

Ответы на экзаменационные вопросы по гистологии
№1
Определение и задачи гистологии
Гистология (от греч.Histos – ткань, logos – учеяние) – наука о строении, развитии и жизнедеятельности тканей животных организмов. Гистология вместе с другими фундаментальными медико-биологическими науками изучает закономерности структурной организации живой материи, является одним из подразделений науки о жизни – биологии. Гистологию характеризуют 3 признака : 1- материя, которую она изучает, 2- методы, которые она использует, 3-законы,которые она познает. Материалом для гистологии являются ткани животных и человека, клетки, из которых состоят ткани и органы, сформированные из тканей. Гистология исследует филогенез и онтогенез тканей. По своему фундаментальному содержанию гистология рассматривается как фундаментальная наука.
Актуальными задачами гистологии являются: - разработка общей теории гистологии, отражающей эволюционную динамику тканей и закономерности эмбрионального и постнатального гистогенеза; - изучение гистогенеза как комплекса координированных во времени и пространстве процессов пролиферации, дифференциации, детерминации, интеграции, адаптивной изменчивости, программированной гибели клеток и др.; - выяснение механизмов гомеостаза и тканевой регуляции (нервной, эндокринной, иммунной), а также возрастной динамики тканей; - изучение закономерностей реактивности и адаптивной изменчивости клеток и тканей при действии неблагоприятных экологических факторов и в экстремальных условиях функционирования и развития, а также при трансплантации; - разработка проблемы регенерации тканей после повреждающих воздействий и методов тканевой заместительной терапии; - раскрытие механизмов молекулярно-генетической регуляции клеточной дифференцировки, наследования генетического дефекта развития систем человека, разработка методов генной терапии и трансплантации стволовых эмбриональных клеток; - выяснение процессов эмбрионального развития человека, критических периодов развития, воспроизводства и причин бесплодия.
№2
Основные периоды исторического развития гистологии.
1 период:
Накопление первоначальных фактов (1595-1800г), изобретение Янсоном первого микроскопа. Микроскоп увеличивал не более чем в 20 раз, вследствие чего не использовался по назначению. Для исследований использовались одиночные линзы, увеличивающие до 200 раз. Антуан ванн Левенгук изобрел линзы, увеличивающие в 300 раз. Именно он стал основоположником научной микроскопии. Левенгук в капле воды обнаружил много живых существ. Гук, будучи физиком, изготовил более совершенный микроскоп и впервые ввел понятие «Клетка» в 1665 году. Грю, будучи ботаником, изучал части растений, кору, листья, цветы. Систему однородных элементов он назвал тканью в 1682 году.
2 период:
В 1800г Франсуа Ксавье, не используя микроскоп, методом мацерации (настаивания), выделил 21 вид тканей и заложил основы гистологии под названием «Микроскопическая анатомия». Майер (1818) и Хойзингер (1821) переименовывают ее в гистологию. В 1838-1839 Шванн формирует клеточную теорию.
3 период:
Период борьбы за господство клеточной теории. Под ее влиянием бурно развивается гистология, и на основе ее цитология, гематология, эндокринология, иммунология, патологическая анатомия. Клеточная теория оказала значительное воздействие на общественную жизнь людей того времени.
4 период:
В 1945г. Портер и соавторы, исследуя под электронным микроскопом фибробласты, открывают ЭПС и хоронят идею о цитоплазме, как о бесструктурной системе. Новые знания о строении клеток были дополнены информацией об их функции.
№3
Гистология как учебная дисциплина, ее содержание.
Как учебная дисциплина гистология включает несколько разделов: 1) цитологию — учение о клетке; 2) эмбриологию — науку о развитии зародыша, закономерностях закладки и образования тканей и органов; 3) общую гистологию — учение о развитии, структуре и функциях тканей; 4) частную гистологию, изучающую микроскопическое строение органов и систем органов.
Цитоло́гия (греч. κύτος — «вместилище», здесь: «клетка» и λόγος — «учение», «наука») — раздел биологии, изучающий живые клетки, их органоиды, их строение, функционирование, процессы клеточного размножения, старения и смерти.
Эмбриология (от древнегреческого ἔμβρυον, зародыш, «эмбрион»; и -λογία, -логия) — это наука, изучающая развитие зародыша. Зародышемназывают любой организм на ранних стадиях развития до рождения или вылупления, или, в случае растений, до момента прорастания. Многими учёными, в том числе отечественными, эмбриология определяется более широко, как синоним биологии развития .
Гистология (от греч. histos — ткань, logos — учение) — наука о строении, развитии и жизнедеятельности тканей животных организмов.
Частная гистология служит основой для изучения микроскопического строения морфофункциональных единиц органов и органов в целом.
№4
Клеточная теория – теоретическая фундаментальная основа гистологии.
Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа строения и развития мира растений, животных и остальных живых организмов с клеточным строением, в котором клетка рассматривается в качестве общего структурного элемента живых организмов.
Клеточная теория — основополагающая для биологии теория, сформулированная в середине XIX века, предоставившая базу для понимания закономерностей живого мира и для развития эволюционного учения. Маттиас Шлейден и Теодор Шванн сформулировали клеточную теорию, основываясь на множестве исследований о клетке (1838). Рудольф Вирхов позднее (1858) дополнил её важнейшим положением (всякая клетка происходит от другой клетки).
Шлейден и Шванн, обобщив имеющиеся знания о клетке, доказали, что клетка является основной единицей любого организма. Клетки животных, растений и бактерии имеют схожее строение. Позднее эти заключения стали основой для доказательства единства организмов. Т. Шванн и М. Шлейден ввели в науку основополагающее представление о клетке: вне клеток нет жизни. Клеточная теория дополнялась и редактировалась с каждым разом.
Основные положения клеточной теории:
1) Клетка – единица строения, жизнедеятельности, роста и развития живых организмов, вне клетки жизни нет.
2) Ядро – главная составная часть клетки эукариот.
3) Новые клетки образуются только в результате деления исходных клеток.
4) Клетка – единая система, состоящая из множества закономерно связанных друг с другом элементов, представляющих собой определенное целостное образование.
5) Клетки многоклеточных организмов образуют ткани, ткани образуют органы. Жизнь организма в целом обусловлена взаимодействием составляющих клеток.
№5
Симпласт и синцитий как формы организации протоплазмы.
Симпласт – крупное образование, состоящее из цитоплазмы с множеством ядер. Примерами симпластов могут быть мышечные волокна позвоночных, наружный слой трофобласта плаценты. Симпласты возникают вторично за счет слияния отдельных клеток, или же в результате деления одних ядер без разделения цитоплазмы , без цитотомии.
Синцитий (соклетия) – первичная надклеточная форма организации жизни, представляющая собой протоплазматическую решетку, в узлах которой лежат ядра. У человека синцитиально связанные между собой клетки сохранились в семеннике, где эти связи синхронизируют развитие сперматоцитов.
№6
Характеристика межклеточного вещества.
Межклеточное вещество – продукт жизнедеятельности определенных групп клеток.Составная часть соединительной ткани позвоночных и многих беспозвоночных животных, включающая соединительнотканные волокна и аморфное основное вещество, выполняющая механическую, опорную, защитную и трофическую функции.
