм, автоматически охраняющий нервные клетки от чрезмерного возбуждения.
Тормозным медиатором у клетки Реншоу является в спинном мозге глицин, ГАМК в коре БП.
Латеральное (боковое) торможение
Вставочные клетки формируют тормозные синапсы на соседних нейронах, блокируя боковые пути распространения возбуждения. В таких случаях возбуждение направляется только по строго определенному пути.
Именно латеральное торможение обеспечивает, в основном, системную (направленную) иррадиацию возбуждения в ЦНС.
Реципрокное торможение
Примером реципрокного торможения является торможение центров мышц-антагонистов.
Суть этого вида торможения заключается в том, что возбуждение проприорецепторов мышц-сгибателей одновременно активирует мотонейроны данных мышц и вставочные тормозные нейроны. Возбуждение вставочных нейронов приводит к постсинаптическому торможению мотонейронов мышц-разгибателей.
7. ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
Иррадиация и концентрация возбуждения и торможения, их взаимная индукция.
-возбуждение и торможение движутся в пространстве ЦНС и во времени
-нервные процессы широко распространяются в нервной системе
-распространение иррадиированной волны возбуждения ограничивается торможением
-процесс взаимного ограничения возбуждения и торможения закон концентрирования возбуждения и торможения.
-иррадиация и концентрация нервных процессов усложняется индукцией –наведением в нервных центрах.
1. РЕЦИПРОКНОСТИ;
2. ОБЩЕГО КОНЕЧНОГО ПУТИ;
3. ДОМИНАНТЫ;
4. СУБОРДИНАЦИИ;
5. ОБРАТНОЙ АФФЕРЕНТАЦИИ.
Процессы возбуждения и торможения в ЦНС не остаются только в тех центрах, где они вызваны, а распространяются (иррадиируют) на другие нервные центры, а также вызывают (индуцируют) друг друга в сопряженно работающих центрах.
Возбуждение и торможение движутся в пространстве ЦНС и во времени. Законы движения возбуждения и торможения определяют координацию т.е. согласованное течение всей сложной рефлекторной деятельности организма.
В норме распространение иррадиированной волны возбуждения ограничивается торможением, которое и определяет течение возбуждения в пространственно определенных участках нервной системы.
Этот процесс взаимного ограничения возбуждения и торможения был назван законом концентрирования возбуждения и торможения.
Иррадиация и концентрация нервных процессов усложняется индукцией → наведением в нервных центрах, одновременно работающих с возбужденным или заторможенным в данный момент, противоположного процесса.
РЕЦИПРОКНАЯ ИННЕРВАЦИЯ
Для центров безусловно-рефлекторной деятельности взаимная индукция наиболее рельефно выступает в центрах сопряженно работающих сгибателей и разгибателей конечностей.
Установлен закон т.н. реципрокной (соотносительной) иннервации мышц-антагонистов → мышцы антагонисты не противодействуют друг другу в работе, а содействуют - в то время, когда происходит сокращение сгибателей, соответствующие им разгибатели расслабляются.
Данный эффект обусловлен тем, что при возбуждении центров сгибателей в центрах разгибателей одноименной стороны индуцируется процесс торможения.
ПРИНЦИП ОБЩЕГО ПОЛЯ
Если иметь в виду только чувствующие нейроны, несущие импульсы к спинному мозгу, то их количество примерно в 5 раз превышает число мотонейронов.
Если же учесть количество вставочных нейронов, которые по существу тоже относятся в воспринимающим раздражение нейронам НС, то количество воспринимающих и анализирующих раздражение внешней среды нервных клеток колоссально возрастает по сравнению с числом нейронов-исполнителей - мотонейронов, сосудодвигательных, секреторных, трофических и т.д..
ЦНС можно представить в качестве "воронки", с широким входным отверстием, куда поступают раздражения с различных рецепторов, и узким выходным отверстием - узким пучком эффекторных нейронов, через которые возбуждение покидает НС.
В эту воронку одновременно вступают импульсы, возникающие при раздражении многих рецепторов. Все они "претендуют" на то, чтобы вызвать возбуждение одной и той же группы мотонейронов, использовать их для осуществления рефлекторного акта.
ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ
Актуальным становится самый сильный раздражитель.
Принцип общего поля обеспечивает использование одних и тех же исполнительных механизмов - мотонейронов с их рабочей периферией - в разнообразных направлениях, для разных целей.
Например, передние конечности животных могут быть использованы и для защитных реакций, и для почесывания, плавания. Человек еще использует верхние конечности для письма, жестикуляции, рисования, игры на музыкальных инструментах и т.д.
ПРИНЦИП ОБРАТНОЙ СВЯЗИ
В осуществлении рефлекторных реакций и их координации огромное значение принадлежит обратной связи, которая осуществляется в результате раздражения проприорецепторов, осморецепторов и др. Импульсы, текущие от них в центры, сигнализируют о степени выполнения действия, могут усилить или затормозить осуществляемый рефлекс.
Положительные обратные связи имеются в тех случаях, когда импульсы с периферии, возникающие в результате какой-либо рефлекторной реакции, ее усиливают.
Отрицательные → когда эти импульсы угнетают рефлекторную реакцию.
Чаще всего отрицательные и положительные обратные связи сосуществуют.
Например, вторичные афферентные импульсы, возникающие при осуществлении сокращения скелетной мускулатуры, вызывают или усиливают возбуждение одних центров, и тормозят другие.
8. МЕТОДЫ ИССЛЕДОВАНИЯ ЦНС
Существуют следующие методы исследования функций ЦНС: 1. Метод перерезок ствола мозга на различных уровнях. Например, между продолговатым и спинным мозгом. 2. Метод экстирпации (удаления) или разрушения участков мозга. 3.Метод раздражения различных отделов и центров мозга. 4. Анатомо-клинический метод. Клинические наблюдения за изменениями функций ЦНС при поражении ее каких-либо отделов с последующим патологоанатомическим исследованием. 5. Электрофизиологические методы: а. Электроэнцефалография - регистрация биопотенциалов мозга с поверхности кожи черепа. Методика разработана и внедрена в клинику Г.Бергером. б. регистрация биопотенциалов нервных различных центров, используется вместе со стереотаксической техникой, при которой электроды с помощью микроманипуляторов вводят в строго определенное ядро в метод вызванных потенциалов, регистрация электрической активности участков мозга при электрическом раздражении периферических рецепторов или других участков; 6. метод внутримозгового введения веществ с помощью микроинофореза. 7. хронорефлексометрия - определение времени рефлексов.
9. СПИННОЙ МОЗГ
Это наиболее простой, примитивный по строению и физиологическим функциям отдел ЦНС.
Спинной мозг представляет собой своеобразный симметричный орган, построенный из однозначных в структурном отношении сегментов, состоящих из серого и белого вещества и связанных с ними двух задних и двух передних корешков.
Задние корешки - состоят из чувствительных проводников, передние - из двигательных (закон Белла-Мажанди).
В спинном мозгу находятся клеточные тела мотонейронов, иннервирующих все скелетные мышцы (за исключением лица) и тела нейронов, направляющих свои волокна к ганглиям вегетативной нервной системы.
ОСНОВНЫЕ РЕФЛЕКСЫ СПИННОГО МОЗГА
Рефлексы растяжения - в основном разгибательные - рефлексы позы, толчковые (прыжок, бег) рефлексы
Сгибательные рывковые рефлексы
Ритмические рефлексы (чесательный, шагательный)
Позиционные рефлексы (шейные тонические рефлексы наклонения и положения)
Вегетативные рефлексы
10. Продолговатый мозг (ПМ) сохраняет в чертах своего строения отдельные признаки сегментарных отношений, типичных для спинного мозга.
Однако правильность в распределении серого и белого вещества здесь значительно нарушена.
В результате структурных и функциональных перестроек, сопровождающих процесс филогенеза, скопления клеточных тел привели к образованию т.н. ядер продолговатого мозга, являющихся центрами рефлекторных функций.
Рефлекторная функция
В продолговатом мозге находятся центры афферентной иннервации различных мимических мышц лица, слизистой оболочки ротовой полости, глаза, лабиринта внутреннего уха и многих внутренних органов - органов дыхания, пищеварения, кровообращения.
Основная биологическая роль рефлекторной деятельности ПМ заключается в обеспечении через регуляцию функций иннервируемых им органов постоянства внутренней среды организма (гомеостаза).
Осуществляется рефлекторная регуляция жизненно важных функций → акта дыхания, защитных рефлексов, связанных с деятельностью дыхательной системы (чихание, кашель), регуляция сердечно сосудистой деятельности, пищеварительного аппарата, рефлексов сосания, жевания, глотания, рвоты, моргания, слезотечения, потоотделения и т.п.
ПМ за счет связи с проприорецепторами выполняет роль регулятора тонуса скелетной мускулатуры.
Обеспечивает тоническое напряжение прежде всего разгибателей, предназначенных для преодоления силы тяжести (позо-тонические рефлексы).
Проводниковая функция
Осуществляется эфферентная нисходящая связь между двигательными зонами коры больших полушарий и двигательными центрами спинного мозга (пирамидный тракт).
Через продолговатый мозг осуществляется и афферентная связь между спинным мозгом и выше лежащими отделами.
ПМ как относительно высший отдел ЦНС регулирует работу более примитивного спинного мозга. Эта координационная функция нацелена на объединение всех сегментов спинного мозга в единое целое, на обеспечение условий для целостной деятельности спинного мозга.
Повреждение продолговатого мозга связано с нарушением или полным выпадением функций, регулируемых этим отделом ЦНС.
Полное разрушение продолговатого мозга ведет к гибели животных, для которых относительное постоянство внутренней среды является обязательным условием существования.
У человека заболевания продолговатого мозга чрезвычайно тяжело сказываются на всех видах деятельности организма.
Непосредственной причиной неизбежной смерти является нарушение деятельности дыхательной системы.
11.Физиология среднего мозга.
В строении среднего мозга (СМ) окончательно утрачиваются сегментарные признаки.
Клеточные элементы образуют здесь сложные скопления в виде ядерных образований, относящихся как непосредственно к среднему мозгу, так и входящих в состав ретикулярной формации мозгового ствола.
