

Тема 3. Наследственность и изменчивость.
Наследственность и изменчивость являются одними из основных факторов эволюции живого на земле. Наследственность – это особое свойство организмов, заключающееся в способности обеспечивать материальную и функциональную преемственность в ряду поколений.
Человек появляется в результате слияния материнской и отцовской половых клеток (гамет) и содержащегося в них генетического материала. В ядрах половых клеток содержатся хромосомы, которые в процессе слияния гамет, образуют зиготу, т.е. оплодотворенную яйцеклетку, содержащую двойной набор хромосом.
В самих хромосомах в форме спирали, которую открыли Уотсон и Крик, содержится генетическая информация в виде стабильных структур, состоящих из нуклеиновых кислот, называемых генами. Как будет выглядеть человек, зависит от состава его генов. Повреждение генов может иметь необратимые последствия. Например, замена в белке всего одного аминокислотного остатка, контролирующего развитие эритроцитов, приводит к тому, что они навсегда приобретают серповидную форму и обуславливают анемию. Этот признак передается в семье рецессивным путем через поколение.
Открытие Г.Менделем законов, отражающих процесс передачи наследственной информации и принципа дискретности, явилось доказательством существования наследственности как материального явления.
В 1856—1863 гг. Мендель провёл обширные, тщательно спланированные опыты по гибридизации растений гороха. Для скрещиваний он отбирал константные сорта (чистые линии), каждый из которых при самоопылении устойчиво воспроизводил в поколениях одни и те же признаки. Сорта различались альтернативными (взаимоисключающими) вариантами какого-либо признака, контролируемого парой аллельных генов (аллелей). Например, окраской (жёлтая или зелёная), формой семян (гладкая или морщинистая), длиной стебля (длинный или короткий) и т.д. Для анализа результатов скрещиваний Мендель применил математические методы, что позволило ему обнаружить ряд закономерностей в распределении родительских признаков у потомков.
В 1866г. Г.Мендель опубликовал результаты своих опытов по скрещиванию различных линий гороха, отличавшихся друг от друга по некоторым «константно-различающимся» признакам, что позволило сформулировать законы передачи наследственных признаков:
.
Закон единообразия гибридов первого поколения, или первый закон Менделя, утверждает, что потомство первого поколения от скрещивания устойчивых форм, различающихся по одному признаку, имеет одинаковый фенотип по этому признаку. При этом все гибриды могут иметь фенотип одного из родителей (полное доминирование), как это имело место в опытах Менделя, или, как было обнаружено позднее, промежуточный фенотип (неполное доминирование). В дальнейшем выяснилось, что гибриды первого поколения могут проявить признаки обоих родителей (кодоминирование). Этот закон основан на том, что при скрещивании двух гомозиготных по разным аллелям форм (АА и аа) все их потомки одинаковы по генотипу (гетерозиготны — Аа), а значит, и по фенотипу.
Закон расщепления, или второй закон Менделя: при скрещивании гибридов первого поколения между собой среди гибридов второго поколения в определенных соотношениях появляются особи с фенотипами исходных родительских форм и гибридов первого поколения. Так, в случае полного доминирования выявляются 75% особей с доминантным и 25% с рецессивным признаком, т. е. два фенотипа в отношении 3:1 (рис. 1). При неполном доминировании и кодоминировании 50% гибридов второго поколения имеют фенотип гибридов первого поколения и по 25% — фенотипы исходных родительских форм, т. е. наблюдают расщепление 1:2:1. В основе второго закона лежит закономерное поведение пары гомологичных хромосом (с аллелями А и а), которое обеспечивает образование у гибридов первого поколения гамет двух типов, в результате чего среди гибридов второго поколения выявляются особи трёх возможных генотипов в соотношении 1АА:2Аа:1аа. Конкретные типы взаимодействия аллелей и дают расщепления по фенотипу в соответствии со вторым законом Менделя.
