

биохимия краткий курс часть 1
.pdfЗатем следует еще одна реакция фосфорилирования за счет АТФ, катализирует ее фосфофруктокиназа. В ходе этой реакции фруктозо-6-фосфат превращается в фруктозо-1,6-бисфосфат.
Этот метаболит далее расщепляется на 2 триозофосфата: ФГА – фосфоглицериноый альдегид или глицероальдегид-3-фосфат и диоксиацетонфосфат (ДОАФ), реакцию катализирует альдолаза.
В последующих реакциях гликолиза используется только ФГА, поэтому ДОАФ превращается с участием фермента триозофосфатизомеразы в ФГА. На этом заканчиваетсяподготовительный этап.
41
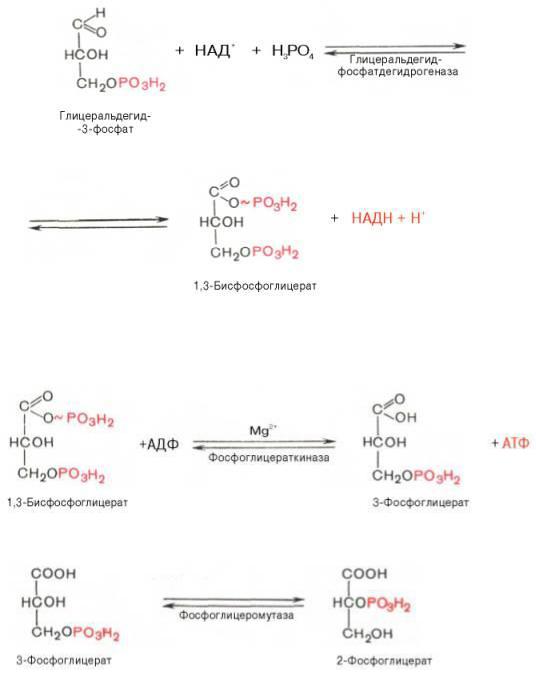
Основной этап аэробного распада включает реакции, связанные с синтезом АТФ. Сначала происходит окисление ФГАприучастии ФГАдегидрогеназы. Этот фермент является НАД+-зависимымым. Восстановленный НАД+ затем окисляется в дыхательной цепи (перенос НАДН в митохондрии, где находится дыхательная цепь, происходит при участии челночных механизмов, это связано с тем, что митохондриальная мембрана непроницаема для НАДН). Кроме того, свободная энергия окисления этой реакции концентрируется в макроэргической связи продукта реакции. Продуктом реакции является 1,3-бисфосфоглицерат.Для фосфорилирования используется Н3РО4.
Вследующей реакции высокоэнергетический фосфат передается на АДФ
собразованием АТФ. Катализирует реакцию фосфоглицераткиназа. АТФ в данной реакции образуется путем субстратного фосфорилирования. Продуктом реакции является 3-фосфоглицерат.
Вследующей реакции происходит перенос фосфата из 3 положения во второе с образованием 2-фосфоглицерата.
Из него при отщеплении воды образуется фосфоенолпируват.
42
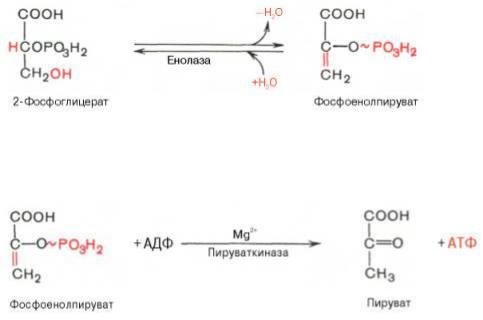
Фосфоенолпируват – макроэргическое соединение, фосфатная группа которого в следующей реакции переносится на АДФ с образованием АТФ при участии пируваткиназы (это вторая реакция субстратного фосфорилирования). Продуктом реакции является пируват.
Во второй стадии пируват превращается в ацетил-КоА, который сгорает в цикле Кребса и дыхательной цепи (третья стадия) до СО2 и Н2О.
Всего при аэробном окислении глюкозы выделяется 32 АТФ (в случае работы малат-аспартатного челночного механазма.При работе
глицерофосфатной челночной системы образуется 30 АТФ).
Анаэробный гликолиз представляет собой распад глюкозы в анаэробных условиях (условиях недостаточного снабжения кислородом), поэтому он не зависит от работы митохондриальной дыхательной цепи. АТФ в гликолизе образуется путем субстратного фосфорилирования. Конечным продуктом является лактат.
