

43.Энергия активации реакции (процесса). Экспериментальной определение величины энергии активации.(см №42 тоже)
Зависимость скорости реакции от температуры дает уравнение Аррениуса:
, (10.16)
где k — константа скорости реакции, R — универсальная газовая постоянная, Е — энергия активации химической реакции. В случае простых реакций величина Е показывает, какой минимальной (избыточной по сравнению со средней) энергией в расчете на 1 моль должны обладать реагирующие частицы, чтобы они могли вступить в химическую реакцию. В случае сложных реакций величина Е называется эмпирической или кажущейся энергией активации и в общем случае зависит от энергий активации отдельных стадий данной реакции.
Проинтегрировав уравнение (10.16), получим уравнение Аррениуса в интегральной форме:
, (10.17) Или ,
где А — предэкспоненциальный множитель. Физический смысл А в случае простых реакций: мономолекулярных — это частота колебаний по разрываемой связи (А 1013 сек-1), бимолекулярных — величина А пропорциональна общему числу столкновений между молекулами реагирующих веществ (А 10-10 10-11 см3/(мол-л·сек).
Проинтегрировав ур-е (10.16) в пределах температур от Т1 до Т2, получим:
![]() . (10.18)
. (10.18)
Э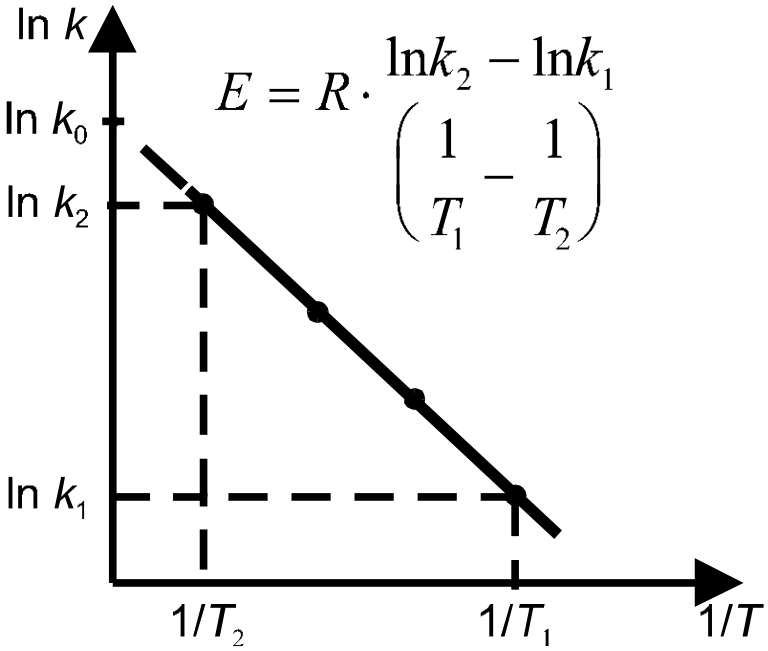 нергию
активации можно определить как
аналитически по уравнению (10.18), так и
графическим методом. Для этого необходимо
знать ряд констант скоростей при разных
температурах. Если реакция подчиняется
уравнению Аррениуса, то зависимость
lnk от 1/T должна выражаться прямой
линией, что следует из уравнения (10.17)
(рис. 7).
нергию
активации можно определить как
аналитически по уравнению (10.18), так и
графическим методом. Для этого необходимо
знать ряд констант скоростей при разных
температурах. Если реакция подчиняется
уравнению Аррениуса, то зависимость
lnk от 1/T должна выражаться прямой
линией, что следует из уравнения (10.17)
(рис. 7).
Рис. 7 К расчету энергии активации реакции по температурной зависимости константы скорости. Для очень большого числа реакций энергия активации находится в пределах от 60 до 240 кДж/моль, т.е. примерно соответствует энергиям химических связей. Энерг актив. Связана с Q10: Е=0,46T1*T2lgQ10
Энергии активации большинства биологических процессов того же порядка, что и для химических реакций. Они группируются в основном у трех величин — 8, 12 и 18 ккал/молъ. Однако в процессах разрушения клеток различными токсическими факторами энергии активации очень велики — порядка 150 ккал/молъ. Такая же высокая энергия активации характерна для реакций денатурации белка. Например, для свертывания гемоглобина - 60 ккал/молъ, для гемолиза при действии горячей воды энергия активации равна 64 ккал/молъ, для бактериолиза В. paratyphosus в феноле — 48,6 ккал/молъ.
Для ферментативных реакций характерны более низкие значения энергии активации. Согласно современным представлениям, ускорение реакции в присутствии ферментов связано с тем, что фермент образует комплекс с субстратом. При этом конфигурация электронного облака реагирующей молекулы изменяется так, что облегчается ее вступление в реакцию, и, следовательно, уменьшается энергия активации реакции. Например, энергия активации гидролиза сахарозы при действии амилазы равна 11 ккал/молъ, а при действии кислот — 25,6 ккал/молъ.
Для фотохимических процессов в клетках Eакт очень низка, например 500—1000 кал/моль для процессов, протекающих в сетчатке глаза.
Энергия активации связана с коэффициентом Вант-Гоффа следующим соотношением:
Eакт = 0,46Т1 · Т2lgQ10. |
|
Возрастание скорости реакции при увеличении температуры происходит тем быстрее, чем больше энергия активации реакции. Поэтому, если процесс осуществляется конкурирующими реакциями, скорость которых по-разному зависит от температуры, при низких температурах основной выход продукта будет идти за счет реакции с малой энергией активации, а при повышении температуры будет возрастать роль реакции, имеющей большую энергию активации. В случае последовательных реакций скорость суммарного процесса будет определяться скоростью наиболее медленно протекающей реакции. При низкой температуре более медленно протекающей реакцией может быть реакция с большей энергией активации, а при повышении температуры — реакция с меньшей энергией активации.