

4.1. Распространение потенциала действия
При распространении потенциала действия отмечается полное возбуждение каждой точки нервного волокна, так что амплитуда потенциала действия везде одна и та же. Протекающие по принципу «всё или нечего» процессы возбуждения в разных участках мембраны связаны посредством электротонического распространения раздражающих токов вдоль волокна. Ионы натрия, которые входят в волокно в возбужденном участке мембраны, служат источником тока для возникновения деполяризующего электротонического потенциала в соседнем, еще не возбужденном участке. Когда эта деполяризация достигает порога, она вызывает в этом участке возбуждение. Таким образом, состояние возбуждения распространяется посредством электротонической связи от возбуждаемых участков мембраны к еще не возбужденным. Однако такое распространение возбуждение довольно медленное. Гораздо быстрее потенциал действия распространяется в миелинизированных нервных волокнах. Это обусловлено особенностям строения миелинизированных волокон: только очень короткие участки этих волокон (перехваты Ранвье) покрыты обычной клеточной мембраной. Поэтому при проведении ток, по существу, не идет через мембрану межперехватных участков, и потенциал действия от одного перехвата Ранвье к другому распространяется через межперхватные участки. Время проведения через межперехватные участки практически равно нулю – возбуждение перескакивает от одного перехвата к следующему (сальтаторное проведение).
4.2. Межклеточная передача возбуждения
Как мы уже отмечали выше, по нервной клетке информация передается в виде потенциалов действия. Передача информации к соседней клетке происходит через специальные межклеточные контакты – синапсы. Синапсы подразделяются нахимическиеиэлектрические. В химических синапсах проведение осуществляется благодаря особым веществам – медиаторам, а в электрических синапсах решающее значение для проведения имеет специфическое распределение токов. Нами будут рассмотрены синапсы первого типа, поскольку именно они обеспечивают сложное взаимодействие клеток в организме человека.


ПД
Синаптические пузырьки
Митохондрии
Синаптическая бляшка






















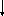

Медиатор






Постсинаптические рецепторы

Синаптическая щель

Постсинаптическая мембрана
Рис. 3. Межнейронный синапс.
На рисунке 3 показаны важнейшие компоненты химического синапса. Распространяющийся по аксону потенциал действия вызывает деполяризацию пресинаптической мембраны, что приводит к высвобождению медиатора и поступлению его в синаптическую щель. Медиатор диффундирует к плазматической мембране постсинаптической клетки (постсинаптической мембране), где происходит его взаимодействие с рецепторами. Рассмотрим эти процессы на молекулярном уровне несколько подробнее. Для этого нам потребуется, прежде всего информация о химическом строении медиаторов и их функции. Одним из наиболее распротсраненных медиаторов является ацетилхолин, который относится к числу возбуждающих медиаторов. К той же группе медиаторов относят и моноаминыдофамин, серотонин, норадреналин, адреналин. Последние два объединены в группукатехоламинов. Среди медиаторов имеются и аминокислоты: γ-аминомасляная кислота и глицин. Оба этих медиатора оказывают тормозящее воздействие на постсинаптические клетки. Аминокислоты, моноамины и ацетилхолин представляют собой группу так называемых «классических» медиаторов. Все они – низкомолекулярные соединения, образующиеся в качестве промежуточных продуктов метаболизма. Каждый из них связывается со специфическим рецепторов постсинаптической мембраны, в результате чего повышается ее проводимость – либо для натрия (и калия) в случае передачи возбуждения, либо для калия и хлора с развитием торможения. Кроме «классических» медиаторов в межклеточном взаимодействии участвуют более сложные по химическому строению и обладающие большей молекулярной массой вещества – пептиды. К важнейшим пептидным медиаторам относят мет-энкефалин, лей-энкефалин, вещество Р, ангиотензин II, вазоактивный кишечный (интерстинальный) пептид, соматостатин, рилизинг-гормон лютеотропного гормона (люлиберин или ЛГРГ). Медиаторы накапливаются в синаптических пузырьках, ассоциированных с участками пресинаптической мембраны. Эту ассоциацию можно рассматривать как аппарат, специализированный для экзоцитоза, т.е. для выброса медиаторов в синаптическую щель. При этом медиатор поступает в синаптическую щель в виде квантов, соответствующих объему одного пузырька. Небольшое количество медиатора выделяется и в состоянии покоя, но пик поступления медиатора в синаптическую щель связан с деполяризацией пресинаптической мембраны – потенциал действия пресинаптического окончания ведет к почти синхронному высвобождению квантов медиатора, которое приводит к генерированию потенциала в постсинаптической мембране. Ключевую роль в квантовом высвобождении медиатора играетвходящий ток ионов кальция.