Межклеточное вещество образуется у зародыша из белков, углеводов, липидов, продуцируемых клетками эмбриональной соединительной ткани, начиная со стадии гаструлы. Гистогенез межклеточного вещества продолжается и в постэмбриональном периоде. Наибольшая роль в образовании межклеточного вещества принадлежит фибробластам, хондробластам, остеобластам. Полагают, что в образовании межклеточного вещества волокнистой соединительной ткани могут участвовать гистиоциты, лаброциты (тучные клетки) и другие.
Соединительнотканные волокна межклеточного вещества могут быть представлены коллагеновыми, эластическими, ретикулярными, или ретикулиновыми (аргирофильными), и другими волокнами, от чего зависит прочность, эластичность и в определенной степени архитектоника соединительной ткани органов (дерма различных участков кожи, сухожилия, строма кроветворных органов и так далее).
Аморфное основное вещество, окружающее соединительнотканные волокна и клетки соединительной ткани, состоит из высокополимерных соединений, от концентрации и состава которых в различных видах соединительной ткани зависят физические, химические и биологические свойства межклеточного вещества (вязкость, гидрофильность, интенсивность метаболических процессов, тургор и другие).
Состав волокон и аморфного вещества неодинаков в различных видах соединительной ткани, в различных ее топографических участках межклеточное вещество может быть минерализованным. При этом кристаллы минералов (фосфорнокислый кальций, углекислый кальций и другие) импрегнируют органическую основу межклеточного вещества твердых скелетных тканей (дентин, кость). С возрастом межклеточное вещество претерпевает инволюционные изменения: меняется соотношение основного вещества и волокон — масса волокнистых структур коллагена и плотность его «упаковки» возрастают, а масса основного вещества уменьшается, происходят конденсация эластических волокон, глубокие физико-химические изменения межклеточного вещества.
В эксперименте на животных выявлено, что недостаточное питание задерживает развитие возрастных изменений коллагена, а «атерогенная» диета вызывает его постарение.
Характером строения межклеточного вещества в значительной мере определяются функциональными особенности тех или иных видов соединительной ткани. Чем плотнее межклеточное вещество, тем сильнее выражена механическая, опорная функция, которая достигает наибольшего развития в костной ткани. Трофическая функция, напротив, лучше обеспечивается полужидким по консистенции межклеточным веществом (интерстициальная соединительная ткань, окружающая кровеносные сосуды).
Коллагеновые и эластические волокна, входящие в состав межклеточного вещества, построены из склеропротеинов — коллагена и эластина. Из коллагена состоят и ретикулиновые волокна отличающиеся повышенным содержанием углеводов и наличием липидов. В эластических волокнах имеется микрофибриллярный компонент, отличный от эластина по аминокислотному составу. Этот же компонент образует особую разновидность немногочисленных, сходных с эластическими волокнами межклеточного веществ (окситалановых), волокон резистентных к действию эластазы. Свойства основного вещества определяются преимущественно углеводно-белковыми биополимерами — гликозаминогликанами и гликопротеидами. Наличие гликозаминогликанов придает основному веществу межклеточного вещества выраженную базофильность. Качественные и количественные соотношения этих биополимеров, отличающихся интенсивным метаболизмом, различны в разных видах соединительной ткани.
№7
Клетка – главная форма организации протоплазмы.
Клетка – ограниченная активной мембраной , упорядоченная система биополимеров, образующих ядро и цитоплазму, участвующих в единой совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом. В своем историческом развитии протоплазма приобретает разнообразные формы, среди которых различают первичные и вторичные. Первичной формой протоплазмы является эукариотическая клетка многоклеточных. Это главная, исторически сложившаяся форма организации живой материи, обладающая всеми основными свойствами жизни, имеющая ядро, цитоплазму и цитоплазматические органеллы. Вторичные формы – весь многоклеточный мир.
№ 8
Величина и форма клеток. Факторы их обуславливающие.
Величина клетки определяется ядерно-цитоплазматическими отношениями и отношением площади поверхности к объему цитоплазмы, которые должны быть постоянными. Контактное торможение, которое определяет положение и пространство, занимаемое клеткой. Смещение константы ведет либо к делению клетки, либо к ее гибели. Форма клетки: Веретеновидная форма – клетка имеет утолщенную среднюю часть, брюшко, где лежит ядро и основные органеллы и 2 конца, фиксирующие к субстрату. (миоциты внутренних полых органов). Клетка призматической формы находится на поверхности органов , несут защитную функцию, приближены друг к другу (эпителиальные клетки). Клетки шаровидной формы – расположены не плотно и находятся в жидкости (лейкоциты). Клетки звездчатой формы (нейроны).
Между формой и содержанием, структурой и функцией имеется диалектическое взаимодействие. Основными структурными компонентами клетки являются:
1) клеточная поверхность (надмембранный комплекс, плазматическая мембрана, подмембранный комплекс);
2) цитоплазма (гиалоплазма, органеллы и включения);
3) ядро (кариолемма, ядрышко, хроматин, кариолимфа).
№9
Классификация цитоплазматических органелл.
|
2.1.2. Классификация органелл цитоплазмы |
|
А. Мембранные органеллы 1. За исключением митохондрий, все мембранные органеллы цитоплазмы объединяются в т. н. вакуолярную систему. В последнюю входят следующие органеллы: а) эндоплазматическая сеть (ЭПС), или эндоплазматический ретикулум (1 на рис.3.3) — совокупность плоских мембранных мешков (цистерн), вакуолей и трубочек; б) комплекс (аппарат) Гольджи (2), или пластинчатый комплекс, — несколько скоплений плоских мембранных цистерн, от которых отшнуровываются мелкие пузырьки; в) эндосомы и лизосомы (3) — мембранные пузырьки, содержащие ферменты гидролиза биополимеров (протеазы, нуклеазы, гликозидазы, липазы и т.д.); г) пероксисомы — мембранные пузырьки, содержащие ферменты, которые нейтрализуют пероксид водорода, а также оксидазы — ферменты окисления субстратов непосредственно кислородом. 2. Особняком от вышеназванных структур стоят митохондрии (4) — органеллы, отграниченные (как и ядро) двумямембранами, из которых внутренняя образует многочисленные впячивания (кристы) внутрь митохондрии. Б. Немембранные органеллы Их тоже можно подразделить на две неравные группы. 1. К первой группе относятся гранулярные органеллы; это рибосомы (5) — многочисленные небольшие частицы, состоящие из двух субъединиц рибонуклеопротеидной природы. 2. Вторая группа немембранных органелл — фибриллярные органеллы: I. сократительные структуры — миофибриллы и миофиламенты, II. а также элементы цитоскелета: а) микрофиламенты (МФ) — нити из белка актина (d = 5-7 нм), пронизывающие гиалоплазму в тангенциальномнаправлении (вдоль длинной оси клетки); б) промежуточные филаменты — более толстые нити (d = 10 нм), белковый состав которых различен в клетках разных тканей; в) микротрубочки (МТ) — полые трубки (d = 24 нм) из белка тубулина, имеющие в основном радиальную ориентацию в клетке. 3. Имеются также структуры, производные органелл второй группы. а) Производным МФ является каркас микроворсинок (п. 2.4.1). б) У МТ — производных больше. I. Во-первых, это центриоли (6 на рис. 3.3), каждая из которых представляет собой полый цилиндр, образованный микротрубочками. В клетке обычно содержится пара центриолей (диплосома), причем она входит в состав клеточного центра — органоида общего значения. II. Другое производное МТ — аксонема, служащая каркасом (осевой структурой) ресничек и жгутиков. |
№10
Клеточная поверхность и ее функции
Клеточная поверхность выполняет следующие функции: разграничительная, барьерно-защитная, рецепторная, транспортная, контактная, опорно-механическая, двигательная. Ее основными химическими компонентами являются: липиды (40%), белки (50%) и углеводы (10%). Соотношение этих веществ может варьировать в зависимости от функциональной активности клетки.