Ядра сенсорного, афферентного значения располагаются в дорзальной части СМ (четверохолмие), ядра эфферентного значения - в вентральной его части (красное ядро, черная субстанция и др.).
Через средний мозг в составе т.н. ножек мозга и мозжечка проходят многочисленные проводящие пути, связывающие в восходящем и нисходящем направлениях между собой большой мозг и мозжечок, продолговатый и спинной мозг.
обеспечивает расширение и усовершенствование рефлекторной деятельности продолговатого мозга;
•координация функций организма сигналами со зрительных и слуховых рецепторов;
•окончательно утрачиваются сегментарные признаки;
•клеточные элементы сложные скопления в виде ядерных образований (средний мозг и ретикулярная формация мозгового ствола).
Основные ядра среднего мозга
Ядра черепномозговых нервов:
III пара -глазодвигательного нерва
IV пара -блокового нерва ---
Ядро Даркшевича - продольный пучок среднего мозга, связывающий ядра глазодвигательного, блокового и отводящего нерва в единую систему
Непарное вегетативное ядро Якубовича-Эдингера - через цилиарный ганглий к мышцам радужки и ресничного тела
Ядра тектальной области:
верхнее или переднее двухолмие -зрительные рефлексы;
нижнее или заднее двухолмие - слуховые рефлексы – четверохолмие
Черная субстанция
Красные ядра
12. Физиология мозжечка.
Мозжечок входит в состав заднего мозга.
У человека этот орган достигает своего наивысшего развития и занимает большую часть задней черепной ямки.
Мозжечок связан с другими отделами мозга с помощью афферентных и эфферентных путей.
Деятельность мозжечка имеет ближайшее отношение к осуществлению произвольных движений.
Однако повреждение мозжечка не влечет за собой двигательных и сенсорных параличей!
ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
РЕГУЛЯЦИЯ ПОЗЫ И МЫШЕЧНОГО ТОНУСА
КОРРЕКЦИЯ МЕДЛЕННЫХ ЦЕЛЕНАПРАВЛЕННЫХ ДВИЖЕНИЙ И ИХ КООРДИНАЦИЯ С РЕФЛЕКСАМИ ПОДДЕРЖАНИЯ ПОЗЫ
ПРАВИЛЬНОЕ ВЫПОЛНЕНИЕ БЫСТРЫХ ЦЕЛЕНАПРАВЛЕННЫХ ДВИЖЕНИЙ ПО КОМАНДАМ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ В СТРУКТУРЕ ОБЩЕЙ ПРОГРАММЫ ДВИЖЕНИЙ
Мозжечок нельзя считать органом равновесия, ему нельзя приписывать и значение органа, координирующего движения тела.
Роль мозжечка сводится к воздействиям на нижележащие центры, направленные на поддержание тонуса, устойчивости реакций в нервных центрах в точном соответствии этих реакций с текущими задачами, выдвигаемыми перед организмом в данный момент.
Функция мозжечка заключается в рефлекторном обеспечении правильного перемещения тела в пространстве, что в свою очередь достигается обеспечением правильного напряжения различных мышечных групп, торможением лишних движений, лишних примитивных двигательных рефлексов.
Физиологической основой этой функции являются рефлекторные регулирующие влияния на двигательные нейроны, иннервирующие скелетную мускулатуру.
13. Зрительный бугор и его ядра.
Главными образованиями промежуточного мозга являются зрительные бугры и подбугровая область.
Зрительный бугор (thalamus opticus) является своеобразной сенсорной промежуточной станцией - областью переключения всех афферентных путей, идущих к коре больших полушарий. Бугор является средоточием всех рецептивных нейронов ЦНС, выполняя роль высшего подкоркового центра всей чувствительности тела. Нервные связи бугра с соседними областями головного мозга отличаются исключительным обилием и функциональным многообразием.
Основные ядра таламуса
Специфические ядра
переключающие
ассоциативные
моторные
Неспецифические ядра
срединные ядра, надколенное ядро, пограничное ядро, парафасцикулярное ядро, ретикулярное ядро (проекция к полосатому телу и V - VI слоям всех областей коры больших полушарий
Волокна от специфических ядер образуют синапсы на ограниченном числе зон коры, а волокна от неспецифических ядер таламуса дают большое количество разветвлений в разных участках коры больших полушарий и вовлекают в процесс возбуждения большое количество корковых нейронов.
Показано, что неспецифическая система таламуса принимает участие в быстрой и кратковременной активации коры в противоположность медленной и длительной активации, осуществляемой ретикулярной формацией мозга.
РФ среднего мозга выполняет функции поддержания тонуса всей коры (см. ниже), а неспецифические ядра таламуса активируют лишь те ее структуры, которые принимают участие в осуществлении конкретных рефлекторных реакций (организация процесса внимания).
Таламус имеет большое значение как центр формирования ощущений, в частности - как высший центр формирования болевой чувствительности.
Еще является центром непроизвольных выразительных движений и эмоциональных проявлений.
Разрушение таламуса приводит к выпадению чувствительности и выпадению сокращений мускулатуры лица, непроизвольно сокращающейся при эмоциях - маска страха, гнева, плача
Основные функции ассоциативных систем таламуса
ТАЛАМОПАРИЕТАЛЬНАЯ СИСТЕМА
1) Центральный аппарат анализа и синтеза обстановочной афферентации, запуска ориентационных движений глаз и туловища
2) Один из центральных аппаратов «схемы тела» и сенсорного контроля текущей двигательной активности
3) Аппарат формирования полимодальных образов
ТАЛАМОФРОНТАЛЬНАЯ СИСТЕМА
Корковый модулятор лимбической системы, программирование целенаправленных поведенческих актов на основе опыта и мотивации
14. Гипоталамус. Ядра гипоталамуса.
В состав подбугровой области входят следующие основные ядра: серый бугор, nucleus paraventricularis, nucleus supraopthicus, corpora mamillaria.
Ядра гипоталамической области связаны с ядрами вегетативных нервов среднего, продолговатого и спинного мозга.
ФУНКЦИИ ГИПОТАЛАМУСА:
Высший центр регуляции вегетативной нервной системы
Высший центр регуляции эндокринных функций
Регуляция мотиваций пищевого поведения
Высший трофический центр
Вегетативное обеспечение и реализация эмоций
Половые, оборонительные, агрессивные мотивации
Участие в терморегуляции
Участие в регуляции цикла «сон - бодрствование»
15. Роль гипоталамуса в осуществлении вегетативных функций обмена веществ.
Гипоталамус является главным подкорковым центром вегетативной нервной системы, промежуточным звеном, связывающим основные воспринимающие образования НС с вегетативными ганглиями на периферии.
Является центральным источником импульсов для осуществления вегетативных реакций.
При раздражении серого бугра возникают зрачковые, сосудистые реакции, изменение потоотделения и обмена веществ.
16. Лимбическая система
филогенетически старые отделы переднего мозга и производные подкорковых структур;
зоны, отделяющие неокортекс от гипоталамуса (поясная и гиппокампова извилина, обонятельный мозг(rhynencеphalon),миндалина, септальные ядра и переднее таламическое ядро;
(преоптическое ядро, гипоталамус и мамиллярные тела)
ОСНОВНЫЕ ФУНКЦИИ ЛИМБИЧЕСКОЙ СИСТЕМЫ
1. Организация вегетативно-соматических компонентов эмоций
2. Организация кратковременной и долговременной памяти
3. Участие в формировании ориентировочно-исследовательской деятельности
4. Организация простейшей мотивационно-информационной коммуникации (речи)
5. Участие в механизмах сна
6. Центр обонятельной сенсорной системы
Функциональная структура лимбики по МакЛину
Нижний отдел - миндалина и гиппокамп - центры эмоций и поведения для выживания и самосохранения
Верхний отдел - поясная извилина и височная кора - центры общительности и сексуальности
Средний отдел - гипоталамус и поясная извилина - центры биосоциальных инстинктов
17. Базальные ядра (подкорковые ядра), базальные ганглии, скопление серого вещества больших полушарий головного мозга позвоночных, участвующие в координации двигательной активности и формировании эмоциональных реакций. Базальные ядра вместе с корой мозга составляют клеточное вещество конечного мозга. Состоят из хвостатого ядра, скорлупы (объединяются в полосатое тело), бледного шара (объединяются со скорлупой в чечевицеобразное ядро), ограды, миндалевидного тела. У рыб и земноводных базальные ядра представлены только бледным шаром. У пресмыкающихся впервые появляются хвостатое ядро и скорлупа, особенно хорошо развитые у птиц. Базальные ядра характеризуются множественными афферентными и эфферентными связями с корой больших полушарий, средним и промежуточным мозгом, лимбической системой и мозжечком. Для нормального функционирования базальных ядер исключительно важное значение имеет дофамин, выполняющий роль тормозного медиатора, а также ацетилхолин.
Кроме коры, образующей поверхностные слои конечного мозга, в глубине каждого полушария имеются скопления серого вещества в виде отдельных ядер. Эти ядра находятся в толще белого вещества , ближе к основанию мозга, поэтому получили название базальных, или подкорковых ядер. Это старое топологическое название не отражает ни происхождения, ни функций этих структур.
К базальным ядрам по традиции относят полосатое тело и миндалевидное тело. В состав полосатого тела в свою очередь входят хвостатое ядро, чечевицеобразное ядро и ограда.
Они получают информацию об активности двигательной системы (полосатое тело, ограда и миндалевидное тело), управляют движениями и регулируют мышечный тонус. В толще белого вещества под мозолистым телом располагаются боковые желудочки, которые являются полостями полушарий большого мозга. Таким образом, в каждом полушарии условно можно выделить 4 подкорковых ядра:
- хвостатое ядро
- чечевицеобразное ядро
- ограду
- миндалевидное тело
Бледный шар представляет собой филогенетически более древнее образование древнее полосатое тело и отличается от других частей полосатого тела не только по макроскопическому виду, но и гистологически. Новое полосатое тело формируется млекопитающих в связи с развитием сенсомоторных центров новой коры. В неостриатум входят хвостатое ядро, скорлупа и ограда.
