

Физиология_сенсорных_систем
.PDF11
Корковые представительства анализаторов
Рис. 4. Корковые представительства анализаторов слухового восприятия на определенный частотный диапазон – тембр голоса).
Окончательная переработка информации о действующем стимуле происходит в сенсорных областях коры (рис. 4).
Корковые концы анализаторов называют также «сенсорными зонами», которые не являются строго ограниченными участками и могут перекрывать друг друга. Результаты современных исследований позволили ученым говорить о наличии проекционных (первичных и вторичных) и ассоциативных (третичных) зон коры. Возбуждение от соответствующих рецепторов в первичные зоны направляется по быстропроводящим специфическим путям, тогда как активация вторичных и третичных зон происходит по полисинаптическим неспецифическим путям. Кроме этого, корковые зоны связаны между собой многочисленными ассоциативными волокнами. По толщине коры нейроны распределяются неравномерно и, как правило, образуют шесть слоев. Основные афферентные пути в кору заканчиваются на нейронах верхних слоев, которые наиболее сильно развиты в центральных отделах зрительного, слухового и кожного анализаторов. Афферентные импульсы с участием звездчатых клеток коры передаются пирамидным нейронам, откуда уже обработанный сигнал уходит из коры к другим структурам мозга.
В коре входные и выходные элементы вместе со звездчатыми клетками образуют так называемые колонки –
12
функциональные единицы коры, организованные в вертикальном направлении. Соседние колонки имеют взаимосвязи, организующие участие множества колонок в осуществлении той или иной реакции. Возбуждение одной из колонок приводит к торможению соседних.
ФУНКЦИИ СЕНСОРНЫХ СИСТЕМ
Процесс передачи сенсорных сигналов сопровождается многократным их преобразованием и перекодированием и завершается высшим анализом и синтезом (опознанием образа), после чего и формируется ответная реакция организма. Не случайно
основными функциями сенсорной системы выделяются следующие:
обнаружение сигнала (начинается в рецепторе);
различение сигнала начинается в рецепторах. Однако в этом процессе участвуют нейроны всей сенсорной системы. Характеризует то минимальное различение стимула, которое сенсорная система может заметить);
передача и преобразование сигнала (в результате до высших центров мозга доводится наиболее существенная информация о раздражителе в форме, наиболее удобной для его надежного и быстрого анализа);
кодирование сигнала (преобразование информации в условную систему – код);
детектирование сигнальных признаков (избирательное выделение сенсорным нейроном того или иного признака раздражителя, имеющего значение. Осуществляется нейронами-детекторами, избирательно реагирующими лишь на определенные параметры стимула);
опознание образов (конечная и наиболее сложная операция сенсорной системы. Заключается в отнесении образа к тому или иному классу объектов, с которыми ранее встречался организм, т.е. классификации образов. Синтезируя сигналы от нейронов-детекторов, высший отдел нервной системы формирует «образ» раздражителя и сравнивает его с множеством образов, хранящихся в памяти.
Обнаружение и первичное различение сигналов обеспечивается рецепторами, а детектирование и опознание сигналов - нейронами коры больших полушарий.

13
ОБЩИЕ СЕНСОРНЫЕ МЕХАНИЗМЫ
А. Классификации рецепторов.
1. Источники стимулов (местоположение рецепторов): Функция рецепторов заключается в трансформации энергии
стимула в электрическую, а также в передаче стимулами в ЦНС информации об окружающем мире.
Все рецепторы классифицируются по следующим параметрам.
а) экстерорецепторы (ухо, глаз, рецепторы обоняния) получают импульсы, исходящие вне организма;
б) интерорецепторы (хемо- и барорецепторы) получают стимулы, исходящие из разных отделов организма.
2. Типы энергии стимулов:
а) механорецепторы определяют звуки, а также повреждение кожи;
б) терморецепторы определяют окружающую температуру;
Кора б. п/ш |
Кора б. п/ш |
изменение
приоритета
сенсорных
потоков
Таламус 
Кожа |
|
Слух |
|
Зрение |
|
Вкус |
|
Кожа |
|
Слух |
|
Зрение |
|
Вкус |
|
|
Как себя чувствует правая |
|
|
Студент слушает лекцию… |
||||
|
пятка? |
|
||
|
|
|
||
|
23 |
|||
Рис. 5. Роль таламуса в переработке сенсорной информации.