Надмембранный комплекс – это гликокаликс клетки, образован молекулами олигосахаридов, связанных с интегральными и покровными белками плазмолеммы. В состав гликокаликса входят: мальтоза, глюкоза, галактоза. Они образуют над плазмолеммой гетерополиморфные ветвящиеся цепочки. Гликокаликс осуществляет сортировку макромолекул из межклеточной среды, их удержание.
Плазматическая мембрана – образована слоем гидрофильных и гидрофобных липидов. Между липидами вставлены молекулы интегральных и полуинтегральных белков. Снаружи липидного комплекса лежат покровные белки. Плазматическая мембрана имеет каналы для натрия, калия, кальция, хлора и рецепторы к медиаторам, гормонам и цитокинам.
Подмембранный комплекс – слой глобулярных белков цитозоля и фиксированных к плазмолемме концов микротрубочек и филаментов клеточного скелета.
Производные клеточной поверхности: микроворсинки, реснички и жгутики.
Транспорт:
1)Простая диффузия – перемещение веществ по градиенту концентрации.
2)Облегченная диффузия - пассивный транспорт воды из клетки в клетку через специальные каналы.
3)Активный транспорт – перенос веществ с затратой энергии.
4)Экзо- и эндоцитоз.
№11
Основные функции клетки.
1)Деление клетки –вид размножения клеток. Наиболее распространенным способом деления соматической клетки является митоз. Во время митоза клетка проходит ряд последовательных стадий, в результате которых каждая дочерняя клетка получает такой же набор хромосом, как и у материнской клетки.
2)Обмен веществ. Основная функция клетки. Из межклеточного вещества в клетку постоянно поступают питательные вещества и кислород, и выделяются продукты распада. Обмен веществ выполняет 2 функции: обеспечение клетки строительным материалом. Из веществ, поступающих в клетку, непрерывно происходит биосинтез белков, углеводов, липидов, из которых впоследствии формируются органеллы. Вторая функция – это обеспечение клетки энергией. (Энергетический и пластический обмен).
3)Раздражимость – реакция на физические и химические воздействия окружающей среды. В возбужденном состоянии разные клетки выполняют свойственные им функции.
№12
Синтетический аппарат клетки.
Синтетический аппарат клетки обеспечивает синтезы различных веществ и включает ЭПС, кГ и рибосомы.
Рибосомы – округлые мелкие, немембранные органеллы, состоящие из двух округлых субъединиц – малой и большой. Каждая субъединица образованы рибосомальной РНК и сложным набором белков. Синтез рРНК и сборка субъединиц происходит в ядрышке, а их объединение – уже в цитоплазме. Рибосомы обеспечивают процесс трансляции белка. Малая субъединица связывается с иРНК, а большая катализирует образование пептидных связей между аминокислотами.
Одиночные рибосомы неактивны и для белкового синтеза они объединяются в цепочки, нанизываясь на молекулу иРНК. Так образуются полисомы. Свободные полисомы синтезируют белки, которые диффузно распределяются в гиалоплазме.
Синтез мембранных белков, лизосомальных белков и секреторных белков, которые будут выведены за пределы клетки, осуществляют полисомы, прикрепленные к ЭПС.
При синтезе секреторных и лизосомальныз белков используется особый механизм, который позволяет полипептидной цепи проникать внутрь полости ЭПС. После завершения синтеза молекула приобретает вторичную и третичную структуру и выйти наружу уже не может. Синтез таких белков начинается с особого участка – сигнального пептида. Благодаря ему рибосома соединяется с рибофорином – белком, который встроен в мембрану ЭПС. В присоединении участвует еще и специальный мембранный рецептор ЭПС. После присоединения рибосомы рибофорин приобретает форму канала, через который проходит синтезируемая полипептидная цепь. Когда белковая молекула готова, сигнальный участок отсоединяется.
Если синтезируется мембранный белок, то в полипептидной цепи оказывается еще один участок, который заякоривает белковую цепь в мембране.
ЭПС представляет собой сложную систему мембранных полостей. Обычно в форме плоских цистерн, распределенных по всей клетке.
Есть два типа ЭПС – гранулярная и агранулярная. К поверхности грЭПС прикрепляются полисомы.
Итак, главные функции грЭПС: синтез, химическая модификация, накопление и транспортировка белков.
Агранулярная ЭПС является продолжением грЭПС, но лишена я ЭПС является продолжением грЭПС, но лишена рибосом и имеет иной набор белков-ферментов. аЭПС – трубчатыми каналами. У нее множество функций:
Синтез липидов и холестерина, поэтому ее много в клетках, синтезирующих стероидные гормоны и жиры.
Синтез гликогена (клетки печени)
Детоксикация вредных веществ (лекарственные препараты, алкоголь, токсины)
Накопление Са2+, необходимого для сокращения мышечных клеток.
От ЭПС отшнуровываются транспортные пузырьки, содержащие синтезированные вещества, перемещаются в сторону комплекса Гольджи и сливаются с ним.
Комплекс Гольджи – мембранная органелла, представленная диктиосомами (стопка из 3-10 плоских цистерн). Диктиосома имеет незрелую поверхность, обращенную к ЭПС (цис-) и зрелую, обращенную к плазмолемме (транс-). С цис-поверхностью сливаются транспортные мембранные пузырьки, содержащие продукты синтеза, которые отшнуровываются от ЭПС. Вещества, попавшие в полости кГ, направляются в различные части диктиосомы, где подвергаются процессингу. Это химические превращения молекул – к ним могут присоединяются сахара, сульфатные и фосфатные группы, белковые молекулы могут частично расщепляться и т.д.
От боковых участков кГ отшнуровываются гидролазные пузырьки, заполненные гидролитическими ферментами. Из них формируются лизосомы.
Функции кГ:
Синтез полисахаридов и гликопротеинов (слизь, гликокаликс).
Процессинг молекул
Накопление продуктов синтеза, их упаковка и транспортировка.
Формирование лизосом.
,
№13
ГЭРЛ-система и поток мембран в клетке.
ГЭРЛ (Г — Гольджи, Р- эндоплазм.ретикулом, Л -лизосомы) — — система и поток мембран в клетке: синтетический и транспортный компармент клетки, связанный с потоком мембран клеток. Чем активней клетка, тем активней поток. Мембранный транспорт, или мембранный поток («membrane flow»), является необходимым для обмена веществ между различными клеточными органеллами. В настоящее время для описания этого процесса принята следующая модель:
1) Внутриклеточный транспорт является направленным; транспорт от эндоплазматического ретикулума через аппарат Гольджи к лизосомам, эндосомам или клеточной поверхности называется антероградным; транспорт в обратном направлении — ретроградным .
2) Транспортный путь проходит через ряд отдельных компартментов, перенос вещества между которыми осуществляется преимущественно с помощью окаймленных мембранных пузырьков . В клетке существует несколько типов окаймляющих белков и, соответственно, несколько типов окаймленных пузырьков, курсирующих между определенными компартментами.
