

Функции днк
1. Днк является носителем генетической информации. Функция обеспечивается фактом существования генетического кода.
2. Воспроизведение и передача генетической информации в поколениях клеток и организмов. Функция обеспечивается процессом репликации.
3. Реализация генетической информации в виде белков, а также любых других соединений, образующихся с помощью белков-ферментов. Функция обеспечивается процессами транскрипции и трансляции.
![]()
Определение:
белки
- это нерегулярные полимеры, мономерами
которых являются
![]() L-аминокислоты.
L-аминокислоты.
![]()
Аминокислоты
В природе существуют две формы стереоизомеров: L (левовращающие) и D (правовращающие). Помимо L - аминокислот, входящих в белки, в организме есть и D-аминокислоты, которые в белки не включаются.
Общая формула аминокислоты показана на рисунке.
|
|
Она верна для 19 из 20 аминокислот, встречающихся в белках. В состав белков, кроме этих 19 аминокислот, входит одна иминокислота - пролин. Во
всех аминокислотах имеется
|
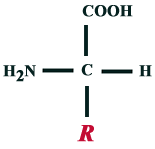
![]()
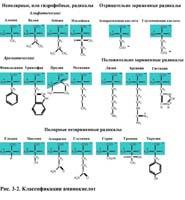
Классификация аминокислот, входящих в состав белков, по принципу полярности (неполярности) радикала
|
|
1. Неполярные или гидрофобные радикалы. Алифатические - аланин, валин, лейцин, изолейцин. Серусодержащий метионин. Ароматические - фенилаланин, триптофан. Иминокислота пролин. 2. Полярные, но незаряженные радикалы. Глицин. Оксиаминокислоты - серин, треонин, тирозин. Содержащий сульфгидрильную группу цистеин. Содержащие амидную группу: аспарагин, глутамин. 3. Отрицательно заряженные радикалы. Аспарагиновая кислота, глутаминовая кислота. 4. Положительно заряженные радикалы. Лизин, аргинин, гистидин. |
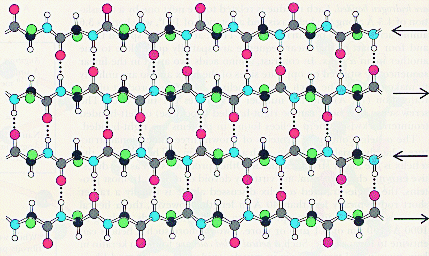
Первичная структура белка
![]()
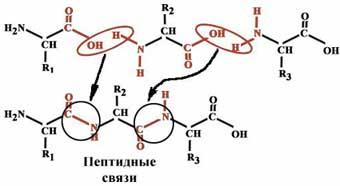
Определение: первичная структура белка - это последовательность расположения аминокислотных остатков в полипептидной цепи.
![]()
|
|
Аминокислоты соединяются в полипептид с помощью ковалентных пептидных (амидных) связей. |
|
У трипептида, состоящего из трех разных аминокислот, возможно 3! = 6 различных первичных структур. У олигопептида, состоящего из двадцати разных аминокислот, разнообразие первичных структур 20!, это ≈ 2*1018. Разнообразие первичных структур среднего по размеру белка (примерно 500 аминокислот) составляет уже ≈ 20500 вариантов (если все аминокислоты представлены в эквимолярных соотношениях). |
|
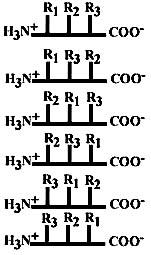
На Земле не было, нет и не будет двух людей с полностью одинаковым набором белков.
![]()
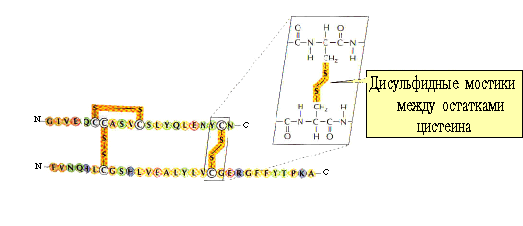
Третичная структура белка
![]()
Определение: третичная структура белка - это пространственная конформация полипептида, имеющего вторичную структуру, и обусловленная взаимодействиями между радикалами.
![]()
Существует четыре типа взаимодействий между радикалами:
|
1. Ковалентные связи между остатками двух цистеинов(дисульфидные мостики). |
|
|
2. Ионные (электростатические) взаимодействия между противоположно заряженными аминокислотными остатками (три радикала со знаком "+" и два со знаком "-"). Например, положительно заряженная ε-аминогруппа лизина (-NH3+ ) притягивается отрицательно заряженной карбоксильной группой - (СОО-) глутаминовой или аспарагиновой кислоты. |
|
|
3. Водородные связи. Участвуют все аминокислоты, имеющие гидроксильные, амидные или карбоксильные группы. |
|
|
4. Гидрофобные взаимодействия . Образуются между неполярными радикалами в водной среде. Участвуют 8 аминокислот (первый класс).
|
|

Третичная структура полностью задается первичной.
![]()
Определяющими являются гидрофобные взаимодействия в силу неизбирательности (неспецифичности) и многочисленности.
Гидрофобное ядро существует у большинства белков.