Ограда расположена кнаружи от чечевицеобразного ядра. Она представляет собой пластинку серого вещества толщиной до 2 мм. Медиальный край пластинки ровный, а на латеральном крае имеются небольшие выпячивания серого вещества. Миндалевидное тело находится в толще белого вещества височной доли полушария и принадлежит старому полосатому телу. Миндалевидное тело относится к подкорковым обонятельным центрам и к лимбической системе. Базальные ядра полушарий отделены друг от друга прослойками белого вещества, которые в данном случае называются капсулами . Различают три капсулы: внутреннюю капсулу (capsula interna), наружную капсулу и крайнюю капсулу.
Внутренняя капсула - это толстая, изогнутая под углом пластинка белого вещества, расположенная между головкой хвостатого ядра, таламусом и бледным шаром чечевицеобразного ядра (рис. 63, рис. 64). Между хвостатым ядром и чечевицеобразным ядром находится передняя ножка внутренней капсулы, между таламусом и чечевицеобразным ядром - задняя ножка внутренней капсулы. Обе ножки соединяются под углом, открытым латерально. Место соединения ножек называется коленом внутренней капсулы. У низших позвоночных со слабо развитой корой больших полушарий базальные ядра выполняют функцию основного интегративного аппарата головного мозга, у высших - сохраняют важную роль в регуляции произвольных движений.
18. Кора больших полушарий головного мозга или кора головного мозга (лат. cortex cerebri) — структура головного мозга, слой серого вещества толщиной 1,3—4,5 мм, расположенный по периферии полушарий большого мозга, и покрывающий их. Наибольшая толщина отмечается в верхних участках предцентральной, постцентральной извилин и парацентральной дольки.
Кора головного мозга играет очень важную роль в осуществлении высшей нервной (психической) деятельности.
У человека кора составляет в среднем 44% от объёма всего полушария в целом. Площадь поверхности коры одного полушария у взрослого человека в среднем равна 220 000 мм². На поверхностные части приходится 1/3, на залегающие в глубине между извилинами — 2/3 всей площади коры.
Величина и форма борозд подвержены значительным индивидуальным колебаниям — не только мозг различных людей, но даже полушария одной и той же особи по рисунку борозд не вполне похожи.
Всю кору полушарий принято разделять на 4 типа: древняя (палеокортекс), старая (архикортекс), новая (неокортекс) и межуточная кора. Поверхность неокортекса у человека занимает 95,6%, старой 2,2%, древней 0,6%, межуточной 1,6%[
Слои коры больших полушарий
1 слой - верхний молекулярный - ветвления дендритов
пирамидных нейронов, редкие горизонтальные нейроны
и клетки-зерна, волокна неспецифических ядер таламуса.
2 слой - наружный зернистый - звездчатые клетки, пути
реализующие циркуляцию импульсов, волокна
неспецифических ядер таламуса.
3 слой - наружный пирамидный - малые пирамидные клетки и корково-корковые связи различных извилин коры.
4 слой - внутренний зернистый - звездчатые клетки, окончание специфических таламокортикальных путей.
5 слой - внутренний пирамидный - крупные пирамидные
клетки Беца - выходные нейроны кортико -мозговых путей.
6 слой - полиморфных клеток - кортикоталамические пути.
ФУНКЦИОНАЛЬНЫЕ ЗОНЫ КОРЫ
СЕНСОРНЫЕ (зрительные, слуховые, кожные и др.);
МОТОРНЫЕ (первичные, вторичные, комплексные);
АССОЦИАТИВНЫЕ (лобные, теменные, височные)
полисенсорность, пластичность, длительность хранения следов.

ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ
1. Управление врожденными поведенческими реакциями при помощи накопленного опыта;
2. Согласование внешних и внутренних мотиваций поведения;
3. Разработка стратегии поведения и программы действия;
4. Мыслительные особенности личности.
19. Инсти́нкт — совокупность врождённых тенденций и стремлений, играющих мотивационную роль в формировании поведения. В узком смысле, совокупность сложных наследственно обусловленных актов поведения, характерных для особей данного вида при определённых условиях. Инстинкты составляют основу поведения животных. У высших животных инстинкты подвергаются модификации под влиянием индивидуального опыта.
Общепринятого определения инстинкта не разработано до сих пор. Некоторые проблемы, в частности, применимость термина инстинкт по отношению к человеку носят дискуссионный характер.
Инстинкт — комплекс двигательных актов или последовательность действий, свойственных организму данного вида, реализация которых зависит от функционального состояния животного (определяемого доминирующей потребностью) и сложившейся в данный момент ситуации. Инстинктивные реакции носят врожденный характер, и их высокая видовая специфичность часто используется как таксономический признак наряду с морфологическими особенностями данного вида животных.
Отличия инстинкта и рефлекса
Инстинкт - это комплекс действий, а рефлекс - одиночное действие. Инстинкт - это врожденный набор действий, который запускается в определенных условиях и производится неосознанно.
Рефлекс, в отличие в инстинкта - это простое и одиночное действие.
Для животных защищать потомство - это инстинкт, а сесть по команде - рефлекс. В первом случае это целая цепочка действий: найти укромное логово, проложить к нему невидимые тропы, не охотиться возле логова и так далее. А во втором случае - действие одно: посадить попу на землю.
Теорий инстинктов много. Многие этологи долгое время не могли придумать определение инстинкту.
К.Лоренц предположил, что инстинкт - это комплекс врожденных действий, которые запускаются в определенных условиях и при наличии драйва.
Долгое время после этого формулировали - что же такое драйв. В итоге малопонятный "драйв" заменили понятием о "функциональном состоянии организма", то есть о таком особенном состоянии всех систем организма, при котором возможна реализация инстинктивного поведения.
Например у маленького волчонка есть уже заложенное генетически половое поведение. Но волчицами он интересуется пока не очень - стимул есть, действия есть, неужных гормонов, развитиых центров головного мозга, развитых половых желез и др. - нет.
Есть гипотеза, что инстинкт в мире людей называется шаблоном поведения.
20. ФИЗИОЛОГИЯ РЕТИКУЛЯРНОЙ ФОРМАЦИИ
Структуры ретикулярной формации были описаны еще в прошлом веке немецким ученым Дейтерсом. Эти структуры находятся во многих отделах головного мозга. В. И. Бехтерев подобные образования обнаружил в спинном мозге. Таким образом, установлено, что ретикулярная формация имеется практически во всех отделах ЦНС, исключая кору головного мозга. Основная функциональная роль ретикулярной формации приписывается стволу мозга: продолговатому мозгу, варолиеву мосту, среднему мозгу. Большинство физиологов считает, что главным открытием ХХ-го века в области физиологии является выяснение функции особой структуры мозга, получившей название ретикулярной формации. В разработку этой проблемы физиологии большой вклад внесли известные отечественные и зарубежные ученые такие как Джаспер (Канада), Моруцци (Италия), Мегун (США), Анохин (СССР) и др.
Эти структуры, с анатомической или гистологической точки зрения, представляют собой нейроны разной величины и формы, имеющие многочисленные отростки, которых в сотни раз больше, чем в других отделах ЦНС. Поэтому ретикулярная формация получила и дополнительное название - сетчатая структура.
Как было отмечено раньше, особая физиологическая роль принадлежит структурам ретикулярной формации, расположенным в области ствола мозга. В чем она заключается? Как удалось выяснить, ретикулярная формация ствола мозга оказывает влияние на нижерасположенные отделы ЦНС, т. е. на сегментарный аппарат спинного мозга. Такое влияние получило название нисходящего и выражается в действии ретикулярной формации на мотонейроны спинного мозга. В ретикулярной формации ствола мозга выделено два вида структур: одни получили название облегчающих, другие - тормозных, а в целом их название подразумевает их влияние на функциональное состояние мотонейронов спинного мозга (следовательно, и на сократительную способность мышечной системы).
В классических исследованиях Мегуна и Моруцци кошке в состоянии наркоза в облегчающие структуры ствола мозга вживлялись электроды и записывался коленный рефлекс при ударе молоточка по сухожилию четырехглавой мышцы. При электрической стимуляции облегчающих структур имело место увеличение коленного рефлекса и повышение мышечного тонуса. Если же электроды вживлялись в тормозные структуры, то при их раздражении величина коленного рефлекса заметно уменьшалась, так же падал и мышечный тонус. Ретикулярная формация изменяет активность мотонейронов спинного мозга опосредовано через вставочные нейроны.
Кроме нисходящих влияний в стволе мозга были обнаружены структуры, оказывающие восходящее влияние на вышележащие отделы ЦНС, а именно, на корковые нейроны. Эти влияния носят медленный и длительный характер, тормозя или активируя нервные клетки. Более изученным является активирующее влияние. Возможно потому, что оно имеет более важное значение. Следует помнить, что это влияние носит неспецифический (диффузный) характер, т. е. оно практически касается увеличения активности всех корковых нейронов без исключения. Для доказательства такого неспецифического влияния ретикулярной формации на корковые нейроны животным вживляются многочисленные электроды, с которых снимались электрические реакции в виде фоновой электрокортикограммы. Затем через электроды, вживленные в активирующие структуры ствола мозга, проводилась электростимуляция. В этом опыте на всех без исключения отведениях электрокортикограммы наблюдалось уменьшение амплитуды и увеличение частоты волн, свидетельствующие о возрастании активности корковых нейронов (реакция десинхронизации). Еще одним свидетельством в пользу активирующего и тормозного влияния ретикулярной формации на активность корковых нейронов является тот факт, что если во время сна животному раздражать активирующие структуры, то оно просыпается (реакция пробуждения), а если их разрушить, то животное погружается в сон.
Активирующее влияние ретикулярной формации на корковые нейроны создает рабочий тонус коре головного мозга, иначе говоря, “держит” нервные клетки в состоянии постоянного возбуждения. Только на фоне такого постоянного возбуждения корковых нейронов нормально проявляются все функции коры головного мозга (замыкательная деятельность, усвоение, переработка поступающей информации и т. д). Таким образом, ретикулярная формация является своеобразной структурой, как бы аккумулятором, постоянно подпитывающим корковые нейроны, что способствует их функциональной организации.