Все реакции анаэробного гликолиза (11 реакций) протекают в гиалоплазме. Первые 10 реакций до пирувата идентичны аэробному распаду. В последней реакции происходит восстановление пирувата в лактатцитозольным НАДН. (рис. 6). Реакцию эту катализирует лактатдегидрогеназа. С помощью этой реакции обеспечивается регенерация НАД+ из НАДН без участия дыхательной цепи в ситуациях, связанных с недостаточным снабжением клеток кислородом. Роль акцептора водорода от НАДН (подобно кислороду в дыхательной цепи) выполняет пируват. Таким образом, значение реакции восстановления пирувата заключается не в образовании лактата, а в том, что данная цитозольная реакция обеспечивает регенерацию НАД+.
К тому же лактат не является конечным продуктом метаболизма, удаляемым из организма (лактат – тупиковый метаболит). Он выводится в кровь и утилизируется: либо превращается в глюкозу (75%), либо при доступности кислорода окисляется до СО2 и Н2О (25%).
Энергетический выход анаэробного гликолиза составляет 2 АТФ. Недостатками анаэробного гликолиза являются: малый энергетический
выход по сравнению с аэробным распадом и накопление лактата, что приводит к ацидозу, усталости.
43
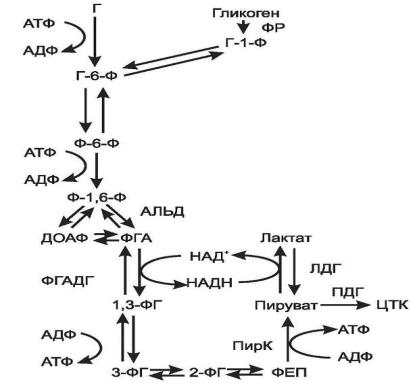
Рис.6. Гликолиз
Достоинства гликолиза: он полезен при выполнении кратковременной работы, является единственным источником энергии для эритроцитов (в них отсутствуют митохондрии), а также необходим в разных органах при дефиците кислорода (наложение жгута, падение давления, нарушение кровотока).
Регуляция.Регуляторными ферментами гликолиза являются: гексокиназа, фосфофруктокиназа и пируваткиназа. Аллостерическими ингибиторами являются АТФ и цитрат. Стимулирует гликолиз инсулин, а тормозят глюкагон, глюкокортикостероиды. Катехоламины тормозят гликолиз в печени и активируют в мышцах.
4.4. ГЛЮКОНЕОГЕНЕЗ
Некоторые ткани нуждаются в постоянном поступлении глюкозы. Когда поступление углеводов в составе пищи недостаточно, содержание глюкозы в крови некоторое время поддерживается в пределах нормы за счет расщепления гликогена в печени. Однако запасы гликогена в печени невелики, они практически полностью исчерпываются после суточного голодания. В этом случае в печени начинается глюконеогенез (ГНГ) – процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. В первую очередь это важно для головного мозга (он не может обеспечивать потребности в энергии за счет окисления жирных кислот) и эритроцитов – в них аэробный путь распада веществ невозможен из-за
44
отсутствия митохондрий. Процесс в основном (на 90%) протекает в печени и менее интенсивно в корковом веществе почек и слизистой тонкого кишечника.
Субстратами ГНГ являются лактат, аминокислоты, глицерин. Аминокислоты включаются в ГНГ при длительном голодании или продолжительной мышечной работе. Глицерин освобождается в процессе липолиза из жировой ткани в период голодания или при длительной физической нагрузке. Лактат – продукт анаэробного гликолиза. Его источником в покое являются эритроциты, а при работе – мышцы. Таким образом, лактат используется в глюконеогенезе постоянно. Большинство реакций глюконеогенеза протекает за счет обратимых реакций гликолиза и катализируется теми же ферментами. Однако три реакции гликолиза необратимы. На этих стадиях реакции ГНГ протекают другими путями (обходные пути ГНГ).
