1.5. Распространение потенциала действия
ПД – волна возбуждения, распространяющаяся по мембранам нервных и мышечных клеток. ПД обладает способностью к самораспространению. ПД – обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам. Синоним ПД – нервный импульс, спайк.
Сложная информация о действующих на организм раздражениях кодируется в виде отдельных групп ПД – рядов. Амплитуды и длительности отдельных ПД постоянны (закон “все или ничего”), а частота ПД и их количество в ряду зависят от интенсивности раздражения. Такой способ кодирования и передачи информации является наиболее помехоустойчивым.
В живых организмах информация может передаваться и гуморальным путем.
Преимущества ПД.
Информация более целенаправленна.
Передается быстро.
Адресат точно известен.
Информация может быть точнее закодирована.
ПД распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками. Из-за перезарядки мембраны во время генерации ПД возникает способность к самораспространению ПД. Возникнув на одном участке, он является стимулом для соседних участков.
Рефрактерность во время возбуждения в данном участке мембраны, обусловливает поступательное движение ПД.
Распространение одиночного потенциала действия само по себе не требует энергетических затрат. Однако восстановление исходного состояния мембраны и поддержание ее готовности к проведению нового импульса связано с затратой энергии на работу натрий-калиевого насоса.

Рис. 7. Распространение нервного импульса посредством локальных токов (Hodgkin, 1964).
I– распространение импульса в немиелинизированном нерве;II– сальтаторное проведение в миелинизированном нерве от одного перехвата Ранвье к другому.
Конкретные особенности распространения возбуждения связаны со строением мембраны клетки, нервных волокон.
По мембранам мышечных клеток и в безмякотных нервных волокнах возбуждение распространяется непрерывно вдоль всей мембраны (рис. 7.I).
В волокнах, покрытых миелиновой оболочкой, образованной шванновскими клетками, ПД может распространяться только скачкообразно (сальтаторно), перепрыгивая через участки волокна, покрытые миелином, с одного перехвата Ранвье на другой (рис. 7.II).
Причины сальтаторного проведения:
1) В перехватах Ранвье, свободных от миелина, сопротивление электрическому току
минимально.
2) Порог раздражения в перехватах Ранвье минимальный.
3) Велика плотность натриевых каналов на мембране перехвата (12000 на 1 мкм2– это
значительно больше, чем в любом другом участке волокна).
4) Возбуждение, возникающее в одном перехвате Ранвье, вызывает смещение ионов во внешней и внутренней средах данного волокна и этого смещения достаточно, чтобы вызвать возбуждение в соседнем участке.
Особенности декрементного и бездекрементного распространения волны возбуждения. Декремент – постепенное ослабление.
Декрементное проведение:
1) Наблюдается в безмиелиновых волокнах.
2) Свойственно волокнам, которые иннервируют внутренние органы, обладающие низкой
функциональной активностью.
3) Скорость распространения возбуждения невелика и определяется диаметром волокна.
Бездекрементное проведение:
1) ПД проходит весь путь от места раздражения до места реализации без затухания.
2) Характерно для миелиновых волокон, которые передают сигналы к органам,
обладающим высокой реактивностью.
3) Время проведения возбуждения обратно пропорционально длине между перехватами
Ранвье.
Скорость проведения возбуждения по нервному волокну зависит от диаметра волокон и наличия перехватов Ранвье. Диаметр волокна определяет характеристики электротонического (зависимого от полюсов) распространения мембранных токов. При увеличении диаметра волокна продольное сопротивление его внутренней среды, определяемое площадью поперечного сечения, снижается относительно сопротивления мембраны. В результате электротонические токи распространяются на большее расстояние и возрастает скорость проведения. Длина участков между перехватами Ранвье различна и также зависит от толщины нервного волокна: чем оно толще, тем длиннее расстояние между перехватами.
Классификация нервных волокон по Эрлангеру и Гассеру (1937)
|
Группа волокон |
Диаметр волокна, мкм |
Скорость проведения, м/с |
|
Aa Ab Ag Ad |
18 – 22 8 – 12 4 – 8 1 – 4 |
70 – 120 40 – 70 15 – 40 5 – 15 |
|
B |
1 – 3 |
3 – 4 |
|
C |
0,5 – 1 |
0,5 – 2 |
Сложный потенциал действия нервных стволов.
Нервные волокна бывают объединены в тракты и образуют нервные стволы, в большинстве своем смешанные, содержащие волокна разного размера и типа с различной скоростью проведения.
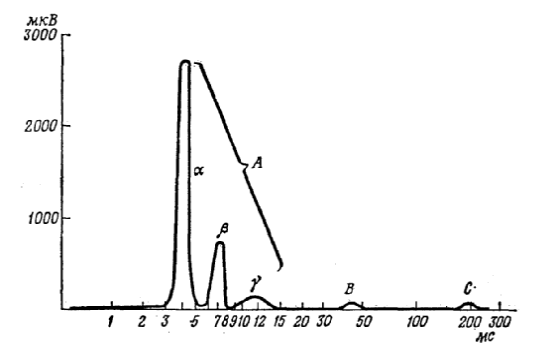
Одиночный электрический удар, достаточно сильный, чтобы возбудить все волокна смешанного нерва, вызывает на некотором расстоянии от точки стимуляции серию электрических ответов (рис. 8). Эрлангер и Гассер показали, что все эти комплексы представляют собой ответы разных типов волокон в смешанном нерве. Слабый стимул, неспособный возбудить тонкие волокна с низкой возбудимостью (Аβ), вызовет ответ только в крупных (Аα) волокнах.
|
Рис. 8. Реконструкция сложного потенциала действия. Показаны относительные размеры и временные соотношения различных компонентов. Спайк А с его вершинами – альфа, бета и гамма – проводится по волокнам А-группы; В-волна – по волокнам В-группы с более медленным проведением; С-волна – по волокнам с самым медленным проведением. Седалищный нерв лягушки.
|
|