Показано, в частности, что на фоне стимуляции определенных структур ретикулярной формации быстрее вырабатываются условные рефлексы и лучше усваиваются другие формы информации. В то же самое время разрушение определенных структур ретикулярной формации сопровождается затруднением выработки условных рефлексов и ухудшением фиксации других видов информации.
В ретикулярной формации ствола мозга имеются образования, оказывающие восходящее тормозное влияние на деятельность корковых нейронов. Однако, это тормозное влияние ее не проявляется так сильно, как активирующее, и, возможно, имеет меньшее физиологическое значение. В свою очередь, получены экспериментальные данные, свидетельствующие о том, что кора головного мозга, в свою очередь, в определенных случаях может влиять на активность ретикулярной формации. Ретикулярная формация постоянно получает информацию с многочисленных анализаторов (чувствительного, зрительного, слухового и др.), благодаря которым создается ее собственный тонус и постоянное возбуждение ЦНС.
Кроме того, тонус ретикулярной формации имеет и гуморальную природу. Такими гуморальными веществами для ретикулярной формации являются многочисленные медиаторы, гормоны и другие биологически активные вещества, содержащиеся в крови и взаимодействующие с ее специфическими рецепторами (адрено-, холино- и др.). Доказано, что в ретикулярной формации мозга преобладают адренорецепторы, при взаимодействии с которыми биологически активные вещества возбуждают ретикулярную формацию и обусловливают ее тонус. Наряду с этим, в ретикулярной формации имеются и холинэргические образования, при действии на которые указанные выше некоторые вещества приводят к подавлению активности ретикулярной формации и уменьшению ее тонуса. Например, применение в клинике аминазина, блокирующего адренорецепторы ЦНС, сопровождается снижением активности ретикулярной формации. В результате наблюдается угнетение активности корковых нейронов, обусловливающее успокоительное действие аминазина на нервную систему и вызывающее даже сон. В связи с таким эффектом – опосредованным эффектом влияния на ретикулярную формацию - аминазин нашел широкое применение в медицине, особенно, в психоневрологической практике.
21. функции вегетативной нервной системы
Основная функция ВНС–регуляция деятельности внутренних органов.
Симпатическая НС, как правило, вызывает
мобилизацию деятельности жизненно важных органов,
повышает энергообразование в организме –за счет активации процессов гликогенолиза, глюконеогенеза, липолиза,
оказывает эрготропное влияние.
Парасимпатическая НС оказывает трофотропное действие, способствует восстановлению нарушенного во время активности организма гомеостаза.
Метасимпатическая НС оказывает регулирующее воздействие на мышечные структуры в ЖКТ, регулируя его моторику, и в сердце, регулируя его сократительную активность.
Известен тройственный характер влияния НС на ткани и органы:
пусковое или корригирующее - приводит в деятельное состояние тот или иной орган или изменяет его функцию;
сосудодвигательное - заключается в том, что путем изменения просвета сосудов увеличивается или уменьшается кровоснабжение данного органа;
трофическое - за счет которого изменяется течение тканевого обмена веществ. В этом случае создаются благоприятные условия для работы органа.
Одним из отличительных свойств ВНС является более активное выделение ею медиаторов и больший удельный вес их в передаче возбуждения на ткани.
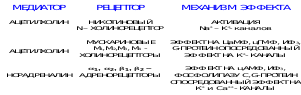
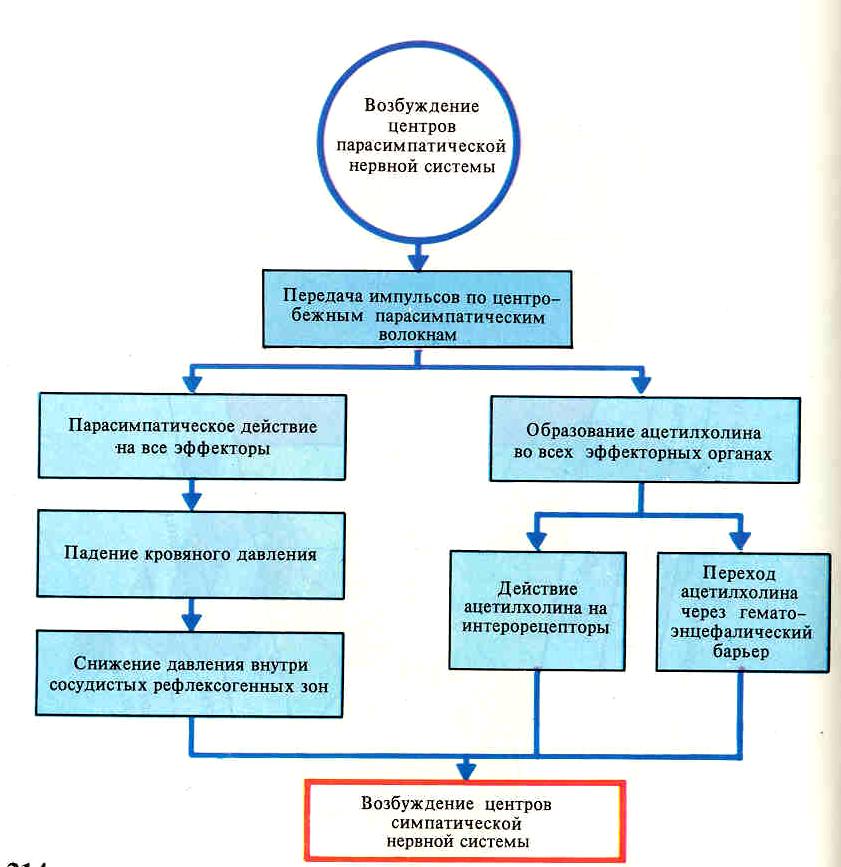
22. Взаимоотношения между симпатической и парасимпатической НС
В ряде случаев влияния, оказываемые симпатической и парасимпатической НС носят противоположный характер.
Известно, что симпатические нервы стимулируют деятельность сердца, а блуждающий тормозит, симпатикус тормозит деятельность гладких мышц кишечника, а парасимпатикус - стимулирует.
Следует помнить, что такие "антагонистические" отношения проявляются не всегда и не везде. В ряде органов функциональный антагонизм отсутствует.
Нельзя говорить об антагонистических отношениях симпатических нервов, расширяющих зрачок, и парасимпатических, суживающих его. В этом случае оба типа волокон оказывают стимулирующее влияние, но на разные мышцы. Даже тогда, когда орган имеет и симпатическую, и парасимпатическую иннервацию, антагонизм часто отсутствует.
Так, для слюнных желез секреторным для жидкой фазы является парасимпатикус, а для ферментов – симпатикус.
Симпатические и парасимпатические эффекты

2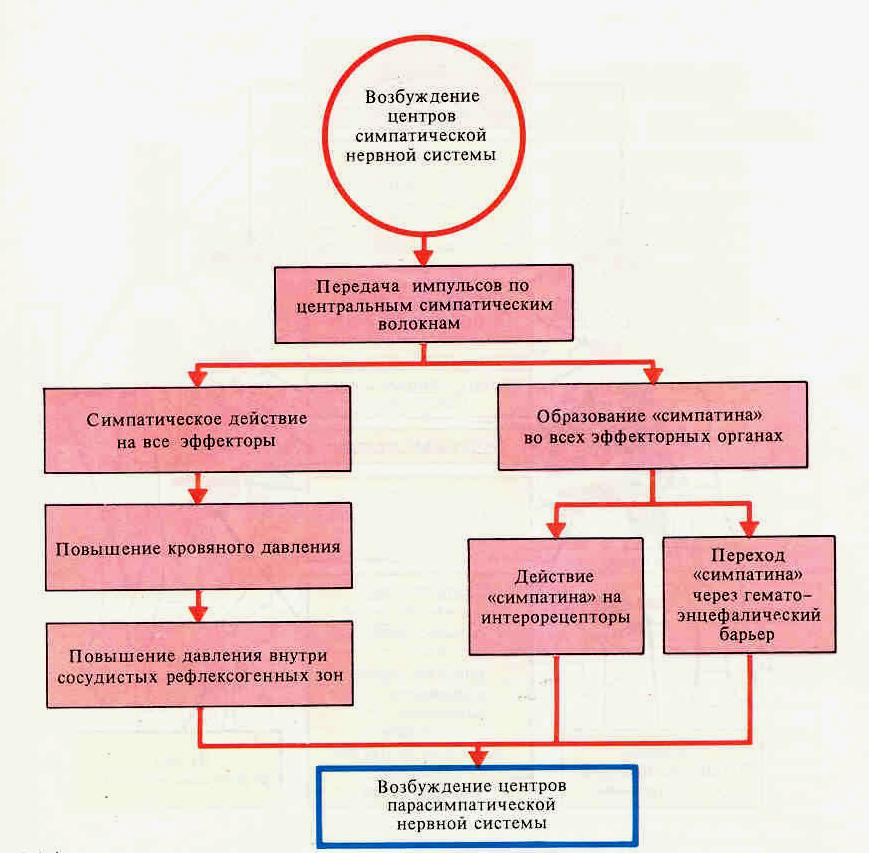 3.
МЕТОДЫ
исследования ВНД:
3.
МЕТОДЫ
исследования ВНД:
1) Метод условных рефлексов (в сочетании с различными дополнительными исследованиями или воздействиями).
Условный рефлекс – это выработанная в онтогенезе реакция организма на раздражитель, ранее индифферентный для этой реакции.
Основные правила выработки условных рефлексов:
– неоднократное совпадение во времени действия индифферентного раздражителя с безусловным;
– условный стимул должен предшествовать безусловному.
=> услоный рефлекс образуется на базе бзусловного.
Условия формирования условного рефлекса:
– оптимальное состояние организма (здоровье), и в первую очередь состояние коры больших полушарий;
– функциональное состояние нервного центра безусловного рефлекса;
– отсутствие посторонних сигналов как из внешней среды, так и от внутренних органов;
– оптимальное соотношение силы условного и безусловного раздражителей.
2) Электроэнцефалография – регистрация суммарной электрической активности мозга с поверхности головы.