Первая необратимая реакция – образование из пируватафосфоенолпирувата. В ГНГ это происходит в ходе двух реакций. Пируват в митохондриях карбоксилируется с образованием оксалоацетата при участии пируваткарбоксилазы – биотинзависимого фермента. Реакция протекает с использованием энергии АТФ. Фосфоенолпируваткарбоксикиназа с затратой энергии ГТФ превращает оксалоацетат в фосфоенолпируват. Дальнейшие реакции ГНГ вплоть до образования фруктозо-1,6-бисфосфата протекают в цитозоле и катализируются гликолитическими ферментами. Затем следует еще одна необратимая реакция ГНГ, катализируемая фруктозо-1,6- бисфосфатазой, в которой происходит отщепление остатка фосфорной кислоты гидролитическим путем. Образующийся при этом фруктозо-6-фосфат гликолитическим ферментом изомеризуется до глюкозо-6-фосфата, от которого отщепляется остаток фосфорной кислоты при участии еще одного необратимо работающего фермента – глюкозо-6-фосфатазы. Образовавшаяся свободная глюкоза из клетки выходит в кровь. В ходе ГНГ расходуется 6 моль АТФ на синтез 1 моль глюкозы из пирувата или лактата.
Ключевыми ферментами ГНГ, катализирующими необратимые обходные реакции, являются: пируваткарбоксилаза, фосфоенолпируваткарбоксикиназа, фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза. Активируют ГНГ аллостерически АТФ и цитрат, обязательным активатором первого фермента является ацетил-КоА. ГНГ усиливают гормоны: глюкагон, глюкокортикостероиды. Тормозит ГНГ гормон покоя и сытости – инсулин.
Лактат, образовавшийся в ходе анаэробного гликолиза, не является конечным продуктом метаболизма. Использование лактата связано с его превращением в печени в пируват и с дальнейшим использованием последнего. Лактат, поступивший из интенсивно работающих мышц, выходит в кровь, а затем поступает в печень. В печени в лактатдегидрогеназной реакции лактат превращается в пируват, который включается в ГНГ, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами. Эту последовательность событий называют «глюкозо-лактатным циклом», или «циклом Кори». Цикл Кори выполняет две важнейшие функции: обеспечивает утилизацию лактата и предотвращает лактатацидоз. Часть пирувата,
45
образовавшегося из лактата (25%), окисляется печенью до СО2 и Н2О с освобождением энергии.
4.5. ПЕНТОЗОФОСФАТНЫЙ ПУТЬ ПРЕВРАЩЕНИЯ ГЛЮКОЗЫ
Функционирует в цитозоле клетки с пластической целью, то есть его главные продукты – рибозо-5-фосфат и НАДФН используются для синтезов. Пентозофосфатный путь (ПФП) может протекать в двух вариантах – окислительного и неокислительного.
Окислительный вариант включает 2 реакции дегидрирования, катализируемых НАДФ-зависимыми дегидрогеназами.
Суммарное уравнение окислительного варианта можно представить в таком виде:
Глюкозо-6-фосфат + 2 НАДФ+ Рибозо-5-фосфат + 2 НАДФН+ 2Н+ +СО2
Рибозо-5-фосфат + 2 НАДФН+ 2Н+ +СО2
Неокислительный вариант ПФП включает серию обратимых реакций, катализируемых транскетолазой и трансальдолазой, первая в качестве кофермента использует ТПФ. В качестве промежуточных соединений в этом варианте образуются углеводы, содержащие от 3 до 7 атомов углерода. В этом варианте нет реакций дегидрирования, поэтому он используется только для синтеза пентоз.
Суммарное уравнение реакций неокислительного варианта:
5 глюкозо-6-фосфат  6 рибозо-5-фосфат
6 рибозо-5-фосфат
Роль ПФП определяется его конечными продуктами: рибозо-5фосфатом и НАДФН. Рибозо-5-фосфат необходим для синтеза нуклеотидов, а затем и нуклеиновых кислот (ДНК и РНК). Быстрый синтез ДНК происходит в быстро делящихся тканях: красный костный мозг, эмбрион, базальный слой эпителия кожи, слизистая тонкого кишечника и др. Также рибозо-5-фосфат необходим для синтеза РНК и далее биосинтеза белка – в печени, экзокринных железах, синтезирующих белок на экспорт, а также усиленный синтез белка происходит при гипертрофии тканей. При этом у взрослого человека есть ткани, которые практически не делятся, а значит, в них ПФП неактивен. Но и в этих тканях ПФП может активироваться, например, при процессах репарации. Другой продукт ПФП – НАДФН необходим для реакций гидроксилирования и для восстановительных биосинтезов. Он активно используется для биосинтеза жирных кислот в жировой ткани, в печени; для синтеза холестерина (печень), для синтеза стероидных гормонов в эндокринных железах, для реакций гидроксилирования эндогенных веществ и ксенобиотиков. В эритроцитах НАДФН необходим для работы антиоксидативной системы, что обеспечивает их устойчивость к гемолизу.