Поступивший в синаптическую щель медиатор взаимодействует с рецептором постсинаптической мембраны, вызывая изменение ее проводимости. Механизм этого взаимодействия описан в рамках теории молекулярной рецепции – раздела биокинетики. Несколько слов об истории вопроса. В начале XX-го века П.Эрлих и И.И.Мечников сформулировали теорию боковых цепей (Нобелевская премия, 1908). Согласно этой теории на клеточной поверхности имеются особые химические соединения, которые образуют «боковые цепи». Химические соединения способны взаимодействовать с этими боковыми цепями. В результате взаимодействия происходит клеточное распознавание веществ и формирование клеточного ответа. П.Эрлих назвал эти боковые цепи рецепторами (от латинского recipere– получать, узнавать). Он сформулировал основной постулат рецепторного механизма действия химических соединений: “вещество не действует, если не фиксировано” (“corporanonagunnisifixata”). Первое количественное описание величины ответа клетки в зависимости от концентрации добавленного химического соединения было предпринято А. Кларком в 1926г. Аналогично тому, как это сделано в ферментативной кинетике, Кларк предположил, что химические вещества находятся в избытке по отношению к рецепторам. Кроме того, он предположил, что взаимодействие химических веществ с рецепторами обратимо. В настоящее время химические вещества, способные взаимодействовать с рецепторами, принято называтьлигандами( от латинскогоligo– связываю). Такое взаимодействие лигандов с рецепторами, согласно Кларку записывается так:
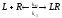 (ф.2),
(ф.2),
где L
–лиганд; R
– рецептор; LR
– комплекс лиганда с рецептором или
лиганд-рецепторный комплекс;
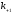 и
и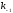 - константы скоростей образования и
распада лиганд-рецепторных комплексов.
- константы скоростей образования и
распада лиганд-рецепторных комплексов.
Согласно современным представлениям, большинство биологически активных веществ и лекарств оказывает воздействие на организм, регулируя функции клеток. При этом многие вещества не проникают в клетку, а оказывают свое действие опосредованно, взаимодействуя на клеточной мембране со специфическими молекулами, называемыми рецепторами. Рецепторы способны специфически распознать биологически активное вещество, обратимо или необратимо взаимодействовать с ним и передавать специфический сигнал об этом взаимодействии в клетку. Под рецепторами понимают макромолекулы, расположенные на цитоплазматической мембране клеток или внутриклеточно, способные специфически взаимодействовать с ограниченным набором биологически активных веществ (лигандов) и трансформировать сигнал об этом взаимодействии в специфический клеточный ответ. Все лиганды делятся на агонистыиантогонисты.Агонистами называют лиганды, которые, связываясь с рецепторами данного типа, активно вызывают клеточный ответ. В синапсе в качестве агонистов могут выступать как медиаторы, так и другие вещества. Это обусловлено тем, что специфичность мембранных рецепторов не абсолютна. Так, в холинергическом синапсе в качестве агонистов могут выступать карбамилхолин и суберилдихолин. Другие вещества, также связывающиеся с рецепторами медиаторов, но не столь эффективно изменяющие мембранную проницаемость, называютсячастичными агонистами.Антагонистами называются лиганды, которые связываются с рецепторами и не вызывают активного клеточного ответа. Антагонисты препятствуют связыванию агонистов с рецепторами, угнетая клеточные функции. Связывание их может быть обратимым: спустя некоторое время антагонист отделяется от рецептора. Такие вещества называютконкурентными антагонистами, так как они конкурируют с медиаторами и их антагонистами за участки связывания. Хорошо известный конкурентный антагонист ацетилхолина в нервно-мышечном синапсе – яд кураре (d-тубокурарин). По мере повышения концентрации он блокирует все больше рецепторов, и эффект ацетилхолина ослабляется из-за уменьшения доступных мест связывания. Это приводит к уменьшению потенциала концевой пластинки и параличу мышцы.
С учетом всего вышесказанного формула Кларка будет выглядеть следующим образом:
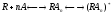 (ф.3),
(ф.3),
где n– число молекул
агониста, связывающихся с одним
рецептором, а переход от состояния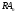 к состоянию
к состоянию отражает сдвиг проводимости, т.е. открытие
постсинаптического ионного канала.
