

Пищеварительная система млекопитающих.
Пищеварительная система состоит из 4 отделов(ротоглоточная полость, пищевод, желудок, кишечник)
Характерно общее удлинение пищеварительного трактапо сравнению с другими группами позвоночных и его более развитая дифференцировка, также происходит значительное развитиепищеварительных желёз
Развивается симбиотическое пищеварение
Ротовое отверстие окружено мягкими губами
Зубыдифференцированы по функциям, сидят в альвеолах
При попадании в ротовую полость пища пережёвывается зубами и смачивается слюной(содержит ферменты), из полости рта попадает вглотку, а оттуда впищеводи желудок
Желудоку большинства млекопитающих простой (однокамерный), но у нек. имеется несколько отделов (желудков)
Кишечникразделён на тонкий и толстый (к последнему присоединена слепая кишка)
Большая часть переваривается в тонком кишечнике, через стенки которого пит.вещ-ва всасываются в кровь, остатки поступают втолстый кишечникгде происходят бродильные процессы с участием бактерий
Непереваренные остатки выводятся через анальное отверстие
Присутствуют пищеварительные железы, облегчающие пищеварение и выделяющие различные ферменты
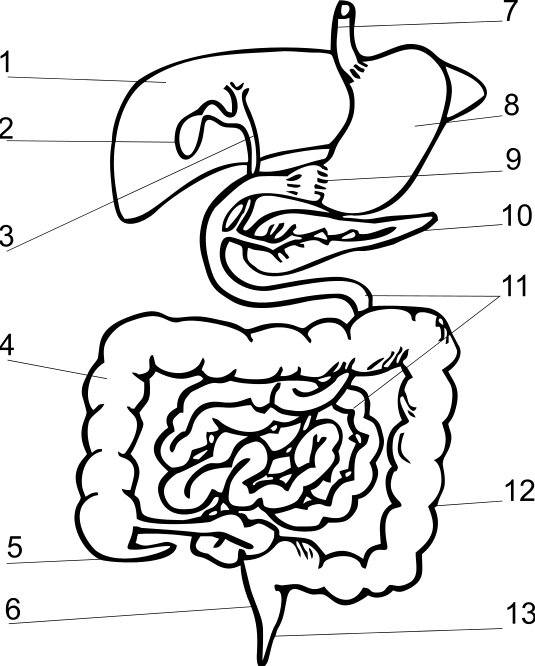
Пищеварительная система млекопитающего.
1 — печень,
2 — жёлчный пузырь,
3 — жёлчный проток,
4, 12 — толстая кишка,
5 — слепая кишка,
6 — прямая кишка,
7 — пищевод,
8 — желудок,
9 — привратник желудка,
10 — поджелудочная железа,
11 — тонкая кишка,
13 — анальное отверстие.
Происхождение млекопитающих.
Млекопитающие появились ещё в верхнем карбонеот звероподобных рептилий, которые обладали рядом примитивных признаков: амфицельный позвонки, подвижные шейные и поясничные ребра и т.д.
Длительное время звероподобные рептилии существовали мало, отличаясь от своих предков и сохраняли многие черты организации амфибий (этим можно объяснить большое количество у млекопитающих кожных желез)
Согласно современным представлениям, млекопитающие произошли от синапсид из группы цинодонтов, выделившись в конце триасового периода. Наиболее продвинутые цинодонты уже сильно напоминали млекопитающих — как, например, Oligokyphus из семейства Tritylodontidae с его развитым шерстным покровом, живший в позднем триасе и ранней юре.
Тогда же имели место и начальная дивергенция млекопитающих: в отложениях позднего триаса найдены ископаемые остатки кюнеотерия и харамиид. Последних обычно рассматривают как ранних представителей подкласса (или инфракласса) аллотериев, куда включают также многобугорчатых — наиболее разнообразный и многочисленный из мезозойских отрядов млекопитающих, просуществовавший свыше 100 млн лет, что касается морганукодонтов, то они по своему облику и строению чрезвычайно близки к предполагаемому предку всех более поздних млекопитающих.
В верхнем триасе выделились и другие основные линии млекопитающих, известные останки которых относятся к более позднему времени: линия, включающая однопроходных; линия три конодонтов (юра — мел); наконец, та линия, к которой принадлежат сумчатые и плацентарные, отделившиеся друг от друга в юрском периоде.
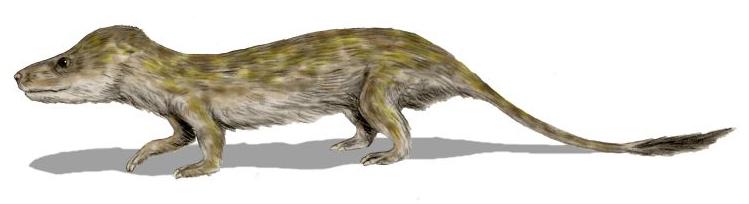
Цинодонт Oligokyphus (современная реконструкция)

