

Углеводная часть
|
|
|
|
рибоза (-Д-рибофураноза) – в составе РНК |
дезоксирибоза (-Д-2-дезоксирибофураноза) – в составе ДНК |


Нуклеиновые кислоты (НК) бывают двух типов: ДНК и РНК.
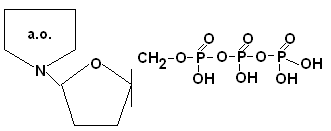
Мономерным звеном нуклеиновых кислот являются мононуклеотиды, которые состоят из 3-х компонентов: азотистого основания (АО), углеводной части и остатка фосфорной кислоты.
Состав нуклеиновых кислот
Дезоксирибоза
Таким образом, звеном нуклеиновых кислот являются мононуклеотиды, состоящие из трех химически связанных компонентов. Соединение АО с сахаром называется нуклеозидом. Он образуется из мононуклеотида путем отщепления Н3РО4 при гидролизе или под воздействием специфических факторов.
Соединение этих трех компонентов происходит за счет b-N-гликозидной связи N1- пиримидинового или N9 пуринового азотистого основания с С1 атома пентозы и за счет сложноэфирной связи С5 атома пентозы и фосфорной кислотой.
|
| |
|
5-цитидиловая кислота цитидин-5-фосфат (5-дезоксицитидиловая) 5¢-уридиловая 5¢-адениловая 5¢гуаниловая |
ЦМФ (5-дезоксицитидиловая) (5-дезоксиадениловая) (5-дезоксигуаниловая)
|
|
|
|

|
|
Если нуклеозид соединяется 1, 2 или 3 атомами Фн, то мононуклеозид называется соответственно цитидинмонофосфат – ЦМФ, цитидиндифосфат – ЦДФ, цитидинтрифосфат – ЦТФ
|
Первичная структура нуклеиновых кислот – это порядок, последовательность расположения мононуклеотидов в полинуклеотидные цепи ДНК или РНК, соединенных между собой 35-фосфодиэфирной связью.
|
|

Наиболее распространенные нуклеотиды клетки.
Нуклеотиды – это высокомолекулярные соединения (ВМС). Их молекулы имеют нитевидную форму, что обусловливает высокую вязкость их растворов. Длина молекул ДНК занимает несколько см. Общая длина ДНК в 23 парах хромосом человека около 1,5 м. Молекулы РНК короче: около 0,01 мм. Основная часть ДНК находится в ядре клетки – в составе хроматина; 0,25% ДНК – в митохондриях. РНК обнаружено во всех частях клетки. РНК состоит обычно из 2 полинуклеотидной цепи. ДНК – из двух. В редких случаях возможны двухцепочечные РНК и одноцепочечные ДНК.
Нуклеотиды в клетке могут быть в виде мононуклеотидов: АТФ, АДФ, АМФ, ЦТФ, ЦДФ и т.д.
В зависимости от вида пентозы нуклеотиды делят на 2 типа – рибонуклеотиды и дезоксирибонуклеотиды.
Дезоксирибонуклеотиды используются для образования ДНК.
Функции рибонуклеотидов более разнообразны. Они идут на образование РНК, могут выполнять роль коферментов в некоторых реакциях (трансферазных). Адениловые рибонуклеотиды входят в состав НАД, НАДФ, ФАД, КоА. Нуклеозидтрифосфаты (АТФ, ЦТФ и т.д.) содержат 2~связи, то есть связи, при гидролизе которых выделяется Е (~50 Кдж).
Вторичная структура днк. Правила Чаргаффа.
Изучая состав ДНК, Чаргафф (1949г.) установил важные закономерности соотношения АО в ДНК, что в дальнейшем помогло раскрыть их структуру.
1) сумма пуриновых оснований = сумме пиримидиновых оснований.
А
+ Г = Ц + Т или ![]()
2) соединение А = соединению Т
А
= Т или ![]()
3) соединение Г = соединению Ц
Г
= Ц или ![]()
4) сумма А и Ц = сумме Г и Т
А+Ц
= Г+Т или ![]()
5) изменчива только сумма А+Т и Г+Ц
если А+Т Г+Ц, то это АТ-тип ДНК
если наоборот, то это ГЦ-тип ДНК.
Эта разница выражается через коэффициент специфичности, который является важной характеристикой молекулы ДНК.
![]() (в
% от суммы Г+Ц от 0,54
до 0,94 для
живых, то есть преобладает АТ-тип; от
0,45 до 2,57 –
для микроорганизмов).
(в
% от суммы Г+Ц от 0,54
до 0,94 для
живых, то есть преобладает АТ-тип; от
0,45 до 2,57 –
для микроорганизмов).
Правила Чаргаффа характерны только для ДНК, но не свойственны РНК.
На основании этих правил и большого числа разнообразных экспериментальных данных в 1953 г. Д. Уотсон и Ф. Крик разработали модель ВС ДНК.
Молекула ДНК построена из двух полинуклеотидных. цепей, ориентированных антипараллельно (3′ конец одной цепи соседствует с 5′ концом второй цепи); цепи на всем протяжении связаны Н-связями. При этом каждый мононуклеотид участвует в образовании Н-связей.
Н-связи образованы между А одной цепи и Т другой, между Г одной цепи и Ц другой. Основания, образующие такие пары комплементарны друг другу, то есть между ними легче образуется Н-связь.