Рецепторы делятся на несколько групп.
К первой из них относятсярецепторы
– ионные каналы. Наиболее часто эти
рецепторы представляют собой несколько
белков, образующих ионный канал. В
неактивном состоянии эти белки расположены
так, что ионный канал закрыт. Связывание
одной или нескольких белковых молекул
с лигандом приводит к открытию канала
и прохождению ионов в клетку или выход
из клетки. В этом случае ионы могут
выступать в качестве вторичных
мессенджеров.Вторичными мессенджерами(от английскогоmessage–
сообщение) называются внутриклеточные
молекулы, осуществляющие сопряжение
рецепторов с внутриклеточными эфферентными
системами (ферментами, ионными каналами,
геномом и т.д.). В результате
лиганд-рецепторного взаимодействия
концентрация вторичных мессенджеров
в клетке меняется (возрастает или
убывает). Известны четыре системы
вторичных посредников:
отражает сдвиг проводимости, т.е. открытие
постсинаптического ионного канала.
Рецепторы делятся на несколько групп.
К первой из них относятсярецепторы
– ионные каналы. Наиболее часто эти
рецепторы представляют собой несколько
белков, образующих ионный канал. В
неактивном состоянии эти белки расположены
так, что ионный канал закрыт. Связывание
одной или нескольких белковых молекул
с лигандом приводит к открытию канала
и прохождению ионов в клетку или выход
из клетки. В этом случае ионы могут
выступать в качестве вторичных
мессенджеров.Вторичными мессенджерами(от английскогоmessage–
сообщение) называются внутриклеточные
молекулы, осуществляющие сопряжение
рецепторов с внутриклеточными эфферентными
системами (ферментами, ионными каналами,
геномом и т.д.). В результате
лиганд-рецепторного взаимодействия
концентрация вторичных мессенджеров
в клетке меняется (возрастает или
убывает). Известны четыре системы
вторичных посредников:
аденилатциклаза – циклический аденозин – моно – фосфат (цАМФ);
гуанилатциклаза – циклический гуанозин – монофосфат (цГМФ);
фосфолипаза С - инозитол – три – фосфат (Ифз);
ионизированный кальций.
В значительной части клеток организма присутствуют или могут образовываться почти все из перечисленных выше вторичных посредников (исключением является цГМФ). Это обуславливает различные взаимосвязи между вторичными посредниками: 1) равноправное участие, когда разные посредники необходимы для медиаторного эффекта; 2) один из посредников является основным, а другой лишь способствует реализации эффектов первого; 3) посредники действуют последовательно (например, инозитол – 3- фосфат обеспечивает освобождение кальция, диацилглиццерол облегчает взаимодействие кальция с протеинкиназой С); 4) посредники дублируют друг друга для обеспечения избыточности с целью надежности регуляции; 5) посредники являются антагонистами, т.е. один из них включает реакцию, а другой тормозит (например, в гладких мышцах сосудов инозитол–3–фосфат и кальций реализуют их сокращение, а цАМФ – расслабление).
Эффект от воздействия биологически активного вещества во многом зависит от того, с каким типом рецепторов он образовал лиганд-рецепторную связь. В группу рецепторов – ионных каналов входят, в частности, Н-холинорецепторы. Вторая группа рецепторов образована рецепторами, сопряженными с ферментами. Для этих рецепторов характерна каталитическая активность одной из образующих их молекул белка. Лиганд – рецепторное взаимодействие приводит к изменению каталитической активности данной субъединицы, а значит и синтеза внутриклеточных вторичных мессенджеров. По такому механизму функционируют рецепторы факторов роста, инсулиновые рецепторы и др. К третьей группе рецепторов относятся рецепторы, сопряженные сG-белками.G-белки – это особые ГТФ-связывающие белки, расположенные на внутриклеточной стороне цитоплазматической мембраны. Посредством этих белков рецепторы могут изменять активность внутриклеточных ферментативных систем и вызывать открытие ионных каналов. КG-сопряженным рецепторам относятся, например, β-адренорецепторы.G-белки представляют собой гетерогенную популяцию, различающиеся за счёт α-субъединицы. Имеется насколько видовG-белков:

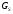 - белки. Оказывают стимулирующее влияние
на аднилатциклазу, оказывает ингибирующее
влияние на медленный тип кальциевых
каналов.