Закон независимого комбинирования (наследования) признаков, или третий закон Менделя: каждая пара альтернативных признаков ведёт себя в ряду поколений независимо друг от друга, в результате чего среди потомков второго поколения в определенном соотношении появляются особи с новыми (по отношению к родительским) комбинациями признаков. Например, при скрещивании исходных форм, различающихся по двум признакам, во втором поколении выявляются особи с четырьмя фенотипами в соотношении 9:3:3:1 (случай полного доминирования). При этом два фенотипа имеют «родительские» сочетания признаков, а оставшиеся два — новые. Этот закон основан на независимом поведении (расщеплении) нескольких пар гомологичных хромосом (рис. 2). Например, при дигибридном скрещивании это приводит к образованию у гибридов первого поколения 4 типов гамет (АВ, Ab, aB, ab) и после образования зигот — закономерному расщеплению по генотипу и соответственно по фенотипу.
Менделевские законы имели важное значение — именно на их основе происходило интенсивное развитие генетики на первом этапе. Они послужили основой для предположения о существовании в клетках (гаметах) наследственных факторов, контролирующих развитие признаков. Из них следует, что эти факторы (гены) относительно постоянны, хотя и могут находиться в различных состояниях, парны в соматических клетках и единичны в гаметах, дискретны и могут вести себя независимо по отношению друг к другу. Всё это послужило в своё время серьёзным аргументом против теорий «слитной» наследственности и было подтверждено экспериментально.
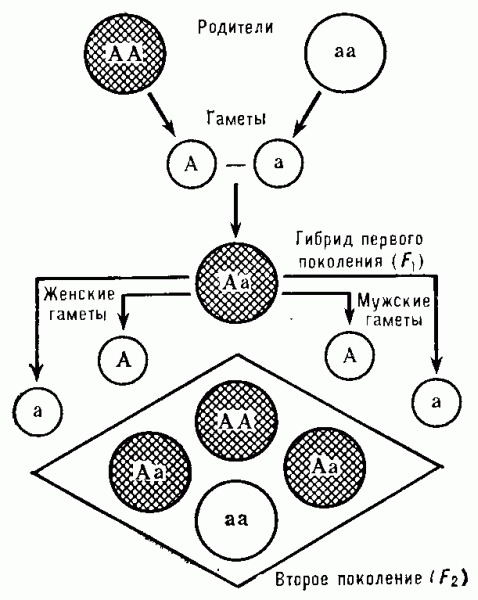
Рис. 3. Схема, иллюстрирующая единообразие гибридов первого поколения F1 (первый закон Менделя) и расщепление признаков у потомства второго поколения F2 с преобладанием доминантного фенотипа над рецессивным в отношении 3 : 1 (второй закон Менделя); A — доминантный ген, а — рецессивный ген. Заштрихованный круг — доминантный фенотип, а светлый — рецессивный.
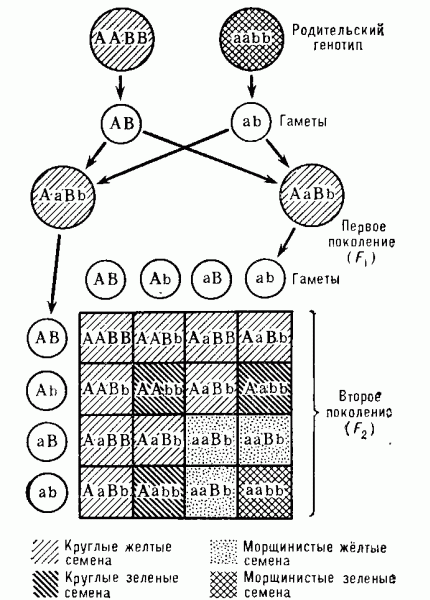
Рис. 4. Схема, иллюстрирующая независимое комбинирование признаков (третий закон Менделя). Наследование жёлтой (В) и зелёной (b) окраски семян, а также круглой (А) и морщинистой (а) их формы. А и В доминируют над аллелями а и b. Генотипы родителей и потомков обозначены комбинацией указанных букв, а четыре разных фенотипа — при помощи различной штриховки.
.(Источник: «Биологический энциклопедический словарь.» Гл. ред. М. С. Гиляров; Редкол.: А. А. Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. — 2-е изд., исправл. — М.: Сов. Энциклопедия, 1986.)
Традиционно в генетике принимают три закона Менделя, хотя сам он формулировал лишь закон независимого комбинирования. Часто как один из законов Менделя приводится и закон чистоты гамет, утверждающий, что в каждую половую клетку попадает только один аллельный ген. Однако, несмотря на фундаментальность этого закона (что подтверждают результаты тетрадного анализа), он не касается наследования признаков и, кроме того, сформулирован не Менделем, а У. Бэтсоном (в 1902).