ПС одной цепи ДНК комплементарна ПС второй цепи, что не означает их идентичности

Обе цепи закручены вправо в спираль, имеющую общую ось. Цепи могут быть разделены только раскручиванием. Такие спирали называются плектонемическими.
Пурины и пиримидиновые основания обращены внутрь спирали. Их плоскости перпендикулярны оси спирали и параллельны друг другу, таким образом получается стопка оснований. Пентозофосфатные остовы обеих цепей обращены наружу от спирали.
Стабилизация ВС ДНК обеспечена за счет:
1) особого вида вандервальсовых сил между АЩ – так называемых стэкинг-взаимодействий;
2) водного окружения фосфатов и особенно нейтрализации их зарядов. –q стремятся оттолкнуться друг от друга, вытягивают цепь ДНК и тем самым стабилизируют ее;
3) образование двойной спирали ДНК возможно только при условии связывания фосфатов (–q) с гистонами – основными белками (+q);
4) Н-связями.
Комплекс белков с ядерной ДНК клеток называется хроматином.
Двойная спираль (-форма) Уотсона и Крика имеет размерность:
1) -спирали – 2,1 нм;
2) один виток (шаг спирали) составляет 10 пар АО; 3,4 нм;
3) высота каждого мононуклеотида 0,34 нм.
В природной ДНК хромосом строгая двуспиральность нарушается на тех участках, которые называются палиндромами (перевертышами). В таких фрагментах ДНК последовательность мононулеотидов одинакова слева направо и справа налево. Комплементарные основания палиндромов спариваются и образуют «кресты» или «шпильки». Они помогают регуляторным белкам узнавать места списывания генетического текста ДНК хромосом.
Третичная структура ДНК образуется в результате дополнительного скручивания в пространстве в виде суперспирали или изогнутой, то есть сломанной двойной спирали.
В составе хроматина ДНК сост 30-45%; гистоны 30-50%; негистиновый белок 4-33%; РНК 1,5-10%.
В электронном микроскопе хроматин напоминает бусы – шаровидные утолщения – нуклеосомы, с перемычками. Каждая нуклеосома содержит 8 молекул гистонов и намотанный на них участок ДНК, содержащий 146 пар АО. При такой укладке длина молекулы ДНК уменьшается ~ в 7 раз. Это лишь первый уровень укладки цепи. Длина молекул ДНК 3-5 см, а длина хромосом всего несколько мкм, то есть степень укорочения ДНК достигает нескольких тысяч. Это происходит в результате дополнительного скручивания нуклеосомной нитки бус.
Вторичная и третичная структура РНК
По особенностям структуры и функций различают 3 основных типа РНК:
1) Р-РНК;
2) Т-РНК;
3) М(И)-РНК.
Р-РНК – это компоненты рибосом и составляют около 80% всей РНК клетки. Существует три вида р-РНК, которые отличаются по массе и коэффициенту седиментации.
К сед. Mr число м/н
1) 28-S-р-РНК ~ 1,5 млн. до 4000
2) 18-S-р-РНК ~ 700 тыс. до 2000
3) 5-S-р-РНК ~ 30 тыс. ~ 100
Вторичная структура р-РНК – это спиралевидные участки, соединенные изогнутой одиночной цепью.
Третичная структура р-РНК является скелетом рибосомы.
Рибосомы – это субклеточные частицы с Ксед. 80S и Mr~1,5 млн. Они состоят из 2 субъединиц – большой (60S) и малой (40S); при снижении концентрации ионов Mg+2 до 0,1 мМоль 80S-частица распадается на субъединицы. Каждая из субъединиц содержит р-РНК и белки. р-РНК выполняет роль каркаса для объединения белков в определенном объеме.
м(и)-РНК – образуется в цитоплазме клетки из предшественника – пре-м-РНК. Последняя имеет копии палиндромов ДНК, поэтому ее вторичная структура содержит «шпильки» и линейные участки. При созревании пре-м-РНК «шпильки» отсекаются ферментами и образуется м-РНК. Кодовым элементом м-РНК является триплет нуклеотидов – кодон. Каждый кодон соответствует определенной аминокислоте. Вторичная структура м-РНК – это изогнутая цепь, а третичная – подобна нити, намотанной на катушку, роль которой играет особый транспортный белок – информофер. м-РНК составляет ~ 2-5% всей РНК-клетки.
т-РНК – составляет ~ 15% всей РНК клетки, имеется несколько десятков видов т-РНК, отличаются они ПС. Mr т-РНК ~ 25 000 ДА. Находятся в цитоплазме.
ВС т-РНК имеет форму «клеверного листа». т-РНК построено из одной полинуклеотидной цепи. Но в этой цепи имеются участки в виде двойной спирали, образованные комплементарными нуклеотидными парами А…V и G…С, они чередуются с неспирализованными участками. Все т-РНК имеют схожую ВС, отличаются ПС.
«Клеверный лист» содержит 5 спирализованных стеблей, 4 из которых заканчиваются петлями из неспирализованных нуклеотидов. В центре молекулы находится неспирализованная часть