- белки. Оказывают стимулирующее влияние
на аднилатциклазу, оказывает ингибирующее
влияние на медленный тип кальциевых
каналов.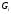 - белки. Оказывают ингибирующее
воздействие на аденилатциклазу,
активируют цАМФ-фосфодиэстеразу.
- белки. Оказывают ингибирующее
воздействие на аденилатциклазу,
активируют цАМФ-фосфодиэстеразу. -белки.
Их активация приводит к угнетению
потенциал-зависимых кальциевых каналов
и стимуляции калиевых каналов.
-белки.
Их активация приводит к угнетению
потенциал-зависимых кальциевых каналов
и стимуляции калиевых каналов. -белки.
Вызывают активацию калиевых каналов
и угнетение аденилатциклазы.
-белки.
Вызывают активацию калиевых каналов
и угнетение аденилатциклазы. -белки.
Приводят к активации фосфолипазы С.
-белки.
Приводят к активации фосфолипазы С.
Процессы, происходящие при воздействии тормозящих и возбуждающих медиаторов на рецепторы постсинаптической мембраны различны. Под воздействием возбуждающего медиатора на постсинаптической мембране возникает возбуждающий постсинаптический потенциал (ВПСП). Данные изучения ионных механизмов ВПСП указывают на то, что ВПСП представляет собой результат кратковременного повышения проводимости для мелких катионов. Другим вариантом развития событий может быть возникновение торможения. Известны два типа торможения –пресинаптической и постсинаптическое. При пресинаптическом торможении уменьшается или прекращается высвобождение медиатора из пресинаптических нервных окончаний, а при постсинаптическом торможении снижается возбудимость сомы и мембраны дендритов. Тормозящий постсинаптический потенциал (ТПСП) по своей сути представляет собой гиперполяризацию постсинаптической мембраны. Возникновение ТПСП связано с тем, что при действии тормозящего медиатора на постсинаптическую мембрану повышается проницаемость последней для ионов калия и хлора. Торможение, вызываемое ТПСП, обусловлено как гиперполяризацией мембранного потенциала, так и начальным повышением проводимости мембраны.
В качестве гистологического субстрата пресинаптического торможения удобнее всего рассматривать аксо-аксональные синапсы. В ходе проведенных различными учеными исследований было установлено, что снижение количества высвобождаемого медиатора в случае пресинаптического торможения связано с активацией именно этого вида синапсов. В механизме пресинаптического торможения большую роль играет деполяризация первичных афферентов (ДПА). Активация аксо-аксонного синапса сопровождается деполяризацией постсинаптической области. Такую деполяризацию можно зарегистрировать внутриклеточно в первичных афферентах спинного мозга, откуда она и получила свое название. ДПА сопровождается уменьшением амплитуды пресинаптического потенциала в результате инактивации натриевых каналов. В конечном счете, может происходить полная блокада проведения в пресинаптических окончаниях, сопровождающаяся высвобождением очень небольшого количество медиатора.
После того, как медиатор диффундировал через синаптическую щель, дальнейшая его диффузия из этого узкого пространства идет довольно медленно и не способна заметно снизить установившуюся концентрацию.
Важнейшим, с точки зрения физиологии, вопросом является взаимодействие между синапсами. И действительно, первый же шаг от этого элементарного уровня взаимодействия двух клеток возбудимых тканей вверх по шкале размерностей, первый же шаг к пониманию целостной функциональной организации нервной системы требует рассмотрения этого взаимодействия.
Объединяясь между собой посредством синапсов, нейроны формируют изумительные по сложности системы, чьё функционирование и определяет, в конечном счетё, высшие функции центральной нервной системы. Здесь мы рассмотрим лишь наиболее простые из этих нейронных сетей. Эти элементарные нейронные цепи служат, например, для усиления слабых сигналов, уменьшения слишком интенсивной активности, выделения контрастов, поддержания ритмов или сохранения рабочего состояния нейронов путём регулировки их входов. Сама собой напрашивается аналогия с интегральными электрическими цепями – стандартными элементами, которые выполняют наиболее часто повторяющиеся операции, и могут быть включены в схемы самых разнообразных электронных приборов. Эти простейшие нейронные системы расположены во всех отделах головного и спинного мозга и характеризуются некоторым набором структурно функциональных феноменов, к числу которых относится конвергенция, дивергенция, временная и пространственная суммация (облегчение), различные виды торможения и реверберации.