3) Не все органеллы способны к прямому обмену пузырьками, например, транспорт между эндоплазматическим ретикулумом и эндосомами, лизосомами и плазматической мембраной может быть осуществлен только через аппарат Гольджи .
4) Процесс, в результате которого нужные вещества направляются в соответствующие компартменты, называют сортингом («sorting»).
№14
Митохондрии и их энергетические функции.
Митохондрии – органеллы синтеза АТФ. Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии для синтеза молекул АТФ.
Форма и размеры митохондрий митохондрий животных клеток разнообразны и их количество в клетках варьирует от единиц до сотен.
Основная функция митохондрий – синтез АТФ, происходящий в результате процессов окисления органических субстратов и формирование АДФ. Начальные этапы этих сложных процессов совершаются в гиалоплазме. Здесь происходит первичное окисление субстратов (например сахаров до ПВК с одновременным синтезом небольшого количества АТФ. Эти процессы совершаются при отсутствии кислорода. Все последующие выработки энергии (дыхание) – анаэробное окисление и синтез основной массы АТФ осуществляется с потребление кислорода и локализ. внутри митохондрий. При этом происходит дальнейшее окисление пирувата и других субстратов энергетического обмена с выделение углекислого газа и переносом протонов на их акцепторы. Эти реакции осуществляются с помощтю ряда ферментов ЦТК, которые локализированы в матриксе митохондрий.
В мембранах крист митохондрий располагаются системы дальнейшего переноса электронов и сопряженного с ним фосфорилирования АДФ. При этом происходит перенос электронов с одного белка-акцептора электронов к другому и,наконец, связывание их с кислородом, вследствие чего образуется вода. Одновременно с этим часть энергии, выделяемой при таком окислении в цепи переноса электронов запасается в виде макроэргической связи при фосфорилировании АДФ, что приводит к образованию большого количества молекул АТФ. Именно на мембранах крист митохондрий происходит процесс окислительного фосыорилирования с помощью здесь расположенных белков цепи окисления и ферментов фосфорилирования АДФ, АТФ-синтетазы.
№15
Пищеварительный аппарат клетки – лизосомы.
Класс шаровидных структур, ограниченных одиночной мембраной. Характерным признаком является наличие в них гидролитических ферментов – гидролаз (протеинкиназы, липазы), расщепляющих различные биополимеры. Можно выделить 3 типа лизосом: первичные, вторичные и остаточные тельца.
Первичные лизосомы – мелкие мембранные пузырьки, заполненные бесструктурным веществом, содержащим гидролазы, в т.ч. и фосфатазу, которая является маркерным лизосомным ферментом. Местом синтеза фосфатазы является гранулярная ЭПС, затем этот фермент появляется в проксимальных участках диктиосом, а затем в мелких везикулах по периферии диктиосом и, наконец, в первичных лизосомах. Вторичные лизосомы – вторичные пищеварительные вакуоли, формируются при слиянии первичных лизосом с фагосомами (фагоцитарными вакуолями) или пиноцитозными вакуолями, образуя гетерофагосомы. Вещества, попавшие в состав вторичной лизосомы, расщепляются гидролазами до мономеров, которые транспортируются через мембрану лизосом в гиалоплазму где они включаются в обменные процссы. Расщепление, переваривание биогенных молекул внутри лизосом может идти в ряде клеток не до конца. В этом случае в полостях накапливаются непереваренные продукты. Такая лизосома называется остаточным тельцем.
№16
Регуляция синтеза белка в клетке
Биосинтез белка – многостадийный процесс синтеза полипептидной цепи из а/к остатков, происходящий на рибосомах клеток живых организмов с участием молекул мРНК и тРНК.
Биосинтез можно разделить на 3 стадии: транскрипция, процессинг и трансляция.
Во время транскрипции происходит считывание генетической информации, зашифрованной в молекулах ДНК, и запись этой информации в молекулы и РНК. В ходе ряда последовательных стадий процессинга из мРНК удаляются некоторые фрагменты, ненужные в последующих стадиях, и происходит редактирование нуклеотидных последовательностей. После транспортировки когда из ядра к рибосомам происходит собственно синтез белковых молекул путем присоединения отдельных а/к остатков к растущей полипептидной цепи.
Процессинг РНК: Между транскрипцией и трансляцией молекула мРНК претерпевает ряд изменений, которые обеспечивают созревание функционирование матрицы для синтеза полипептидной цепочки. К 5’ концу присоединяется кэп (модифицированный гуаниновый нуклеотид, который добавляется на 5’ (передний) конец незрелой матричной рибонуклеиновой кислоты), а к 3’ концу поли – А- хвост, который увеличивает длительность жизни иРНК. С появлением процессинга в эукариотической клетке стало возможно комбинирование экзонов гена для получения большего разнообразия белков, кодируемых единой последовательностью нуклеотидов ДНК.
Трансляция: заключается в синтезе полипептидной цепи в соответствии с информацией, закодированной в мРНК. А/к последовательность выстраивается при помощи тРНК, которые образуют комплексы с а/к. Каждой а/к соответствует своя тРНК, имеющая соответствующий антикодон, «подходящий» к кодону мРНК. Во время трансляции рибосома движется вдоль мРНК, по мере этого наращивается полипептидная цепь. Энергией биосинтез белка обеспечивается за счет АТФ. Готовая белковая молекула затем отщепляется от рибосомы и транспортируется в нужное место клетки. Для достижения своего активного состояния некоторые белки требуют дополнительной посттрансляционной модификации.
№17
Строение и функции ядра
Ядро – система генетической детерминации и регуляции белкового синтеза.
Функции ядра:Хранение и поддержание наследственной информации в виде неизменной структуры ДНК . В ядре происходит воспроизведение и редупликация молекул ДНК, что дает возможность при митозе двум дочерним клеткам получить совершенно одинаковые в качественном и количественном отношении объемы генетической информации. Вторая функция – создание аппарата белкового синтеза. Это не только синтез, транскрипция на молекулах ДНК разных иРНК, но и транскрипция всех видов тРНК и рРНК. В ядре происходит образование субъединиц рибосом.
Структура ядра:Ядрышко – имеет величину от 1-5 мкм, очень богаты РНК, базофильны, не является самостоятельной структурой или орагнеллой. Ядрышко является местом образования рРНК и рибосом, на которых происходит синтез полипептидных цепей в цитоплазме. Образование ядрышек и их число связаны с активностью и числом определенных участков хромосом – ядрышковых организаторов, которые расположены большей частью в зонах вторичных перетяжек. Вокруг ядрышек есть зона конденсированного хроматина. Схема участия ядрышек в синтезе цитоплазматических белков: на ДНК ядрышкового организатора образуется предшественник рРНК, который в зоне ядрышка одевается белком, здесь происходит сборка рибонуклеопротеидных частиц – субъединиц рибосом; субъединицы, выходя из ядрышка, участвуют в синтезе белка. Ядерная оболочка – состоит из внешней ядерной мембраны и внутренней мембраны оболочки, разделенных перинуклеарным пространством или цистерной ядерной оболочки. Ядерная оболочка содержит ядерные поры. Внешняя мембрана ядерной оболочки непосредственно контактирующая с цитоплазмой клетки, имеет ряд структурных особенностей, позволяющих отнести ее к собственно мембранной системе ЭПС: на ней со стороны гиалоплазмы расположены многочисленные полирибосомы , а сама внешняя ядерная мембрана может прямо переходить в мембраны ЭПС. Внутренняя мембрана связана с хромосомным материалом ядра. Кариоплазма(от карио... и плазма), кариолимфа, ядерный сок, содержимое клеточного ядра, в к-рое погружены хроматин, ядрышки, а также различные внутриядерные гранулы. После экстракции хроматина химич. агентами в К. сохраняется т. н. внутриядерный матрикс, состоящий из белковых фибрилл толщиной 2-3 нм, к-рые образуют в ядре каркас, соединяющий ядрышки, хроматин, поровые комплексы ядерной оболочки и др. структуры.