1. Акцепторный участок – состоит их 4-х линейных нуклеотидов, три из которых имеют одинаковую последовательность ССА. Гидроксил 3′-ОН аденозина свободен. К нему присоединяется СООН-группа аминокислоты. Связанную с 3′-концом аминокислоту т-РНК доставляет к рибосомам, где происходит синтез белка.
2. Антикодоновая петля – образуется 7 нуклеотидами. Содержит специфический для каждой т-РНК триплет нуклеотидов – антикодон. Он по принципу комплементарности спаривается с кодоном м-РНК. Кодон-антикодоновое взаимодействие определяет порядок расположения аминокислот в ППЦ во время ее сборки на рибосоме.
3. Ψ-петля (псевдоуридиловая) – состоит из 7 нуклеотидов и обязательно содержит минорный компонент – остаток псевдоуридиловой кислоты (до 15% минор нуклеотидов). Полагают, что именно этой петлей т-РНК взаимодействует с рибосомой.
4. Д-петля (дигидроуридиловая) – состоит из 8-12 нуклеотидов, в ней всегда соединяются несколько остатков минорного компонента дигидроуридила. Участвует в узнавании аминокислотой своей т-РНК.
5. Во всех т-РНК имеется добавочная петля разной длины и размеров у разных т-РНК. Функции ее мало изучены. Полагают, что с ее помощью уравнивается длина т-РНК.
6. Третичная структура т-РНК – изучена у т-РНКфен и т-РНКасп полученных из дрожжей. Она очень компактна, образуется путем сближения отдельных частей «клеверного листа», имеет форму локтевого сгиба, при этом лепестки петель заворачиваются на тело молекулы, удерживаясь дополнительно Ван-дер-Ваальсовыми силами. Доказано, что т-РНК остаются неизменными вот уже на протяжении 500 млн. лет.
ЛЕКЦИЯ 5