Электроэнцефалограмма (ЭЭГ) – кривая, зарегистрированная при этом.
Электрокортикограмма – запись ЭЭГ с коры ГМ.
Регистрация ЭЭГ производится с помощью биполярных (оба активны) или униполярных (активный и индифферентный) электродов, накладываемых симметрично в лобных, центральных, теменных, височных и затылочных областях ГМ.
Основными анализируемыми параметрами ЭЭГ являются частота и амплитуда волновой активности.
На ЭЭГ регистрируются 4 основных физиологических ритма:
1. a-Ритм – (частота 8–13 Гц, амплитуду до 70 мкВ) – наблюдается в состоянии физического, интеллектуального и эмоционального покоя. Это упорядоченный регулярный ритм.
Его доминирование à ЭЭГ синхронизированная.
2. b-Ритм – (имеет нерегулярную частоту 14–30 Гц, низкую амплитуду – до 30 мкВ) – сменяет a-ритм при сенсорной стимуляции. Отражает высокий уровень функциональной активности мозга.
Смена a-ритма b-ритмом à десинхронизация ЭЭГ.
Объясняется активирующим влиянием на кору БП восходящей РФ ствола и лимбической системы.
3. q-Ритм – (частота 4–7 Гц, амплитуда до 200 мкВ) – у бодрствующего человека регистрируется обычно в передних областях мозга при длительном эмоциональном напряжении и почти всегда регистрируется в процессе развития фаз медленноволнового сна. Отчетливо регистрируется у детей, пребывающих в состоянии неудовольствия.
4. D-Ритм – (частота 0,5–3,0 Гц, амплитуда 200–300 мкВ) – эпизодически регистрируется во всех областях ГМ. Стабильно фиксируется во время глубокого медленноволнового сна.
Происхождение q- и D-ритм ЭЭГ связывают с активностью соответственно мостовой и бульбарной синхронизирующих систем ствола мозга.
3) Метод вызванных потенциалов (ВП) – регистрация колебания электрической активности, возникающего на ЭЭГ при однократном раздражении периферических рецепторов (зрительных, слуховых, тактильных).
4) Магнитоэнцефалография – мозг генерирует не только электрические, но и слабые магнитные волны
Преимущество этой методики: повышенная точность локализации очага корковой активности, т.к. сигналы от соседних участков не накладываются др. на др.
5) Компьютерная томография – через мозг пропускается тонкий пучок рентгеновских лучей, источник которого вращается вокруг головы в заданной плоскости; прошедшее через череп излучение измеряется сцинтилляционным счетчиком.
à рентгенографические изображения каждого участка мозга с различных точек.
6) Микроэлектродный метод – основан на подведении к одиночным нейронам микроэлектродов.
Позволяет изучать активность одиночных нейронов ЦНС; измерять мембранные потенциалы покоя, регистрировать постсинаптические потенциалы, а также потенциалы действия.
7) Метод молекулярной биологии – направлены на изучение роли молекул ДНК, РНК и др. биологически активных веществ в образовании условных рефлексов.
8) Методы холодового выключения структур ГМ – дают возможность визуализировать пространственно-временную мозаику электрических процессов мозга при образовании условного рефлекса в разных функциональных состояниях.
9) Стереотаксический метод – позволяет ввести электрод в различные подкорковые структуры ГМ à подготовить животное для хронического эксперимента.
10) Метод перерезки и выключения различных участков ЦНС – позволяет обратимо видоизменять активность мозга в целом и наблюдать за изменением условно-рефлекторного поведения.
11) Реоэнцефалография – основана на регистрации изменений сопротивления ткани мозга переменному току высокой частоты в зависимости от кровенаполнения à позволяет косвенно судить о величине общего кровенаполнения мозга, тонусе, эластичности его сосудов, состоянии венозного оттока.
12) Эхоэнцефалография – основана на свойстве ультразвука по-разному отражаться от структур мозга, его патологических образований, цереброспинальной жидкости, костей черепа и др.
à дает возможность: -определения размеров локализации тех или иных образований мозга; -оценивать скорость направления движения крови в сосудах, участвующих в кровоснабжении мозга.
24. Ретикулярная формация - скопление нервных клеток, расположенное в стволовой части мозга и представляющее собой как бы след нервных путей, соединяющих рецепторы органов чувств с участками коры мозга. Благодаря ратикулярной формации человек способен настораживаться, реагировать на незначительные изменения в окружающей среде. Она же обеспечивает возникновение ориентировочного рефлекса. с ее восходящими и нисходящими волокнами является нейрофизиологическим аппаратом, обеспечивающим одну из наиболее важных форм рефлекторной деятельности, известную под названием ориентировочного (или ориентировочно -- исследовательского) рефлекса. Его значение для понимания физиологических основ внимания особо велико.
Среди всех видов рефлекторной деятельности нужно выделить один, при котором поведение животного не возбуждается ни одним из перечисленных выше мотивов поведения и который не является ни пищевым, ни оборонительным, ни половым рефлексом. Основой этой деятельности является активная реакция животного на каждое изменение обстановки, которое и вызывает у животного общее оживление и ряд избирательных реакций, направленных на ознакомление с этими изменениями в ситуации. И.П.Павлов назвал этот вид рефлексов «ориентировочными рефлексами», или «рефлексами «что такое?».
Ориентировочный рефлекс выражается в ряде отчетливых электрофизиологических, сосудистых и двигательных реакций, появляющихся каждый раз, когда в обстановке, окружающей животное, возникает что-нибудь необычное или существенное. К этим реакциям относятся:поворот глаз и головы в сторону нового объекта; реакция настораживания или прислушивания.
Физиологические Механизмы Внимания
Физиологические механизмы внимания - нейрофизиологические структуры мозга разного уровня, обеспечивающие генерализованную и локальную активацию коры больших полушарий. При корковой активации, которая фиксируется на ЭЭГ в виде реакции десинхронизации (блокада альфа - ритма - ), происходит снижение порогов восприятия - и возрастание скорости протекания нервных процессов. При привлечении внимания - к стимулу происходит формирование локальных функциональных корковых объединений, соответствующих реализуемой деятельности - . При реализации какой - либо цели происходит избирательная активация областей коры и их функциональное объединение под контролем лобных отделов коры.
25. Условный рефлекс — это благоприобретенный рефлекс, свойственный отдельному индивиду (особи).[1] Возникают в течение жизни особи и не закрепляются генетически (не передаются по наследству). Возникают при определённых условиях и исчезают при их отсутствии. Формируются на базе безусловных рефлексов при участии высших отделов мозга. Условно-рефлекторные реакции зависят от прошлого опыта, от конкретных условий, в которых формируется условный рефлекс.
Существует бесчисленное множество условных рефлексов. При соблюдении соответствующих правил любой воспринимаемый раздражитель можно сделать стимулом, запускающим условный рефлекс (сигналом), а любую деятельность организма - его основой (подкреплением). По роду сигналов и подкреплений, а также по отношениям между ними созданы разные классификации условных рефлексов. Что касается изучения физиологического механизма временных связей, то здесь исследователям предстоит большая работа.
Определилась классификация условных рефлексов по следующим частным признакам: 1) обстоятельства образования, 2) род сигнала, 3) состав сигнала, 4) род подкрепления, 5) отношение во времени условного раздражителя и подкрепления.
Общие признаки условных рефлексов. Условный рефлекс а) является индивидуальным высшим приспособлением к меняющимся условиям жизни; б) осуществляется высшими отделами центральной нервной системы; в) приобретается путем временных нервных связей и утрачивается, если вызвавшие его условия среды изменились; г) представляет собой предупредительную сигнальную реакцию.
Итак, условный рефлекс - это приспособительная деятельность, осуществляемая высшими отделами центральной нервной системы путем образования временных связей между сигнальным раздражением и сигнализируемой реакцией.
Натуральные и искусственные условные рефлексы. В зависимости от характера сигнального раздражителя условные рефлексы делят на натуральные и искусственные.
Натуральными называют условные рефлексы, которые образуются в ответ на воздействие агентов, являющихся естественными признаками сигнализируемого безусловного раздражения.
Примером натурального условного пищевого рефлекса может служить выделение слюны у собаки на запах мяса. Этот рефлекс с неизбежностью образуется естественным путем в течение жизни собаки.
Искусственными называют условные рефлексы, которые образуются в ответ на воздействие агентов, не являющихся естественными признаками сигнализируемого безусловного раздражения. Примером искусственного условного рефлекса может служить выделение слюны у собаки на звук, метронома. В жизни этот звук не имеет никакого отношения к еде. Экспериментатор искусственно сделал его сигналом приема пищи.
Натуральные условные рефлексы природа вырабатывает из поколения в поколение у всех животных соответственно их образу жизни. В результате натуральные условные рефлексы легче образуются, скорее укрепляются и оказываются более прочными, чем искусственные.
Свойства условных рефлексов и механизмы их образования.
Механизм образования условных рефлексов
Согласно учению И. П. павлова (1903), временная связь образуется между корковым центром безусловного рефлекса и корковым центром анализатора, на рецепторы которого действует условный раздражитель, т.е. связь замыкается в коре большого мозга.
На первом этапе возбуждение иррадиирует в коре большого мозга, в том числе между двумя возбужденными центрами (от действия условного и безусловного раздражителей).
На втором этапе формируется синаптическое облегчение (проторение пути) между двумя возбужденными центрами.
На третьем этапе формируются два доминантных очага возбуждения (от действия условного и безусловного раздражителей). Таким образом, в основе замыкания временной связи лежит процесс доминантного взаимодействия между возбужденными центрами. Импульсы, вызываемые индифферентным (условным) сигналом с любого участка кожи и других органов чувств (глаз, ухо), поступают в кору большого мозга, обеспечивая там образование очага возбуждения. Если после индиффферентного сигнала подается пищевое подкрепление (подкормка), то возникает более мощный второй очаг возбуждения в коре больших полушарий, к которому направляется ранее возникшее и иррадиирующее по коре возбуждение. Неоднократное сочетание в опытах индифферентного (условного) сигнала и безусловного раздражителя (подкрепление) облегчает прохождение импульсов от коркового центра индифферентного сигнала к корковому представительству безусловного рефлекса – феномен облегчения (проторение пути). Образование временной связи в коре больших полушарий И. П. Павлов назвал замыканием новой условнорефлекторной дуги: теперь подача только условного сигнала приводит к возбуждению коркового центра безусловного рефлекса и возбуждает его, т.е. возникает рефлекс на условный раздражитель – условный рефлекс. Э. А. Асратян показал, что образование условных рефлексов с прямыми и обратными связями является общей закономерностью высшей нервной деятельности.