№18
Способы репродукции протоплазмы
Можно выделить несколько источников и способов обновления живой материи. Главным является клеточный камбий, а в его составе стволовые клетки. Большинство стволовых клеток находится в клеточном цикле. Каждая стволовая клетка способна совершить от 50 до 65 циклов. Всякий раз из нее образуются 2 клетки. Одна из них вступает в фазу G0, получает программу развития, дифференцируется и функционирует в составе органа, другая сохраняя свойства стволовой клетки, превращается в GI.
Другим источником репродукции являются дифференцированные клетки органов, не имеющие стволовых. Здесь каждая делящаяся клетка дает 2, получающие обе программы развития. Образующиеся клетки называют дочерними. Например, все дифференцированные гепатоциты сохраняют способность к пролиферации, благодаря чему печень обладает хорошими регенеративными способностями.
Клетки, утратившие способность к делению, обновляют живую материю с помощью эндорепродукции, т.е их новая протоплазма образуется в прежней форме. Это двуядерные клетки, которые могут возникать с помощью абортивного митоза, так и митоза.
Соматическая протоплазма возникает наиболее часто при атипичных формах митоза – полицентрическом и моноцентрическом. П. митозы характерны для мегакариоцитов красного костного мозга. Единственным полноценным способом воспроизводства клеток является митоз.
№19
Жизненный цикл клетки
Делящиеся и неделящиеся клетки. Митоз. Дифференцировка и специализация клеток. Этапы жизненного цикла специализированной клетки. Некроз и апоптоз. Регуляция численности клеток в организме.
До сих пор много тайн клетки остаются неразгаданными. Загадочным во многом остается и запрограммированный генетически алгоритм ее жизни, названный жизненным циклом клетки (клеточным циклом). Жизненный цикл клетки (рисунок 1) начинается с момента ее образования после деления родительской клетки и заканчивается либо новым делением, либо превращением в специализированную клетку.

|
Рисунок 1. Жизненный цикл клетки: 1 - интерфаза; 2 - митоз; 3 - дифференцировка; 4 - функционирование специализированной клетки |
Большинство клеток продолжает делиться. Им свойственен клеточный цикл, состоящий из периодически повторяющихся стадий: так называемой интерфазы (1) – этапа подготовки к делению и непосредственно процесса деления – митоза (2). К этапам дифференцировки (3) и функционирования специализированной клетки (4) мы вернемся чуть позже.
На стадии подготовки к делению происходит удвоение генетического материала (редупликация ДНК). Масса клетки во время интерфазы увеличивается до тех пор, пока она примерно вдвое не превысит начальную. Отметим, что сам процесс деления намного короче этапа подготовки к нему: митоз занимает примерно 1/10 часть клеточного цикла.
Цикличность (периодическое повторение) стадий интерфазы и митоза можно проиллюстрировать на примере фибробластов – одного из видов клеток соединительной ткани (рисунок 2). Так, нормальные фибробласты эмбриона человека размножаются приблизительно 50 раз. Каков генетически запрограммированный предел возможных делений клетки – это одна из неразгаданных тайн биологии.
![]()
|
Рисунок 2. Цикличность стадий интерфазы и митоза: 1 - интерфаза, стадия подготовки к митозу; 2 - митоз (деление клетки) |
Жизненный цикл клеток базального слоя эпидермиса в обычных условиях составляет 28-60 дней. При повреждении кожи (конкретнее – при повреждении мембран и разрушении клеток эпидермиса под воздействием внешних факторов) выделяются особые биологически активные вещества. Они значительно ускоряют процессы деления (это явление называется регенерацией), именно поэтому ранки и ссадины так быстро заживают. Максимальной регенеративной способностью обладает эпителий роговицы: одновременно в стадии митоза находятся 5-6 тысяч клеток, продолжительность жизни каждой из которых 4-8 недель.
Хотя все клетки появляются путем деления предшествующей (материнской) клетки (“Всякая клетка от клетки”), не все они продолжают делиться. Клетки, достигшие некоторой стадии развития при дифференцировке, могут терять способность к делению.
Дифференцировка – возникновение различий в процессе развития первоначально одинаковых клеток, приводящее к их специализации. Процесс дифференцировки заключается в последовательном считывании и использовании наследственной информации, что обеспечивает синтез различных белков (в первую очередь ферментов), характерных для данного вида клеток. Другими словами, различия между клетками определяются набором белков, синтезируемых в клетках определенного вида.
Различия между клетками определяются набором синтезируемых в них белков.
При дифференцировке набор хромосом в клетке не меняется, изменяется лишь соотношение активных и неактивных генов, кодирующих различные белки.
Существуют два типа регуляции экспрессии (активации или блокирования) генов:
Кратковременная адаптивная активация (реже блокирование), зависящая, в частности, от концентрации вещества, включающегося в обмен веществ (исходного вещества или продукта метаболизма). Этот механизм выработался эволюционно как приспособительная реакция и особенно ярко проявляется у животных (например, быстрый синтез пигментов у хамелеона в зависимости от условий).
Длительное (в течение всей жизни клетки и/или многих генераций клеток!) блокирование или активация гена, возникающее в ходе клеточной дифференцировки. Например, в ДНК любой клетки желудка есть ген, отвечающий за синтез белков, из которых состоит ноготь. Но он необратимо блокирован гистонами и другими белками (этот участок ДНК плотно упакован), что никогда не позволит считывать с него информацию. Поэтому в желудке не растут ногти; а гены, ответственные за синтез гемоглобина, функционируют только у молодых форм эритроцитов, но не действуют в зрелых эритроцитах или других клетках.
На рисунке 1. цифрами 3 и 4 отмечены этапы дифференцировки и активного функционирования специализированной клетки.
Нервные клетки мозга, однажды возникнув, уже не делятся. В течение жизни число нейронов постепенно уменьшается. Поврежденные ткани мозга неспособны восстанавливаться путем регенерации. Однако изначально число нейронов в мозге настолько велико, что до конца жизни человека они способны поддерживать необходимые связи в нервной системе.
В качестве примера клеток, неспособных к делению, можно рассмотреть эритроциты. Как известно, эритроциты в процессе специализации теряют ядро, следовательно, не имеют в своем составе ДНК. Возникают эритроциты из так называемой стволовой клетки костного мозга. Клеткой-предшественницей(стволовой клеткой) называют клетки кроветворной ткани, которые на протяжении всей жизни человека сохраняют способность делиться и, тем самым, поставлять дочерние клетки, которые в дальнейшем будут специализироваться в одном направлении и замещать погибшие клетки. Срок жизни и активного функционирования эритроцитов невелик (около 4 месяцев), затем они разрушаются, в основном в селезенке.