Morganucodon — триасовый прототип позднейших млекопитающих
Происхождение птиц.
Происхождение птиц долгое время оставалось предметом оживлённых дискуссий. За обозримый период времени было выдвинуто несколько научных версий происхождения и родственных связей птиц и возникновения у них полёта, причём в течение более сотни лет они были чисто гипотетическими.
Впервые теория эволюции птиц от рептилий возникла после открытия в 1860 году в Германии окаменелых останков археоптерикса — животного, жившего около 150 млн лет назад в верхней юре. Он обладал характеристиками типичного пресмыкающегося — особым строением таза и рёбер, зубами, когтистыми лапами и длинным, как у ящерицы, хвостом. При этом окаменелости имели хорошо сохранившиеся отпечатки маховых крыльев, аналогичные таковым у современных птиц. На протяжении многих десятилетий история развития птиц рассматривалась как эволюция группы, развивавшейся от археоптерикса.
Именно на его изучении и основывались все первые гипотезы и теории о происхождении и родственных связях птиц: теория древесного («с деревьев вниз», Marsh, 1877) и теория бегательного («с земли вверх», Williston, 1879) возникновения полёта у птиц. Соответственно этим представлениям трактовалось и происхождение самих птиц — от триасовых текодонтов (архозавроморф) при древесной теории или от юрских бегающих тероподных динозавров при наземной теории.
В настоящее время археоптерикса уже не рассматривают в качестве общего предка всех современных птиц. Однако он, вероятно, имеет близкое родство с их настоящим предком. Точное положение археоптерикса в эволюционном дереве определить трудно. Согласно кладистическому анализу китайских палеонтологов археоптерикс может представлять собой параллельную тупиковую ветвь на общем стволе динозавров. Однако более тщательный филогенетический анализ не подтвердил размещение археоптерикса среди дейнонихозавров, и поэтому его продолжают считать древнейшей и наиболее примитивной птицей (в составе группы Avialae).
Тем не менее, были обнаружены более древние ископаемые останки, которые также могут быть отнесены к кладу Avialae, хотя на данный момент они рассматриваются как динозавры: Anchiornis, Xiaotingia и Aurornis.


Археоптерикс (реконструкция) и его археологический отпечаток
Происхождение рептилий.
Остатки наиболее древних пресмыкающихся известны из верхнего карбона(прим. 300 млн. лет назад)
Однако обособление рептилий должно было происходить ещё немного раньше (ок. 320 млн. лет.), когда от примитивных стегоцефалов, обособлялись формы, видимо обладавшие большей наземностью
В среднем карбоне от подобных форм возникает новая ветвь – сеймуриоморф, они занимают переходное положение от земноводных к млекопитающим, имея при этом множество черт рептилий
Когда сложился присущий амниотам характер размножения и развития яйца в воздушной среде пока не ясно, но можно полагать, что произошло это в карбоне при становлении котилозавров. Крыша их черепа была сплошной, завершилось образование атланта и эпистрофея
Основной предковой группой, давшей начало все многообразие современных пресмыкающихся были котилозавры

Происхождение хордовых животных.
Попытки выработать эволюционные отношения хордовых привели к рождению нескольких гипотез. В настоящее время достигнут консенсус в том, что хордовые являются потомками одного общего предка, который сам является хордовым, а ближайшими родственниками позвоночных (лат. Vertebrata) являются головохордовые (лат. Cephalochordata).
Все обнаруженные окаменелости ископаемых хордовых были найдены в раннем кембрии и включают в себя два вида позвоночных, классифицируемых как рыба. Поскольку окаменелости хордовых плохо сохранились, только метод молекулярной филогенетики предполагает разумную перспективу исследования их возникновения. Однако, использование метода молекулярной филогенетики для изучения эволюционных процессов является спорным.
Билатеральные животные делятся на два больших таксона —первичноротые и вторичноротые. Хордовые относятся ко вторичноротым. Весьма вероятно, что ископаемое кимберелла, жившая 555 млн лет назад, принадлежала к первичноротым. Жившая 549—543 млн лет назад в эдиакарии эрниетта была уже явно вторичноротым животным. Таким образом, первичноротые и вторичноротые должны были разделиться до времени существования этих животных, то есть до начала кембрийского периода.
Первые известные ископаемые двух близких к хордовым групп —иглокожих и полухордовых — обнаруживаются с раннего и среднего кембрия, соответственно. С другой стороны, ископаемые других хордовых весьма редки, поскольку у них нет твёрдых частей тела.
Исследования родственных отношений хордовых начались с 90-х годов XIX века. Они основывались на анатомических, эмбриологических и палеонтологических данных и приводили к разным филогенетическим деревьям. Некоторое время ближайшими родственниками хордовых считали полухордовых, но ныне эта гипотеза отвергнута. Сочетание данных классических методов с данными по анализу последовательностей генов рРНК привело к появлению гипотезы о том, что оболочники — живые представители группы, базальной для других вторичноротых. Относительно взаимоотношений внутри хордовых некоторые учёные считают, что наиболее близкими родственниками позвоночных являются головохордовые, но есть основания считать таковыми оболочников.
Время происхождения хордовых, на основании метода молекулярных часов, было оценено в 896 млн лет.

Размножение и развитие рептилий
Пресмыкающиеся — раздельнополые животные, двуполое размножение.
Половая система самцов состоит из пары семенников, которые расположены по бокам поясничного отдела позвоночника. От каждого семенника отходит семенной канал, который впадает в вольфов канал. С появлением туловищной почки у пресмыкающихся вольфов канал у самцов выступает лишь как семяпровод и полностью отсутствует у самок. Вольфов канал открывается в клоаку, образуя семенной пузырёк.
Половая система самок представлена яичниками, которые подвешены на брыжейке к спинной стороне полости тела по бокам позвоночника. Яйцеводы (мюллеровы каналы) также подвешены на брыжейке. В переднюю часть полости тела яйцеводы открываются щелевидными отверстиями — воронками. Нижний конец яйцеводов открывается в нижний отдел клоаки на её спинной стороне.
Развитие - оплодотворение внутреннее. Развитие зародыша происходит в яйце с кожистой или известковой оболочкой, наряду с этим встречается яйцеживорождение и (реже) истинное живорождение. У пресмыкающихся прямое постэмбриональное развитие.
Для многих представителей характерна забота о потомстве, в частности самки крокодилов переносят потомство от места кладки к водоёмам в ротовой полости, хотя в некоторых случаях могут съесть детёныша.