Следует отметить, что индифферентный сигнал вообще не является индифферентным, так как в начале своего применения вызывает собственную реакцию в виде ориентировочно – исследовательского рефлекса, который по мере повторения ослабляется. В процессе выработки условного рефлекса образуется двусторонняя связь между двумя центрами – корковым концом анализатора, на рецепторы которого действует условный раздражитель, и центром безусловного рефлекса, на базе которого вырабатывается условный рефлекс. Это было показано в опытах, где использовались два безусловных рефлекса: мигательный рефлекс, вызываемый струей воздуха у глаз, и безусловный пищевой.
При их сочетании выработался условный рефлекс: если подавали воздушную струю, возникал пищевой рефлекс, а при даче пищевого раздражителя отмечалось мигание.
Взаимосвязь левого и правого полушарий при выработке условных рефлексов обеспечивается комиссуральной системой (мозолистое тело, передняя, задняя, гиппокампальная и хабенулярная комиссуры, межбугровое сращение), которая анатомически соединяет два полушария головного мозга. Полученные результаты свидетельствуют о переносе возбуждения из одного полушария в другое. Например, если рефлекс вырабатывается на тактильное раздражение кожи одной стороны тела, он легко воспроизводится при таком же раздражении симметричных участков кожи другой стороны тела, т.е. в процессе выработки рефлекс «переносит» временную связь в другое полушарие. Предварительная перерезка мозолистого тела прекоащала (или сильно затрудняла) «перенос» условного рефлекса. Вместе с тем с помощью метода условных рефлексов было установлено, что перерезка всех комиссур больших полушарий полностью не исключает возможность межполушарного взаимодействия при выработке временной связи. Эти результаты свидетельствуют о том, что элементарное взаимодействие двух полушарий может происходить через четверохолмие и ретикулярную формацию ствола мозга.
На клеточном и молекулярном уровнях временная связь замыкается с помощью механизмов памяти. В начале выработки условного рефлекса связь осуществляется только с помощью механизмов кратковременной памяти – распространения возбуждения между двумя возбужденными корковыми центрами. По мере повторения действия условного и безусловного раздражителей и повторяющегося возбуждения соответствуюших центров, распространения возбуждения между ними кратковременная память переходит в долговременную (структурные и биохимические изменения в нейронах).
27. ВИДЫ УСЛОВНОГО ТОРМОЖЕНИЯ
Условное торможение образуется при жизни или вырабатывается в процессе дрессировки собак в виде отрицательных тормозных условных рефлексов. Возникает внутри условно-рефлекторной дуги через обратную афферентацию и по этой причине называется внутренним торможением. У собаки это торможение вырабатывается дрессировщиком в виде условного рефлекса и называется активным. Существует несколько видов условного торможения: угасательное, запаздывательное, дифференцировочное и условный тормоз.
Угасательное торможение развивается при неподкреплении условного раздражителя. Если, например, команду или жест «Рядом» не подкреплять безусловным раздражителем — рывком поводка, то с каждым разом условный рефлекс становится все слабее, и, наконец, совсем исчезает. Угасание условного рефлекса не связано с утомлением нервного центра при частом повторении. Если после полного угасания условного рефлекса сделать перерыв на несколько дней и вновь подкрепить условный раздражитель безусловным, то рефлекс вновь появляется и достигает прежней силы. В практике дрессировки можно наблюдать, когда угасший при неподкреплениях условный рефлекс через некоторое время сам по себе восстанавливается, т. е. угасательное торможение исчезает. Такое явление называется растормаживанием. Повторное угасание условного рефлекса происходит быстрее, чем первое, а последующие — быстрее и легче, чем предыдущие. Многократно повторяемое угасание может привести к полному исчезновению условного рефлекса. Очень слабые условные рефлексы после угасания могут не восстановиться. Закономерности угасательного торможения должны учитываться в методике организации и проведения занятий по дрессировке собак.
Запаздывательное торможение развивается в нервных центрах при отставлении подкрепления во времени от начала действия условного раздражителя и образуется на промежуток времени между действием условного и безусловного раздражителей. Оно лежит в основе образования недеятельной фазы условных рефлексов на время: отставленного, запаздывающего и следового. В дрессировке запаздывательное торможение используется для выработки выдержки при посадке, укладке, стоянии, подноске предметов, сторожевке и др. Благодаря развитию запаздывательного торможения, образовавшиеся условные рефлексы становятся более точными, они уравновешивают и дисциплинируют поведение собаки.
Дифференцировочное торможение проявляется при постоянном подкреплении одного условного раздражителя и неподкреплении других, сходных с ним. Выработка дифференцировочного торможения происходит постепенно, неравномерно, что свидетельствует о борьбе между развивающимся торможением и возбуждением.
Дифференцировка вырабатывается легче и быстрее при отдаленных сходствах раздражителей, при близких — образуется с трудом или совсем не образуется. Тонкую дифференцировку можно выработать после тренировки торможения сначала на более грубых, а затем на более тонких различиях сходных между собой раздражителей. На основе дифференцировочного торможения образуется дифференцировочный условный рефлекс, который обеспечивает способность собаки различать один раздражитель» из множества сходных с ним по внешнему виду, вкусу или запаху. Например, надрессированная собака может различать и выбирать по заданному запаху вещь из 10–12 предметов с запахами других людей. Дифференцировочный рефлекс легко растормаживается при действии на собаку посторонних раздражителей. У собаки это выражается в виде беспричинного возбуждения, перехватывания предметов, ошибочной выборки и др. Для выработки стойкого навыка дифференцировки необходимо многократное повторение упражнений по определенной методике.
Условный тормоз образуется или вырабатывается на условный раздражитель, способный затормаживать действие другого условного раздражителя. Например, запрещающая команда «фу» тормозит любой положительный условный рефлекс, проявляющийся на другой условный раздражитель. В практике дрессировки условный тормоз используется для устранения ненужных действий собаки, мешающих дрессировке и использованию ее на службе. Однако надо помнить, что применение условного тормоза вызывает торможение не только ненужного действия собаки, но и других положительных условных рефлексов.
Поэтому запрещающая команда «Фу» применяется только в необходимых случаях, а при выборке вещей и при работе по запаховому следу, как правило, не применяется.
28. Взаимодействие процессов возбуждения и торможения в коре больших полушарий мозга.
Важнейшей характеристикой деятельности нервных центров является постоянное взаимодействие процессов возбуждения и торможения как между разными центрами, так и в пределах каждого из них. Взаимосочетанная (реципрокная) иннервация мышц-антагонистов. Для сгибательного движения в суставе необходимо не только сокращение мышц-сгибателей, но и одновременное расслабление мышц- разгибателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей — процесс торможения. При возбуждении же центров разгибателей, наоборот, тормозятся центры сгибателей. Такие координационные взаимоотношения между моторными центрами спинного мозга были названы взаимосочетанной или реципрокной иннервацией мышц-антагонистов. Появление и усиление в нервных центрах процесса торможения при одновременном возбуждении других центров получило по аналогии с физическими процессами название индукции (в данном случае это одновременная индукция). В настоящее время выяснены механизмы проявления реципрокной иннервации. Афферентное раздражение (например, болевое раздражение рецепторов кожи) направляется не только по собственному рефлекторному пути к мотонейронам мышц-сгибателей, но и одновременно через коллатерали активирует тормозные клетки Рэншоу. Окончания этих клеток образуют тормозные синапсы на мотонейронах мышц-разгибателей, вызывая в них торможение. Реципрокные отношения между центрами мышц-антагонистов не являются постоянными и единственно возможными. В необходимых ситуациях (например, при фиксации суставов, при точностных движениях) они сменяются одновременным их возбуждением. В этом проявляется большая гибкость, целесообразность сложившихся в организме координации. Показано, что у человека во время ходьбы и бега основной формой координации являются реципрокные отношения, но помимо них имеются фазы одновременной активности мышц-антагонистов голеностопного и, особенно коленного и тазобедренного суставов. Длительность фаз одновременной активности увеличивается с повышением скорости перемещения. Взаимосочетанные (реципрокные) отношения характерны не только для моторных центров спинного мозга, но и для других центров. Еще в 1896 г. Н. Е. Введенский в опытах на животных наблюдал при раздражении двигательной зоны одного полушария коры головного мозга реципрокное торможение моторных центров другого полушария. При этом сокращение мышц одной половины тела сопровождалось расслаблением одноименных мышц другой. Реципрокные отношения формируются также при возникновении доминанты, когда при возбуждении одних центров с помощью сопряженного торможения выключается деятельность других, посторонних, нервных центров. Последовательная смена процессов возбуждения и торможения. Взаимоотношения процессов возбуждения и торможения в центральной нервной системе могут проявляться во времени в виде последовательной смены возбуждения и торможения в одних и тех же нервных центрах. «Возбуждение вслед за торможением» впервые наблюдал И. М. Сеченов. Он описал резкое усиление у лягушки рефлекторной деятельности после ее торможения сильным раздражителем: резкий прыжок с голосовой реакцией и восстановлением кожной чувствительности («рефлекс Сеченова»). Позднее было обнаружено «торможение вслед за возбуждением» (А. А. Ухтомский). После сильных ритмических раздражении лапки лягушки выключение раздражителя приводит к мгновенному расслаблению лапки—поднятая конечность падает, как плеть. Описанные взаимоотношения процессов возбуждения и торможения часто встречаются в коре больших полушарий при условнорефлекторной деятельности. Контрастное усиление одного процесса после другого в одном и том же нервном центре получило название последовательной индукции. Оно имеет большое значение при организации ритмической двигательной деятельности, обеспечивая попеременное сокращение и расслабление мышц.