Этапы жизни специализированной клетки, неспособной к делению (нейрона, эритроцита), условно можно изобразить на оси времени линией, разделенной на несколько отрезков ( рисунок 3 ). Эти отрезки дают представление о временном соотношении периодов жизни такой клетки: рождения, созревания и активного функционирования, угасания (старения) и естественной гибели.
![]()
|
Рисунок 3. Этапы жизненного цикла специализированной клетки: 1 - рождение в процессе деления материнской клетки; 2 - созревание и дифференцировка; 3 - активное функционирование; 4 - угасание (старение); 5 - запрограммированная клеточная гибель |
Время протекания каждого этапа и продолжительность жизненного цикла для однотипных клеток в нормальных условиях практически одинаковы.
Продолжительность жизненного цикла однотипных клеток в нормальных условиях практически одинакова.
Например, эритроциты живут 90-125 дней, а тромбоциты – всего 4 суток. Это говорит о том, что клетки используют для отсчета времени своей жизни некий механизм, алгоритм, заложенный в них природой. И в каждый момент жизни клетка строго следует законам, продиктованным этим алгоритмом.
На всех этапах клеточного цикла варьируют значения некоторых параметров жизнедеятельности клетки, и, в частности, отмечается различная скорость и интенсивность протекания процессов метаболизма( рисунок 4 ). Это обусловлено, в первую очередь, непрерывно меняющейся активностью ферментов, благодаря которым протекают все реакции в клетке. Ферменты могут синтезироваться в клетке “по мере надобности”, активироваться, временно блокироваться или полностью разрушаться (подробнее о ферментах будет сказано позже).
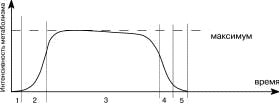
|
Рисунок 4. Интенсивность метаболизма на различных этапах жизни клетки: 1 - рождение; 2 - созревание и дифференцировка; 3 - активное функционирование; 4 - угасание (старение); 5 - запрограммированная клеточная гибель |
Рассмотрим подробнее наиболее характерные процессы, происходящие на каждом из этапов клеточного цикла.
Рождение. Отправным моментом жизни любой клетки (кроме половой, для которой характерен мейоз) считают деление материнской клетки с образованием двух идентичных дочерних – митоз (от греческогоmitos – нить). Во время митоза основная задача материнской клетки – поровну передать равноценный в количественном и качественном отношении генетический материал дочерним клеткам.
Митоз часто называют “танцем хромосом”. Каждая следующая фигура в этом танце не случайна, здесь нет ни одного лишнего или бессмысленного “па” – это еще один четкий, выверенный природой алгоритм. В. Дудинцев в романе “Белые одежды” так описывает процесс деления клетки: “Хромосомы шевелились, как клубок серых червей, потом вдруг выстроились в строгий вертикальный порядок. Вдруг удвоились – теперь это были пары. Тут же какая-то сила потащила эти пары врозь, хромосомы подчинились, обмякли, и что-то повлекло их к двум разным полюсам.”
Деление клетки на две идентичные (митоз) характеризуется сменой нескольких морфологически и физиологически различающихся стадий ( рисунок 5 ). На первой стадии митоза хроматин плотно упаковывается (этот процесс называется суперспирализацией хроматина) с образованием хромосом (1). Каждая хромосома состоит из двух идентичных половинок (хроматид) – будущих дочерних хромосом. Затем при сокращении так называемого веретена деления (2), представляющего собой комплекс микротрубочек и микрофибрилл, дочерние хромосомы расходятся, буквально подтягиваются нитями веретена деления к противоположным полюсам клетки. После окончательного расхождения дочерние хромосомы вновь раскручиваются, превращаясь в длинные и тонкие нити хроматина (3). Веретено деления исчезает, хроматин в дочерних клетках окружается ядерной оболочкой, и между дочерними клетками образуется поперечная перетяжка (4) из клеточных мембран.
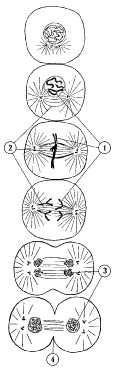
|
Рисунок 5. Последовательность стадий митоза (схема): 1 - хромосомы; 2 - веретено деления; 3 - хроматин; 4 - поперечная перетяжка |
Хромосомы, как мы уже говорили, представляют собой максимально плотно упакованные нити ДНК, с которых на этапе деления невозможно считывание информации. Соответственно, на этапе деления не происходит биосинтеза белка, интенсивность процессов метаболизма минимальна, транспорт веществ в клетку и из нее практически равен нулю. Все процессы в делящейся клетке направлены на выполнение главнейшей задачи – максимально точно, без искажения, передать генетическую информацию дочерним клеткам, – в ущерб второстепенным (на данном этапе!) функциям.
Созревание. В этот период происходит дифференцировка клеток и становление ключевых ферментных систем. Клетка готовится выполнять предназначенные природой функции, постепенно активизируя свой обмен веществ.
Активное функционирование. Интенсивность реакций метаболизма и сопряженного с нимэнергетического обмена в это время максимальны.
В период активного функционирования интенсивность обмена веществ в клетке максимальна.
Процессы в клетке направлены на обеспечение постоянства внутренней среды и выполнение специфических функций: нейрон воспринимает и передает нервный импульс, эритроцит переносит кислород и так далее.
Угасание (старение). Этот процесс запрограммирован генетически и, в первую очередь, проявляется уменьшением выработки и активности ферментов в клетке. При этом замедляются биохимические реакции, тормозится метаболизм и энергетический обмен.
Период старения клетки характеризуется уменьшением выработки и активности ферментов.
Стареющие клетки, как правило, имеют неудвоенное количество ДНК, но сохраняют жизнеспособность и некоторую метаболическую активность в течение определенного времени.
Естественная гибель клетки (апоптоз). К сожалению, до сих пор процесс естественной гибели клеток до конца не изучен.
Известно, что в клетке из-за блокирования ферментов прекращается синтез белка, а нет белка – нет и жизни. Морфологически апоптоз характеризуется разрушением ядра и цитоплазмы. “Осколки” погибшей клетки поглощаются и перерабатываются специальными клетками иммунной системы – фагоцитами. Но ведь клетки могут погибнуть и под воздействием случайных факторов (механических, химических и любых других). Случайная гибель клеток (а также ткани, органа) в биологии называется некрозом. Важно то, что естественная клеточная гибель (апоптоз) в отличие от некроза не вызывает воспаления в окружающих тканях.
Апоптоз не вызывает воспаления в окружающих тканях.
В организме запрограммированная клеточная гибель выполняет функцию, противоположную митозу, и, тем самым, регулирует общее число клеток в организме. Апоптоз играет важную роль в защите организма при вирусных инфекциях. В частности, иммунодефицит при ВИЧ-инфекции определяется нарушениями в контроле апоптоза.
Теперь, когда мы рассмотрели все этапы жизненного цикла клеток, коротко остановимся на процессах регуляции численности клеток в организме. Во время эмбриогенеза (первого этапа внутриутробного развития) число клеток постоянно возрастает, причем в геометрической прогрессии ( рисунок 6 ).