Афферентные волокна, несущие сигнал от периферических рецепторов, сначала входят в спинной мозг в составе дорсальных корешков, а затем ветвятся на много коллатералей, идущих к спинальным нервам. Таким образом один чувствительный нейрон оказывает влияние на несколько других нейронов. Это явление получило название дивергенция. Благодаря дивергенции афферентная информация поступает одновременно к разным участкам ЦНС. Тут следует отметить, что разветвление волокон дорсальных корешков является лишь частным случаем дивергенции. Это явление обнаруживается во всех отделах центральной нервной системы, что позволяет говорить о принципе дивергенции в нейронных сетях.
В качестве структурной и функциональной противоположности дивергенции мы с полным основанием можем рассматривать конвергенцию. При этом явлении несколько нейронов оказывают влияние на функцию одного нейрона. Число таких входов для одного нейрона очень велико (на мотонейроне оканчивается до 6000 коллатералей аксонов), а само явление столь широко представлено в ЦНС, что мы с полным основанием можем говорить о принципе конвергенции в нейронных связях. Благодаря конвергенции возможно интегрирование различным потоков информации на одном нейроне, что является одним из важнейших процессов, происходящих в ЦНС.
В случае, если возбуждающие постсинаптические потенциалы (ВПСП) быстро следуют друг за другом, то они суммируются благодаря своему относительно медленному временно́му ходу, достигая в конце концов порогового уровня. Это повышение возбудимости нейрона в результате последовательно возникающих ВПСП носит название временно́го облегчения. Временное облегчение ответа на активность аксона обусловлено тем, что ВПСП продолжается дольше, чем рефрактерный период аксона. Временное облегчение играет очень важную роль, потому что многие нейронные процессы имеют ритмический характер и, таким образом, могут суммироваться, давая начало надпороговому возбуждению в синапсах.
Для пространственного облегчения характерна суммация ВПСП, возникающих одновременно в разных синапсах. В результате этого возникает распространяющийся потенциал действия – процесс, который не может быть обеспечен одиночным ВПСП. Если рассматривать в целом оба вида облегчения, то можно сказать, что облегчение возникает в том случае, когда количество импульсов больше, чем сумма отдельных входов.
Выше мы уже отмечали, что кроме нейронных цепей, в которых происходит облегчение, существуют и простые тормозные цепи, для которых характерны различные типы торможения. Изучение их мы начнем свозвратного торможения. Эта форма торможения возникает в том случае, когда тормозные вставочные нейроны действуют на те же нервные клетки, которые их активируют. При этом торможение бывает тем глубже, чем сильнее было предшествующее возбуждение. Это позволяет использовать кибернетический принцип обратной связи для описания этих цепей, а само возвратное торможение рассматривать какторможение по принципу обратной связи. Частным случаем возвратного торможения являетсяторможение Реншоу. Этот вид торможение встречается в мотонейронах: мотонейроны посылают коллатерали к вставочным нейронам (клеткам Реншоу), аксоны которых в свою очередь образуют тормозные синапсы на мотонейронах.
Латеральное торможение.При данном виде торможения тормозные вставочные нейроны соединены таким образом, что они влияют не только на возбужденную клетку, но и на соседние клетки с такими же функциями, в которых возбуждение отсутствует или является более слабым. В результате в этих соседних нейронах развивается очень глубокое торможение. Поскольку при этом возбужденный нейрон оказывается со всех сторон окруженным зоной торможения, это явление также называютокружающим торможением. Латеральное торможение играет особенно важную роль в афферентных системах.
Реципрокное торможение.Для изучения данного вида торможения необходимо ввести некоторую терминологию.Гомонимныминазываются все те нейроны, которые посылают аксоны к одной и той же мышце.Мышцы-агонисты, илисинергисты, осуществляют движение в суставе в одном и том же направлении, амышцы-антагонисты -в противоположных направлениях. Нервные волокна, идущие по направлению к гомонимным мотонейронам, образуют на этих нейронах возбуждающие синапсы, а посредством вставочного нейрона – тормозные синапсы на мотонейроны мышц-антагонистов. Если повышается тонус нервных волокон, идущих к мотонейронам мышц-сгибателей, то они оказывают тормозящие влияние на мотонейрон, иннервирующий мышцу-разгибатель, действующую на тот же сустав. Это торможение имеет большой физиологический смысл, потому что обеспечивает согласование тонуса мышц и облегчает движение в суставе автоматически, без дополнительного произвольного или непроизвольного контроля