29Аналитико-синтетическая деятельность коры головного мозга.
Деятельность коры головного мозга подчинена ряду принципов и законов. Основные из них впервые были установлены И.П.Павловым. В настоящее время некоторые положения павловского учения уточнены, развиты, а отдельные из них пересмотрены. Однако для овладения основами современной нейрофизиологии необходимо ознакомиться с фундаментальными положениями павловского учения.
Аналитико-синтетический принцип высшей нервной деятельности.
Как установлено И.П.Павловым, основным фундаментальным принципом работы коры больших полушарий головного мозга является аналитико-синтетический принцип. Ориентация в окружающей среде связана с вычленением отдельных ее свойств, сторон, признаков (анализ) и объединением, связью этих признаков с тем, что является полезным или вредным для организма (синтез). Синтез —это замыкание связей, а анализ —это все более тонкое отчленение одного раздражителя от другого.
Аналитико-синтетическая деятельность коры головного мозга осуществляется взаимодействием двух нервных процессов: возбуждения и торможения. Эти процессы подчинены следующим законам.
Закон иррадиации возбуждения. Очень сильные (так же, как и очень слабые) раздражители при длительном воздействии на организм вызывают иррадиацию —распространение возбуждения по значительной части коры больших полушарий.
Только оптимальные раздражители средней силы вызывают строго локализированные очаги возбуждения, что и является важнейшим условием успешной деятельности.
Закон концентрации возбуждения. Возбуждение, распространившееся из определенного пункта по другим зонам коры, с течением времени сосредоточивается в месте своего первичного возникновения.
Этот закон лежит в основе главного условия нашей деятельности —внимания (сосредоточенности сознания на определенных объектах деятельности).
При концентрации возбуждения в определенных участках коры мозга происходит его функциональное взаимодействие с торможением, это и обеспечивает нормальную аналитико-синтетическую деятельность.
Закон взаимной индукции нервных процессов. На периферии очага одного нервного процесса всегда возникает процесс с обратным знаком.
Если в одном участке коры сконцентрирован процесс возбуждения, то вокруг него индуктивно возникает процесс торможения. Чем интенсивнее сконцентрированное возбуждение тем интенсивнее и шире распространен процесс торможения.
Наряду с одновременной индукцией существует последовательная индукция нервных процессов —последовательная смена нервных процессов в одних и тех же участках мозга.
Только нормальное соотношение процессов возбуждения и торможения обеспечивает поведение, адекватное (соответствующее) окружающей среде.
Нарушение баланса между этими процессами, преобладание одного из них вызывает значительные нарушения в психической регуляции проведения.
Так, преобладание торможения, недостаточное взаимодействие его с возбуждением приводит к снижению активности организма. Преобладание возбуждения может выразиться в беспорядочной хаотической деятельности, излишней суетливости, снижающей результативность деятельности. Процесс торможения —это активный нервный процесс. Он ограничивает и направляет в определенное русло процесс возбуждения, содействует сосредоточению, концентрации возбуждения.
Торможение бывает внешним и внутренним. Так, если на животное внезапно подействует какой-либо новый сильный раздражитель, то прежняя деятельность животного в данный момент затормозится. Это внешнее (безусловное) торможение. В данном случае возникновение очага возбуждения по закону отрицательной индукции вызывает торможение других участков коры.
Одним из видов внутреннего или условного торможения является угасание условного рефлекса, если он не подкрепляется безусловным раздражителем (угасательное торможение). Этот вид торможения вызывает прекращение ранее выработанных реакций, если они в новых условиях становятся бесполезными.
Торможение возникает и при чрезмерном перевозбуждении мозга. Оно защищает нервные клетки от истощения. Этот вид торможения называется охранительным торможением.
На внутреннем виде торможения основана и аналитическая деятельность коры мозга, способность различать близкие по своим свойствам предметы и явления. Так, например, при выработке у животного условного рефлекса на эллипс оно вначале реагирует и на эллипс и на круг. Происходит генерализация, первичное обобщение сходных раздражителей. Но, если постоянно сопровождать предъявление эллипса пищевым раздражителем и не подкреплять предъявление круга, то животное постепенно начинает отчленять (дифференцировать) эллипс от круга (реакция на круг затормаживается). Этот вид торможения, лежащий в основе анализа, дифференцирования, называется дифференцировочным торможением. Оно уточняет действия животного, делает его более приспособленным к окружающей среде.
30. Доминанта и условный рефлекс
Под доминантой понимается более или менее устойчивый очаг повышенной возбудимости, чем бы он ни был вызван, причем вновь приходящие в центры возбуждения служат подтверждением возбуждения в очаге, в прочей НС прочно разлиты явления торможения.
А.А. Ухтомский определял доминанту как временно господствующий рефлекс, но это не единый центр возбуждения в ЦНС, а «комплекс определенных симптомов во всем организме», проявляющихся и в мышечной, и в секреторной, и в сосудистой деятельности. Не собственно очаг или центр возбуждения, а сложный комплекс связных рефлекторных дуг.
Центры, входящие в состав доминирующей функциональной констелляции, характеризуются: 1) повышенной возбудимостью; 2) стойкостью, инертностью возбуждения; 3) способностью к суммированию возбуждения и 4) сопряженно тормозящим действием на другие центры, не входящие в состав данной констелляции. Рассматривая процесс превращения доминанты из симптомокомплекса в общий механизм условнорефлекторной деятельности и поведения организма, A.A. Ухтомский подчеркивал; что морфологически разбросанные нервные центры не просто связаны в определенные констелляции, а функционально объединены единством действий.
В процессе развития формирования условного рефлекса отмечаются определенные ступени и фазы, развертывающиеся во времени в определенных структурах центральной нервной системы и имеющие свои пространственно-временные параметры. Но это не означает, что в процессе выработки условного рефлекса должны непременно прослеживаться все классические фазы его развития. Например, при «экстремальных» условиях ассоциативная связь образуется при одном двух сочетаниях, в результате чего условные связи могут сохраняться длительное время, если не всю жизнь. B.C. Русинов [7] считает, что механизмы условнорефлекторной деятельности, формирования новой условной связи часто протекают как единый процесс, включающий различные ступени развития. Такими ступенями являются простой суммационный рефлекс, сложный суммационный рефлекс -- доминанта и, наконец, условный рефлекс, который сам по себе может быть сложным в различной степени. Все три процесса рассматриваются как генетически взаимосвязанные, но неидентичные. «Доминанта "проходит" сначала суммационный рефлекс, прежде чем стать доминантой. Условный рефлекс является доминантой, прежде чем стать условным рефлексом. Такова схема соотношений нейрофизиологических механизмов доминанты и условного рефлекса», -- пишет B.C. Русинов [7; с. 8].
Нейрофизиологические механизмы образования условного рефлекса связаны с суммацией; в начальной стадии (генерализации) образования условный рефлекс аналогичен по физиологическим механизмам доминанте, но затем становится существенно иным. Доминанта образуется на всех и любых этажах центральной нервной системы (например, спинальная доминанта), и для ее образования не обязательно участие коры головного мозга. Для выработки условного рефлекса необходима кора больших полушарий (у высших животных). Они разные по степени своей сложности, по структурам центральной нервной системы, вовлеченным в процесс, по системам связи и значениям этих связей для поведения [3; с. 20].
Изучение свойств доминанты показало, что нейроны доминантного очага характеризуются обширным генерализованным рецептивным полем и способны отвечать на любые раздражители, не имеющие к ней прямого отношения. Нейроны доминантного очага «знают» свой (специфический) эфферентный выход, «готовы» осуществить эффект, но «не знают», на какой раздражитель необходимо осуществить это эффекторное действие. Любой афферентный импульсный «толчок» может стать эффективным.
Таким образом, направление движения нервных процессов при формировании условного рефлекса связано с механизмом и свойствами доминанты, для которой характерны не только высокая возбудимость, но и способность стойко удерживать свое возбуждение, суммировать его за счет последовательно приходящих импульсов от других нервных центров. Возникший в коре очаг возбуждения от условного стимула может иррадиировать во все стороны, однако при наличии сильного возбуждения, которое обусловлено действием подкрепления (несущего побуждающее безусловно-рефлекторное действие), иррадиация будет «направляться» в сторону этого более сильного очага.
Связь условного рефлекса и доминанты
В.С. Русинов выделяет несколько стадий образования условного рефлекса: простой суммационный рефлекс, доминанта и собственной условный рефлекс (УР). В начальной стадии образования (генерализации) УР = Д, а затем происходит дифференциация. Исследования Русиновым свойств Д - создание искусственного очага постоянного возбуждения. Нейроны, входящие в состав доминанты им генерализованное рецептивное поле и способны отвечать на раздражители, которые не имеют прямого отношения к данной доминанте например, при возбуждении участка коры ГМ (проекции какой-либо конечности) при предъявлении индифферентных стимулов (световой, звуковой) - ответная реакция конечности. Доминанта способна усваивать ритм афферентных раздражителей (ритмические движения конечности). Возникновение доминанты возможно только при неоднократной стимуляции соответствующих РД [9; c. 133].
31. Память — основа приобретения и сохранения человеком индивидуального опыта, она позволяет приспосабливаться к меняющимся условиям жизни. Можно дифференцировать память на механическую (фиксация фактов, цифр, слов, фамилий), абстрактно-логическую, двигательную. Зрительная память, например, была прек-расно развита у И. Левитана, слуховая — у В. А. Моцарта. У слепых преобладает осязательная память. Люди, легко усваивающие двигательные навыки, в большинстве своем потенциально хорошие спортсмены. И тем не менее при всем различии видов памяти ее механизмы едины.
Специалисты определяют память как способность головного мозга запечатлевать воспринимаемую информацию, хранить и воспроизводить следы действовавших ранее возбуждений. Образование следа, или, как теперь говорят, энграммы, памяти происходит в два этапа. Вначале вся информация, которая отражает происходящие вокруг нас события, воспринимается рецепторами органов чувств. Они кодируют ее в виде электрических импульсов различной частоты и амплитуды. Такие импульсы поступают в сеть контактирующих друг с другом нервных клеток мозга, образующих замкнутый круг. Здесь импульсы циркулируют от 2 до 12 минут, оставляя наиболее заметные следы в области контактов между нейронами — в синапсах.