Зигота, образовавшаяся после слияния яйцеклетки и сперматозоида, делится с образованием двух дочерних клеток. Затем, в результате последовательных делений, образуются четыре, восемь, шестнадцать клеток и так далее. Параллельно с увеличением численности на этапе эмбриогенеза происходит дифференцировка клеток – так образуются ткани (смотри лекцию Ткани, их строение и функции).
Во взрослом организме общая численность клеток стабильна, она остается практически неизменной на протяжении многих лет ( рис. 7).
Это происходит за счет уравновешивания процессов возникновения новых клеток (митоза) и гибели клеток, естественной (апоптоза) или случайной (некроза). При смещении равновесия, например, гибели большого количества клеток в результате травмы или другого негативного воздействия, включаются механизмы регенерации (увеличение интенсивности деления клеток для замещения погибших), о которых уже было сказано. Таким образом, общая численность клеток поддерживается практически на постоянном уровне.
№ 20
Клеточный цикл и его фазы
Клеточный цикл состоит из митоза и интерфазы.
Митоз начинается с профазы. В профазе происходит спирализация хромосом, образование нитчатых структур. Хроматин конденсируется в хромосомы и полностью исчезает. Исчезает ядрышко. В конце профазы разрушается кариолемма. В цитоплазме распадается и исчезает ЭПС и к.Гольджи, прекращается образование РНК и синтез белка. Лизосомы и митохондрии сохраняются и начинает формироваться аппарат деления.
Прометафаза: движение хромосом к полюсам клетки.
Метафаза: Свободнолежащие хромосомы мигрируют к экватору клетки и образуют фигуру, которую называют экваториальной пластинкой. Кинетохоры хромосом у полюсов ядра связываются с микротрубочками. Стадию метафазы называют одиночной звездой – монастер или материнской звездой. Каждая хромосома состоит из сестринских гомологичных хроматид, полностью разделенных, исключая район кинетохора. Далее происходит разделение кинетохора.
Анафаза: Поделившиеся материнские хромосомы начинают мигрировать к полюсам клетки. По экватору клетки появляется борозда деления, знаменующая начало цитотомии.
Телофаза: Реконструкция ядра и образование новых клеток. После стабилизации хромосом на полюсах клетки начинается процесс реконструкции ядерной оболочки. Деконденсация хромосом, появляются глыбки хроматина, формируется ядрышко.
Интерфаза
G1 : Образование РНК, синтез белка, рост клетки. В этой фазе клетка задерживается в точке рестрикции и может находиться в ней долго. Для преодоления этой точки нужен специальный белок.
S: В эту фазу попадает диплоидная клетка, имеющая хромосомы. Синтез ДНК. Клетка становится тетраплоидной.
G2: Образование АТФ. Синтез белков-тубулинов для сборки микротрубочек и веретена деления.
№21
Хромосомы и их организация. Хромосомный набор человека.
Хромосомный набор или кариотип, в диплоидном количестве содержат соматические клетки и в гаплоидном – половые. В наборе различают автосомы, ответственные за все признаки организма, исключая принадлежность к полу. Хромосомный набор контролируют 3 закона: Закон постоянства числа хромосом: Диплоидный набор у человека состоит из 46 хромосом. Среди 46 – 23 наследуются по материнской и 23 по отцовской линии и являются гомологичными ( кроме х и у).
Закон постоянства формы хромосом: парные отцовские и материнские хромосомы имеют одинаковую форму. Различают метацентрические, субметацентрические, акроцентрические. Некоторые хромосомы имеют сателлит и поперечную исчерченность.
Закон постоянства величины хромосом: Хромосомы набора имеют разную величину. Самую большую величину имеют хромосомы первой пары, самую малую из аутосомного состава – 22 пара. Наименьшие размеры имеет у-хромосома.
Хромосомы образованы из ДНК, гистонов и негистоновых белков. Гистоны необходимы для укладки, компактизации нитей ДНК в хромосомы. Основу хромосомы составляет двойная спираль ДНК. Первый уровень ее укладки – нуклеосомный (отрезки ДНК связываются в глобулы гистонов). Нуклеомерный - формирование петель из молекул ДНК. Хромонемный – образование нитей. Хроматиды – нитчатые хромонемы составляющие структуру хроматиды.
№22
Паранекроз, дистрофия и смерть клетки. Апоптоз и некроз.
Паранекроз – совокупность неспецифических обратимых изменений цитоплазмы, возникающих под действием различных агентов. При паранекрозе в цитоплазме происходит разрушение белковых тел, ее обводнение и увеличение сорбционных свойств. Важным признаком паранекроза так же считают изменение кислотно-щелочного равновесия. Среда клетки меняется на кислую. Поврежденные клетки резко снижают митотическую активность, часто задерживаются на разных стадиях митоза, главным образом из-за нарушения митотического аппарата, очень чувствительного к изменениям внутренней среды. Повреждение клеток внешними и внутриорганизменными факторами может привести к нарушениям регуляции их метаболизма. При этом происходит интенсивное отложение или наоборот, резорбция ряда клеточных включений. Кроме того наблюдается нарушение регуляции проницаемости клеточных мембран, что приводит к вакуолизации мембранных органелл. Такие изменения называются дистрофией. При необратимых повреждениях клетки гибнут. В самом начале изменения клеток имеют характер обратимых , паранекротических. Отличие состоит в том, что после снятия воздействия они не исчезают, а прогрессируют. Явным признаком клеточной гибели является активация внутриклеточных гидролитических ферментов. Они активируются в гиалоплазме и начинают расщепление белков, липидов. При этом разрешается внутриклеточная мембрана. Это приводит к лизису.
Апопто́з (греч. απόπτωσις — опадание листьев) — программируемая клеточная смерть, регулируемый процесс самоликвидации на клеточном уровне, в результате которого клетка фрагментируется на отдельные апоптотические тельца, ограниченные плазматической мембраной. Фрагменты погибшей клетки обычно очень быстро (в среднем за 90 минут[1])фагоцитируются макрофагами либо соседними клетками, минуя развитие воспалительной реакции. Морфологически регистрируемый процесс апоптоза продолжается 1—3 часа.[2]Одной из основных функций апоптоза является уничтожение дефектных (повреждённых, мутантных, инфицированных) клеток. В многоклеточных организмах апоптоз к тому же задействован в процессах дифференциации и морфогенеза, в поддержании клеточногогомеостаза, в обеспечении важных аспектов развития и функционирования иммунной системы. Апоптоз реализует гены летальности, функции которых активируют внеклеточные факторы, среди которых преимущественное значение имеют оксид азота и цитоксины. Гены, вызывающие апоптоз и блокирующие его, локализуются в ядре, на мембранах ЭПС и митохондриях. Изменения начинаются с ядра. Гетерохроматин собирается в крупные электронно-плотные агрегаты , мигрирующие в цитоплазму. Поверхность клетки вследствие гибели элементов цитоскелета становится неровной, ЭПС и митохондрии вакуолизируются и их мембраны фрагментируются. Процесс кончается образованием апоптозных тел, которые поглощаются макрофагами.
Некро́з (от греч. νεκρός — мёртвый), или омертве́ние — это патологический процесс, выражающийся в местной гибели ткани в живом организме в результате какого-либо экзо- или эндогенного её повреждения. Некроз проявляется в набухании, денатурации и коагуляции цитоплазматических белков, разрушении клеточных органелл и, наконец, всей клетки. Наиболее частыми причинами некротического повреждения ткани являются: прекращение кровоснабжения (что может приводить к инфаркту, гангрене) и воздействие патогенными продуктами бактерий или вирусов (токсины, белки, вызывающие реакции гиперчувствительности, и др.).