В целом деятельность таламуса можно сравнить с воронкой, пропускающей в кору только небольшую часть информации, причем положение «отверстия» воронки может регулировать сама кора.
в) фоторецепторы определяют свет; г) хеморецепторы определяют вещества, обладающие
запахом и вкусом;
14
3. Типы чувствительности.
Рецепторы подразделяются на рецепторы обонятельные, звуковые, вкусовые, болевые, холодовые (в том числе и тепловые - рецепторы температурной чувствительности) и тактильные.
4. Скорость адаптации:
а) медленно адаптирующие (тонические или статические) рецепторы генерируют потенциалы действия постоянно на всем протяжении стимуляции;
б) быстро адаптирующие (фазные или динамические) рецепторы уменьшают количество генерируемых потенциалов действия на протяжении стимуляции.
5. Типы нейронов:
а) Классификация.
Нейроны классифицируются по размерам и по скорости проведения импульсов:
1) Размеры:
а) по размерам нейроны делятся на 4 группы: 1 группа - 12-20 мкм в диаметре;
2группа- 6-12 мкм в диаметре;
3группа - 1-6мкм в диаметре;
4группа – менее 1 мкм в диаметре.
Волокна, отходящие от нейронов всех групп, кроме 4-й, являются миелинизированными.
2) Скорость распространения нервного импульса. Составной потенциал действия - суммированная активность
всех нервных волокон, составляющих нервный пучок. Потенциал действия генерируется на одном конце нервного пучка и распространяется вдоль его на другой конец, где расположены электроды:
а) по волокнам с наивысшей скоростью проведения импульсы потенциала действия первыми попадают к электродам - это первые волны составного потенциала действия. Последующие пики свидетельствуют о попадании импульсов по волокнам с меньшей скоростью проведения на регистрирующие электроды;
б) волокна, ответственные за первый пик, называются А-альфа волокна; ответственные за второй пик - А-гамма волокна; волокна, вызывающие последний пик - А-дельта волокна. Все перечисленные волокна -миелинизированые. Волокна с наименьшей скоростью проведения называются С-волокнами; все они -

15
немиелинизированые.
3) Функциональное распределение.
Некоторые важные двигательные и чувствительные волокна идентифицируются определенными типами нейронов:
1)афферентные нейроны, берущие начало в мышцах, принадлежат к 1 и 2 группам волокон;
2)миелинизированые афферентные нейроны болевой и температурной чувствительности принадлежат к А-дельта волокнам; в случае немиелинизированых волокон, они принадлежат
ктипу' С-волокон;
3)альфа-мотонейроны являются частью А-альфа волокон;
4)гамма-мотонейроны являются частью А-гамма волокон. По способу передачи сигнала в ЦНС рецепторы классифицируются на первичночувствующие и вторичночувствующие. Первичночувствующие разделяются на свободные и инкапсулированные
|
|
|
|
|
первичночувствующие |
|
вторичночувствующие |
||
|
||||
|
|
|
|
|
Рис. 6. Классификация рецепторов пов ЦНСспособу передачи сигнала в ЦНС. Первичные рецепторы – кожные, обонятельные, суставные, вторичные рецепторы – все остальные.
Первичные и вторичные рецепторы: 1 – тело чувствительного нейрона; 2
– периферический отросток чувствительного нейрона (дендрит); 3 – центральный отросток чувствительного нейрона (аксон); 4 – соединительнотканная капсула; 5 – рецептор; 6 – синапс между рецептором и чувствительным нейроном.
Б. Преобразование в рецепторах энергии.
Каждый вид рецепторов настроен на рецепцию определенного вида стимула, которые называются адекватными (соответствующими, подходящими). Каждый рецептор очень тонко
16
реагирует на свой адекватный стимул. Например, сетчатка может определить присутствие 1 фотона света. Однако при определенной силе другого стимула рецептор может отреагировать и на него. Например, такой механический стимул, как удар в глаз, может вызвать чувство света. Место преобразования в данном рецепторе находится на конце нервной терминали. В противоположность тому, место преобразования в зрительном, вкусовом и слуховом рецепторах находится в рецепторной клетке, отдаленной от афферентного волокна, а также в двух клетках, образующих синаптическую трансмиссию:
а) в месте преобразования энергия стимула трансформируется в электрический сигнал, называемый рецепторным (генераторным) потенциалом;
б) у каждого рецептора есть свой механизм образования рецепторного потенциала. В механическом рецепторе стимул генерирует рецепторный потенциал, деформируя нервную терминаль.