Под памятью понимают свойство живых систем, в частности ЦНС, воспринимать, фиксировать, хранить и воспроизводить следы ранее действующих раздражителей. Иными словами: В результате жизнедеятельности, обучения организм (человек, животное) изменяется, иначе реагирует на те же раздражители, что является наличием следов обучения — памяти. Объем памяти, длительность и надежность хранения информации, как и способность к восприятию сложных сигналов среды и выработке адекватных реакций возрастают в ходе эволюции по мере увеличения числа нервных клеток мозга — нейронов и усложнения его структуры. Память характерна не только для ЦНС; существует генетическая память, иммунная память. Нервная память, осуществляемая ЦНС, характеризуется тем, что хранение информации о прошедших событиях внешнего мира и об ответных реакциях организма на эти события используется организмом для построения модели текущего или будущего поведения.
В формировании и осуществлении высших функций мозга очень важное значение имеет общебиологическое свойство фиксации, хранения и воспроизведения информации, объединяемое понятием память. Память как основа процессов обучения и мышления включает в себя четыре тесно связанных между собой процесса: запоминание, хранение, узнавание, воспроизведение. На протяжении жизни человека его память становится вместилищем огромного количества информации: в течение 60 лет активной творческой деятельности человек способен воспринять 1013— 10 бит информации, из которой реально используется не более 5—10 %. Это указывает на значительную избыточность памяти и важное значение не только процессов памяти, но и процесса забывания. Не все, что воспринимается, переживается или делается человеком, сохраняется в памяти, значительная часть воспринятой информации со временем забывается. Забывание проявляется в невозможности узнать, припомнить что-либо или в виде ошибочного узнавания, припоминания. Причиной забывания могут стать разные факторы, связанные как с самим материалом, его восприятием, так и с отрицательными влияниями других раздражителей, действующих непосредственно вслед за заучиванием (феномен ретроактивного торможения, угнетения памяти). Процесс забывания в значительной мере зависит от биологического значения воспринимаемой информации, вида и характера памяти. Забывание в ряде случаев может носить положительный характер, например память на отрицательные сигналы, неприятные события. В этом справедливость мудрого восточного изречения: «Счастью память отрада, горю забвение друг».
В результате процесса научения возникают физические, химические и морфологические изменения в нервных структурах, которые сохраняются некоторое время и оказывают существенное влияние на осуществляемые организмом рефлекторные реакции. Совокупность таких структурно-функциональных изменений в нервных образованиях, известная под названием «энграмма» (след) действующих раздражителей становится важным фактором, определяющим все разнообразие приспособительного адаптивного поведения организма.
Кратковременная память
Кратковременная память удерживает материал иного типа, нежели непосредственный отпечаток сенсорной информации. В данном случае удерживаемая информация представляет собой не полное отображение событий, которые произошли на сенсорном уровне, а непосредственную интерпретацию этих событий. Например, если при вас произнесли какую-то фразу, вы запомните не столько составляющие ее звуки, сколько слова. Обычно запоминается 5-6 последних единиц из предъявленного материала. Сделав сознательное усилие, вновь и вновь повторяя материал, можно удерживать его в кратковременной памяти на неопределенно долгое время.
Долговременная память.
Существует явное и убедительное различие между памятью о только что случившемся событии и событиях далекого прошлого. Долговременная память — наиболее важная и наиболее сложная из систем памяти. Емкость первых названных систем памяти очень ограничена: первая состоит несколько десятых секунд, вторая — несколько единиц хранения. Однако какие-то границы объема долговременной памяти все же существуют, так как мозг является конечным устройством. Он состоит из 10 млрд нейронов и каждый способен удерживать существенное количество информации. Причем оно настолько велико, что практически можно считать, что емкость памяти человеческого мозга не ограничена. Все, что удерживается на протяжении более чем нескольких минут, должно находиться в системе долговременной памяти.
Главный источник трудностей, связанных с долговременной памятью, — это проблема поиска информации. Количество информации, содержащейся в памяти, очень велико, и поэтому сопряжено с серьезными трудностями. Тем не менее, отыскать необходимое удается быстро.
32. Потребности и мотивации
Организм животных и человека время от времени испытывает необходимость в каких-либо веществах, продуктах, а также ситуациях или информации, нужных ему для организации текущего или будущего поведения, что, в конечном итоге, обеспечивает сохранение его жизни и возможность оставить потомство. Такая необходимость в чем-либо называется потребностью. Например, если мы с вами или наши собаки начинаем испытывать голод, то для его удовлетворения совершаем соответствующее пищедобывающее поведение - кто-то идет в магазин, кто-то открывает холодильник, а кто-то и стягивает со стола оставленный без присмотра батон колбасы. Или в другом случае, чтобы собаку заставить сесть, мы довольно сильно нажимаем ей на круп в области крестца, вызывая тем самым неприятные ощущения от давления и, естественно, потребность избавиться от него - в итоге собака садиться. Считается, что потребность является причиной поведения и беспричинного поведения не бывает.
Однако, возникающая потребность вызывает изменение поведения только достигая определенного уровня. Например, при начинающемся недостатке питательных веществ, понижается содержание глюкозы в крови, что регистрируется специальными рецепторами. Незначительный - быстро восстанавливается за счет оперативных резервов организма, то есть гомеостатически, и внешне никак не проявляется. По мере истощения резервов, организм начинает испытывать специфическое чувство голода, которое заставляет его изменить поведение в сторону удовлетворения аппетита. При этом в центральной нервной системе развиваются такое состояние и такие процессы, которые организуют и поддерживают нужное поведение. Это состояние и соответствующая ему деятельность нервной системы называются мотивацией
потребности на 3 основные группы:
1. Потребности жизнеобеспечения (витальные потребности) направлены на сохранение организма. К ним относят пищевую, питьевую, оборонительную потребности, потребность в сне, двигательной нагрузке, положительных ощущениях и тому подобное. Неудовлетворение какой-либо витальной потребности ведет к гибели животного.
2. социальные потребности обеспечивают сосуществование животного с другими животными и способствуют не только сохранению индивидуума, но и сохранению вида. Различают половые, родительские, территориальные потребности, а также потребность в группе и формировании групповых иерархических взаимоотношений. Зоосоциальные потребности для своей реализации требуют участия социальных партнеров своего или другого вида.
3. Потребности саморазвития ориентированы на будущее и их значение заключается в подготовке организма к возможным условиям существования. К потребностям саморазвития относят потребность в свободе, имитационную (подражательную) потребность, потребность в игре, потребность в новой информации и др.
Социальные (социоэмоциональные) потребности человека включают в себя необходимость принадлежать к социальной группе, занимать в ней определенное место, пользоваться уважением и любовью окружающих, т. е. занимать определенную социальную нишу (территорию).
К этой группе относятся также потребности в самоутверждении, в достижении социальных успехов, потребность быть лидером или наоборот - быть ведомым, потребность следовать нормам, принятым в обществе, потребность в одобрении со стороны окружающих, потребности в идеологии и в культуре.
Социальные и биологические потребности делятся на две ветви - потребности «для себя» и потребности «для других».
Биологические потребности «для себя» обеспечивают выживание индивида самого по себе. Биологические потребности «для других» обслуживают выживание вида в целом как общности биологических особей. Предпосылкой развития потребностей «для других» в животном мире являются родительский инстинкт, групповая забота о молодняке, наблюдаемая у многих видов животных, а также способность к эмоциональному резонансу. У человека биологические потребности «для других» проявляются в основном как забота о близких, родных, членах семьи, об их благополучии.
Социальные потребности «для себя» обусловлены представлением человека о своих правах, под их влиянием человек стремится улучшить свое общественное положение. Под влиянием социальных потребностей «для других» человек заботится о справедливости по отношению к другим людям, борется за улучшение социального положения какой-либо группы людей, действует, исходя из представлений о своих обязанностях. Общественно-историческая норма удовлетворения потребности «для других» усваивается человеком в процессе его воспттания и воспринимается им как совесть.
Когда человек занят удовлетворением потребности «для себя», он склонен видеть в другом человеке врага или конкурента. Потребности «для других» предполагают видение в другом друга, соратника, помощника. Потребности «для других» и «для себя» обуславливают альтруистическое и эгоистическое поведение индивида. И то, и другое имеет свое эволюционное значение, способствуя выживанию человечества как целого.
Идеальные (духовные) потребности - это потребности в новой информации, в познании окружающего мира, в поиске своего места в нем, в познании смысла и назначения своего существования на земле, в самовыражении, самореализации, творчестве, эстетическая потребность, потребность освоить некую «идеальную территорию».
Базисной в этой группе является потребность в новой информации, которая проявляется уже у грудных детей. И если уже в этом возрасте не обеспечить ребенку возможность такую информацию получать, то это может в будущем отрицательно сказаться на развитии его интеллектуальных возможностей.
Потребность в познании происходит от присущей всему живому потребности в информации, наряду с потребностью в притоке вещества и энергии. Потребность в новой информации существует у человека независимо от ее практического значения для удовлетворения биологических или социальных потребностей. Она проявляется не только как стремление к новому, неизвестному, но и как стремление это новое объяснить, упорядочить, познать присущие ему закономерности. Слишком много новой информации может действовать на человека пугающе. Объясняя же мир, создавая разные модели, описания мира, человек снимает свой страх перед загадочностью и непредсказуемостью жизни.
Биологические, социальные и идеальные потребности можно разделить на две группы: потребности сохранения (нужды) и потребности развития (роста).
Потребности нужды удовлетворяются в пределах существующих общественно-исторических норм, их неудовлетворение, как правило, сопровождается отрицательными эмоциями, переживаемыми как тревога, ощущение нависшей угрозы, боязнь наказания.
Удовлетворение потребностей развития связано с выходом за пределы существующих общественных норм удовлетворения данной потребности, со стремлением улучшить свое положение. Поэтому удовлетворение потребностей развития вызывает у человека положительные эмоции. Угроза неудовлетворения потребностей развития вызывает у человека тоску, раздражение, вызванные отсутствием удач.