№23
Способы и уровни адаптации клетки
Молекулярный уровень – соматическая полиплоидизация клетки. Субклеточный уровень – гиперплазия цитоплазматических органелл. При физических нагрузках в мышечных волокнах увеличивается количество миофибрилл. Клеточный уровень – осуществляется путем увеличения числа клеток (пролиферация), замена одних клеток на другие (метаплазия).
№24
Информация положения, детерминация, дифференцировка и специализация клеток.
Внешняя среда, устанавливающая число и качество работающих генов, называется информацией положения или позиционной формацией. (сигналы, сообщающие клетке об ее положении).
Позиционная информация может быть: химической (для полной регенерации кости необходимо поступление достаточного количества кислорода). При неправильном наложении гипсовой повязки развивается ложный сустав. Гальванический фактор: обеспечивает направленность векторов гальванических полей, по которым идет рост конечностей. Контактная информация (результат взаимодействия м/д собой отдельных клеток и клеточных групп. Совокупность факторов, обеспечивающих эмбриональную индукцию, которая реализуется чаще всего через контактное взаимодействие, например влияние хорды на развитие НС.
При детерминации тканевых клеток происходит стойкое закрепление их свойств, вследствие чего ткани теряют способность к взаимному превращению (метаплазии). В биологиидетерминация — это процесс определения дальнейшего пути развития клеток.
Дифференцировка клеток — процесс реализации генетически обусловленной программы формирования специализированного фенотипаклеток, отражающего их способность к тем или иным профильным функциям. Иными словами, фенотип клеток есть результат координированнойэкспрессии (то есть согласованной функциональной активности) определённого набора генов.
В процессе дифференцировки менее специализированная клетка становится более специализированной. Например, моноцит развивается вмакрофаг, промиобласт развивается в миобласт, который образуя синцитий, формирует мышечное волокно. Деление, дифференцировка иморфогенез— основные процессы, путём которых одиночная клетка (зигота) развивается в многоклеточный организм, содержащий самые разнообразные виды клеток. Дифференцировка меняет функцию клетки, её размер, форму и метаболическую активность.
№25
Компетентные и коммитированные клетки, конститутивные и индуцибельные гены.
Клетки, способные реагировать на информацию положения, становятся компетентными. Под ее влиянием геном разделяется на гены экспрессированные и репрессированные. Экспрессированные – рабочие, репрессированные – нерабочие. Среди экспрессированных генов имеются конститутивные, однотипные для всех клеток данного вида. Конститутивные гены отвечают за общие признаки жизни и организуют обмен веществ. Другие экспрессрованные гены называются индуцибельными. Они специфичны для клеток, т.е. определяют характерные для нее структурные и функциональные признаки.
№26
Происхождение тканей. Теории тканевой эволюции; роль факторов внешней среды
Принято считать, что в организме имеется 4 первичные функции и в отношении каждой формируется присущий только ей тип ткани (А.А. Заварзин)
Функции:
1 – Пограничная или разграничительная, в направлении ее формируются ткани, имеющие прямой контакт с внешней средой, выполняют защиту от внешних воздействий, получают из внешней среды трофический материал и выводят из нее остаточные продукты метаболизма. Это эпителиальные ткани.
2 – Вторая функция организует внутреннюю среду. В направлении этой функции образуются ткани, формирующие мягкие и твердые скелеты, основу интерстиция органов, защитные и циркуляторные системы организма. Эти ткани развиваются из одного источника – мезенхимы и поэтому называются мезенхимными или тканями внутренней среды, соед. тканями.
3 – Третья функция обеспечивает подвижность, является одним из главных признаков жизни. В направлении ее формируются мышечные ткани. Они делают возможным перемещение тела в пространстве и определяют моторику внутренних органов.
4 – Четвертая функция обогащает живые системы способностью отвечать на раздражители адекватной реакцией и передавать возбуждение на исполнительные органы. В направлении этой функции развивается нервная ткань.
4 типа тканей, возникшие у первых многоклеточных, в последствии у всех представителей животного мира развивались параллельно. Это значит, что у современного животного и человека имеется 4 типа тканей.
№27
Определение и классификация тканей.
Ткань – исторически сложившаяся система клеточных и неклеточных элементов, объединенных общностью происхождения, строения и функций.
Первая группа: ткани общего характера : эпителиальные , мезенхимные. Вторая группа (спец.ткани) мышечные и нервные.
Первая группа: камбиальные ткани, имеющие стволовые клетки. Эти ткани обладают хорошей органотропической регенерацией. Время их жизни ограничено днями (эпителий) и несколькими годами (нерв.клетки). Клетки гибнут путем апоптоза, восстанавливаются с помощью физиологической регенерации. Ткани одного характера обладают широкой структурной и функциональной лабильностью. Трансформация одного вида тканей в другой возможна в пределах типа. Эту перестройку называют метаплазией. Метаплазия не имеет места в специальных тканях, в отсутствие стволовых клеток их клеточные элементы не восстанавливаются. Все ткани имеют дифферонную организацию. Дифферон – группа клеток, однонаправлено развивающихся из стволовой клетки. Диффероны характерны для тканей общего характера.
|
Эпителиальная |
Соединительная |
Мышечная |
Нервная |
|
Много клеток, межклеточного вещества практически НЕТ. Покрывают ВСЕ поверхности. Образует ВСЕ железы |
Мало клеток, много межклеточного вещества. Образует ВСЕ остальное кроме поверхностей, желез, мышц и нервов. |
Возбуждается и сокращается |
Возбуждается и Передает нервные импульсы |
№28
Эпителий – определение и общая характеристика, функции.
Эпителиальные ткани – группа пограничных клеток, выполняющих в организме защитную, всасывательную и выделительную функции. Эпителиальные ткани — древнейшие гистологические структуры, которые в фило- и онтогенезе возникают первыми. Основное свойство эпителиев — пограничность. Эпителиальные располагаются на границах двух сред, отделяя организм или органы от окружающей среды.
Эпителии, как правило, имеют вид клеточных пластов и образуют наружный покров тела, выстилку серозных оболочек, просветов органов, сообщающихся с внешней средой во взрослом состоянии или в эмбриогенезе. Через эпителии осуществляется обмен веществ между организмом и окружающей средой. Важной функцией эпителиальных тканей является защита подлежащих тканей организма от механических, физических, химических и других повреждающих воздействий. Некоторые эпителии специализированы на выработке специфических веществ — регуляторов деятельности других тканей организма. Производными покровных эпителиев являются железистые эпителии. Особый вид эпителия — эпителий органов чувств.
Эпителии развиваются с 3-4-й недели эмбриогенеза человека из материала всех зародышевых листков. Некоторые эпителии, например эпидермис, формируются как полидифферонные ткани, так как в их состав включаются клеточные диффероны, развивающиеся из разных эмбриональных источников (клетки Лангерганса, меланоциты и др.).
Эпителий не имеет кровеносных сосудов, питание осущ. диффузно, через базальную мембрану.Э. не имеет межклеточного вещества между клетками. Клетки связаны с помощи десмосом, плотных контактов и др.
№29
Классификация эпителиальных тканей.