а) деформация открывает проницаемые для Na и К каналы и вызывает деполяризацию мембраны;
2. В области генерации разрядов рецептора рецепторный потенциал преобразуется в серию потенциалов действия:
б) рецепторный потенциал пассивно распространяется с области преобразователя в область генерации разрядов. При пороговой деполяризации области генерации разрядов возникает потенциал действия;
в) в конце потенциала действия рецепторный потенциал вновь вызывает пороговую деполяризацию в области генераций разрядов.
При достижении пороговой деполяризации возникает другой потенциал действия.
В. Адаптация.
Адаптация предназначена для снижения количества поступающей в мозг информации. Однако ответственные за сознание и внимание, стволовые механизмы мозга способны организовать поступающую в мозг информацию в должных пределах, так что для этого не требуется сенсорная адаптация. Основной задачей чувствительной адаптации является своевременное шифрование рецептором поступающей информации.
17
Существуют 2 основных механизма адаптации:
а) при первом механизме, несмотря на постоянное воздействие стимула, преобразовательный механизм не в состоянии генерировать постоянные рецепторные потенциалы;
б) при другом механизме, при наличии рецепторного потенциала генератор разрядов не в состоянии поддерживать серию потенциалов действия. Это происходит по причине снижения возбудимости мембраны генератора разрядов, возможными механизмами развития которой могут быть:
1)увеличение мембранной проводимости для К;
2)усиление инактивации электрического Na - К - насоса;
3)усиление инактивации Nа - каналов.
1. При увеличении частоты нанесения стимула увеличивается скорость генерирования разрядов в фазном рецепторе:
а) когда стимул наносится слишком медленно, адаптация генератора разрядов развивается до образования потенциала действия;
б) при быстром нанесении стимула возбуждение опережает адаптацию, и возникают стойкие потенциалы действия.
Г. Кодирование чувствительной информации.
В сенсорную систему заносится информация о местоположении интенсивности и качестве задействуемого на рецептор стимула.
1. Интенсивность кодируется (определяется) частотой нанесения стимула на чувствительный нейрон:
а) при увеличении интенсивности нанесения стимула увеличивается величина рецепторного потенциала (РП). Данная зависимость выражается законом Стивена:
Р П = К x J n ,
где РП - величина рецепторного потенциала; К и п - постоянные величины;
J - интенсивность нанесения стимула.
1) если предположить, что для определения стимула сенсорной системе необходим рецепторный потенциал, равный, по меньшей мере, 1 мВ, то для определения максимального стимула в сенсорной системе образуется таковой, в 50 раз больший минимального стимула; согласно закону Стивена, относительно

18
низкие значения рецепторного потенциала 1-50 мВ позволяют кодировать информацию, поступающую вместе со стимулами интенсивностью от 1 до 10 мин;
Интенсивность поступающего стимула кодируется частотой разрядов в чувствительном нерве.
2) Местоположение стимула кодируется, в основном, расположением чувствительных связей в коре мозга. Подобный
|
|
Потенциал |
|
|
|
А |
||
|
|
действия |
|
|
|
|
||
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рецепторный |
|
|
|
|
|
|||
потенциал |
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
Стимул |
|
|
|
|
||
|
|
|
|
|
|
|
Время |
|
|
|
|
|
|
|
|
||
|
Потенциал |
|
|
В |
|
|
||
|
|
действия |
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
Рецепторный |
|
|
|
|
|
|||
|
|
|
|
Адаптация тонических ( А ), |
||||
потенциал |
|
|
|
|
промежуточных ( Б ) и фазических |
|||
|
|
|
|
|
|
|
|
рецепторов ( В ) к длительно |
|
|
|
|
|||||
|
|
|
|
|
|
|
|
действующему раздражителю |
|
|
Стимул |
|
|
|
|
|
постоянной силы. |
|
|
|
|
|
|
|
|
|
Время
Кодирование раздражителя на уровне рецептора
Рис. 7. Классификация рецепторов по способности к
адаптации ТоническиеСила раздражителя(А) и фазическиекодируется(В) частотойрецепторыПД,
распространяющихся по сенсорным волокнам
Рис. 8. Кодирование раздражителя на уровне рецептора.