4.6. ГОМЕОСТАЗ ГЛЮКОЗЫ КРОВИ
Уровень глюкозы крови поддерживается на постоянном уровне за счет того, что пути поступления глюкозы в кровь и пути расходования глюкозы крови между собой сбалансированы. При этом в течение суток уровень
46
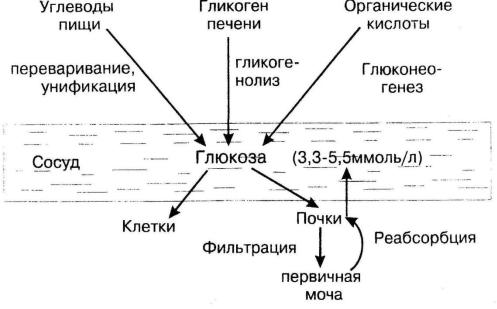
глюкозы не является одинаковым, но у здорового человека эти колебания лежат в пределах нормы (3,3 – 5,5 ммоль/л для капиллярной крови).
Источниками глюкозы крови являются: углеводы пищи, гликоген печени и органические кислоты, которые в результате глюконеогенеза превращаются в глюкозу. Расходуется глюкоза крови всеми клетками организма, при этом в одни клетки глюкоза поступает диффузно (например, головной мозг), а в другие – с помощью инсулина. Кроме того, глюкоза из крови в результате фильтрации попадает в первичную мочу, но затем в результате реабсорбции она возвращается в сосудистое русло (рис. 7).
Рис.7. Гомеостаз глюкозы крови
В регуляции уровня глюкозы крови принимают участие гормоны. Гормоны делятся на гипогликемические, т.е. снижающие уровень глюкозы крови, и гипергликемические, т.е. повышающие уровень глюкозы крови. Единственным гипогликемическим гормоном является инсулин. Гипергликемическими гормонами являются: глюкагон, катехоламины (адреналин и норадреналин), ГКС и соматотропный гормон (СТГ).
При гипергликемии (повышении глюкозы крови, которое может быть вызвано алиментарной причиной, сахарным диабетом, стрессом, некрозом поджелудочной железы, избытком гипергликемических гормонов) усиливается выработка инсулина, который за счет своих эффектов снижает уровень глюкозы крови. Это происходит за счет усиления потребления глюкозы тканями (в связи с увеличением проницаемости мембран для глюкозы в мышечной, жировой и соединительной тканях, усилением синтеза гликогена, гликолиза и пентозофосфатного пути), а также за счет торможения поступления глюкозы из тканей в кровь (в результате торможения распада гликогена и торможения глюконеогенеза).
При гипогликемии (снижении уровня глюкозы крови, которое может быть вызвано алиментарной причиной, патологией печени и ЖКТ, избытком
47
инсулина, при алкоголизме и др.) усиливается выработка гормонов гипергликемических, которые повышают уровень глюкозы крови. Это обусловлено усилением глюконеогенеза и распада гликогена, торможением синтеза гликогена и гликолиза. СТГ при этом действует опосредованно, путем усиления секреции глюкагона.
4.7. ТЕСТОВЫЕ ЗАДАНИЯ
Выбрать один правильный ответ
46.В ПЕРЕВАРИВАНИИ КРАХМАЛА УЧАСТВУЕТ ФЕРМЕНТ
1)амилаза
2)сахараза
3)лактаза
4)липаза
47.ИЗБЫТОК В РАЦИОНЕ САХАРОЗЫ МОЖЕТ УВЕЛИЧИТЬ РИСК
1)аллергических реакций
2)аутоиммунных заболеваний
3)ожирения
4)рака прямой кишки
48.БИОСИНТЕЗ ГЛИКОГЕНА УСИЛИВАЮТ ГОРМОНЫ
1)инсулин
2)глюкагон
3)катехоламины
4)глюкокортикостероиды
49.КОНЕЧНЫМ ПРОДУКТОМ АНАЭРОБНОГО ГЛИКОЛИЗА ЯВЛЯЕТСЯ
1)СО2 и Н2О
2)пируват
3)лактат
4)цитрат
50.НЕДОСТАТКОМ ГЛИКОЛИЗА ЯВЛЯЕТСЯ
1)малый энергетический выход
2)быстрота включения
3)единственный источник энергии для эритроцита
4)накопление пирувата
51.ГЛЮКОНЕОГЕНЕЗ ПРОТЕКАЕТ В
1)мышцах
2)головном мозге
3)эритроцитах
4)печени
48
52.В ГЛЮКОНЕОГЕНЕЗЕ ГЛЮКОЗА МОЖЕТ ОБРАЗОВАТЬСЯ ИЗ
1)пирувата
2)жирных кислот
3)холестерина
4)желчных кислот
53.ПРОДУКТАМИ ПЕНТОЗОФОСФАТНОГО ПУТИ ЯВЛЯЮТСЯ
1)НАДФН
2)гликоген
3)пируват
4)лактат
54.ПРИЧИНАМИ ГИПЕРГЛИКЕМИЙ МОГУТ БЫТЬ
1)сахарный диабет
2)патология печени
б) усиленная утилизация глюкозы
4)избыток инсулина
55.УРОВЕНЬ ГЛЮКОЗЫ КРОВИ ПОСЛЕ 48-ЧАСОВОГО ГОЛОДАНИЯ ПОДДЕРЖИВАЕТСЯ ЗА СЧЕТ
1)гликогена печени
2)гликогена мышц
3)глюконеогенеза
4)пентозофосфатного пути
56.КОНЕЧНЫМИ ПРОДУКТАМИ АЭРОБНОГО РАСПАДА ГЛЮКОЗЫ ЯВЛЯЮТСЯ
1)СО2 и Н2О
2)пируват
3)лактат
4)цитрат
57.УГЛЕВОДЫ ВЫПОЛНЯЮТ ФУНКЦИЮ
1)хранение и передача наследственной информации
2)энергетическую
3)каталитическую
4)транспортную
58.ГИПОГЛИКЕМИЧЕСКМИ ГОРМОНАМИ ЯВЛЯЮТСЯ
1)глюкагон
2)катехоламины
3)глюкокортикостероиды
4)инсулин
59.ПИЩЕВЫЕ ВОЛОКОНА
1)источник глюкозы для организма
49
2)активируют амилазу панкреатического сока
3)стимулируют перистальтику кишечника
60.ГАЛАКТОЗА В ЖЕЛУДОЧНО-КИШЕЧНОМ ТРАКТЕ ОБРАЗУЕТСЯ ПРИ ПЕРЕВАРИВАНИИ С УЧАСТИЕМ ФЕРМЕНТА
1)пепсин
2)-амилаза
3)мальтаза
4)сахараза
5)лактаза
5.ОБМЕН ЛИПИДОВ
5.1. ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ
Липиды обеспечивают 30% от калорийности рациона (при этом каждая группа жирных кислот – насыщенные, мононенасыщенные и полиненасыщенные – по 10%), что при суточной калорийности в 2000–3000 ккал составляет 70–100 г. Полиненасыщенные жирные кислоты (ПНЖК) не синтезируются в организме и обязательно должны поступать с пищей. Их источниками являются растительные масла и жиры морепродуктов. ПНЖК по месту расположения первой двойной связи, считая с гидрофобного конца цепи, делятся на две группы: -3 и -6. Подсолнечное, кукурузное и хлопковое масла являются источником -6 кислот (линолевой и арахидоновой). Соевое, конопляное и льняное масла содержат как -6, так и -3 кислоты (линоленовую и эйкозапентаеновую). Растительные масла содержат цис-ПНЖК, которые необходимы для синтеза фосфолипидов, обеспечивающих полноценность мембран, для образования эйкозаноидов и липидных вторых посредников. Кроме того, они полезны для профилактики атеросклероза и ИБС, необходимы для нормальности кожи, способствуют нормальному росту ребенка и развитию плода.
Переваривание жиров у взрослых происходит в тонком кишечнике при участии липазы поджелудочной железы. Липаза действует только на эмульгированный жир, поэтому перевариванию предшествует процесс эмульгации (дробление крупной липидной капли на мелкие), который осуществляется при участии желчных кислот, входящих в состав желчи. Жиры под действием липазы расщепляются на глицерин и жирные кислоты. Глицерин всасывается свободно, жирные кислоты образуют при участии желчных кислот мицеллы. Снаружи мицеллы находятся гидрофильные группы желчных кислот, а внутри – гидрофобные жирные кислоты. Мицелла путем пиноцитоза попадает в кишечную стенку, где распадается на жирные и желчные кислоты. Желчные кислоты с кровью снова приносятся в печень, откуда вместе с желчью опять попадают в кишечник. А из жирных кислот и глицерина в кишечной стенке происходит ресинтез жира. Из этого жира в кишечной стенке происходит
50