Минимальная интенсивность адекватного раздражителя, которая
Сила раздражителя кодируется частотой ПД, распространяющихся
приводит к реакции рецептора и генерации потенциала действия,
по сенсорнымазываетсяволокнамабсолютным. Минимальнаяпорогом чувствинтенсивностьтель ости рецептораадекватного. раздражителя, которая приводит к реакции рецептора и генерации потенциала действия, называется абсолютным порогом чувствительности рецептора.
19
механизм кодирования, названный топографическим представительством, используется зрительной и соматосенсорной системами для локализации источника поступления стимула. Слуховая сенсорная система использует с этой целью несколько механизмов:
а) каждый чувствительный нейрон получает информацию с определенной области, называемой рецепторным полем. Чем меньше величина рецепторного поля, тем точнее будет кодироваться информация о локализации источника стимуляции:
1)например, рецептивные поля fornea самые маленькие в зрительном анализаторе;
2)рецептивные поля, расположенные на подушечках пальцев, намного меньше таковых, расположенных на ладонях и на спине, с помощью которых тяжело точно локализовать точку нанесения механического стимула;
б) Локализация нанесения стимула может быть уточнена с помощью латерального торможения.
1)на кожу нанесены 2 стимула, и задачей рецепторов является определить наличие каждого из них. Наименьшее расстояние нанесения двух стимулов, на котором они будут восприниматься как различные, называется двухточечным порогом;
2)без латерального торможения два стимула определяются, как раздельные только в случае их нанесения на рецептивные поля, разделенные одним не стимулированным рецептивным полем: то есть 2 стимула вызывают одинаковые разряды во вех трех чувствительных нейронах;
3)латеральное торможение может снижать двухточечный порог, уменьшая количество генерируемых разрядов в иннервирующем центральное рецептивное поле нейроне: таким образом, ЦНС узнает о наличии 2 стимулов.
3. Качество стимулов кодируется при помощи большого количество механизмов:
а) самый простой механизм использует labeled line, в котором стимулы кодируются определенными стимулированными нервными проводящими путями:
1)таким образом, кодируются основные чувствительные
данные.
Так, чувство надавливания возникает при механическом
20
либо электрическом воздействиях на рецепторы кожи. Аналогичным образом, свет возникает вне зависимости от вида стимуляции сетчатки; звук всегда возникает при стимуляции улитки;
2) вид чувствительности зависит от места нанесения стимула на всем протяжении проводящего чувствительного пути. Например, стимуляция зрительной коры вызывает чувство света, а стимуляция обонятельной коры вызывает чувство запаха.
б) более сложный механизм кодирования используется при следующем типе активности в проводящем пути, несущем информацию к мозгу;
1) при временном способе кодирования один нейрон моет нети два типа сенсорной информации, зависящей от вида его активности.
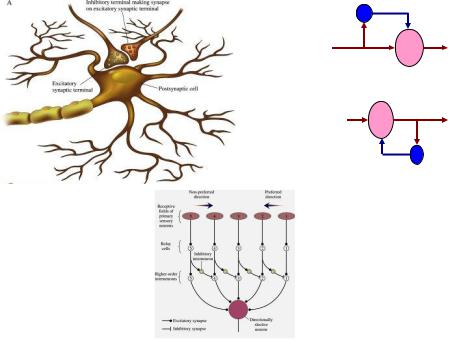
параллельное
возвратное
латеральное
Рис. 9. Типы торможения в нейронных цепях
Например, кожные температурные рецепторы определяют температуру ниже 30°С посредством генерирования "пачечных" разрядов, а температуру выше 30°С посредством генерирования потенциалов действия без "пачечных” разрядов;
2) при пространственном способе кодирования для вызова какого-либо чувства необходима энергия нескольких нейронов. Например, для кодирования разных вкусовых чувств могут потребоваться 3 нейрона. Вкус кислого может вызываться при