Непонятый современниками, Мендель обнаружил дискретную («корпускулярную») природу наследственности и показал ошибочность представлений о «слитной» наследственности. После переоткрытия забытых законов основанное на экспериментах учение Менделя получило название менделизм. Его справедливость была подтверждена хромосомной теорией наследственности.
Поскольку у человека в геноме содержатся десятки тысяч генов, то простая комбинаторика генов (алеллей) порождает колоссальное разнообразие возможных сочетаний и представляет собой одну из сторон уникальности и неповторимости человека.
Следует отметить, что наследственные факторы являются лишь «потенциями развития», которые иногда могут быть нереализованными. Реализация генетических факторов происходит последовательно и зависит также от факторов среды. В пренатальном онтогенезе сначала действуют общие гены, которые определяют вид и пол ребенка. Затем начинают функционировать органоспецифичные гены, которые определяют развитие органов, уровень активности ферментов, метаболизм организма и фенотипические признаки: цвет глаз, волос и т.д.
Менделирующие признаки человека.
Менделирующими признаками называются те, наследование которых происходит по закономерностям, установленным Г.Менделем. менделирующие признаки определяются одним геном – моногенно (от греч. monos – один), т.е. когда проявление признака определяется взаимодействием аллельных генов, один из которых доминирует (подавляет) другой.
Если гены локализованы в половых хромосомах (за исключением гомологичного участка в Х- и Y-хромосомах), или в одной хромосоме сцеплено, или в ДНК органоидов, то результаты скрещивания не будут следовать законам Менделя.
Общие законы наследственности одинаковы для всех эукариот. У человека также имеются менделирующие признаки, и для него характерны все типы их наследования: аутосомно-доминантный, аутосомно-рецессивный, сцепленный с половыми хромосомами (с гомологичным участком Х- и Y-хромосом).
Типы наследования.
аутосомно-доминантный тип наследования. По этому типу наследуются некоторые нормальные и патологические признаки:
белый локон над лбом;
волосы жесткие, приямые (ежик);
шерстистые волосы – короткие, легко секущиеся, курчавые, пышные;
кода толстая;
способность свертывать язык в трубочку;
габсбургская губа – нижняя челюсть узкая, выступающая вперед, нижняя губа отвислая и полуоткрытый рот;
полидактилия ( от греч. polys – многочисленный, daktylos – палец) – многопалость, когда имеется от шести до девяти пальцев на руке или ноге;
синдактилия (от греч. syn – вместе) – сращение мягких или костных тканей фаланг двух и более пальцев;
брахидактилия (короткопалость) – недоразвитие дистальных фаланг пальцев;
арахнодактилия (от греч. arahna – паук) – сильно удлиненные «паучьи» пальцы.
Аутосомно-рецессивные менделирующие признаки у человека.
Если рецессивные гены локализованы в аутосомах, то проявиться они могут при браке двух гетерозигот или гомозигот по рецессивному аллелю.
По данному типу наследуются следующие признаки: волосы мягкие, прямые, кожа тонкая, 0 (I) группа крови, группа крови Rh-, неощущение горечи вкуса фенилкарбамида, неумение складывать язык в трубочку. По этому типу наследуются также многие болезни обмена веществ: фенилкетонурия, галактоземия, гистидинимия а пр.
Менделирующие признаки, сцепленные с полом (неполно).
X- и Y-хромосомы имеют общие гомологичные участки. В них локализованы гены, детерминирующие признаки, наследующиеся одинаково как у мужчин, так и у женщин (подобно признакам, сцепленным с аутосомами).
Гены, локализованные в гомологичных участках X- и Y-хромосом, обусловливают развитие некоторых болезней.
Пигментная ксеродерма – заболевание, при котором под влиянием УФ-лучей на открытых частях тела появляются пигментированные пятна. Вначале они в в идее веснушек, затем в виде более крупных папиллом различной величины и, наконец опухолей. Для большинства людей пигментная ксеродерма заканчивается летально.
Болезнь Огучи – в слое палочек и колбочек, пигментном эпителии наблюдаются дегенаритивные изменения (болезнь чаще встречается в Японии).
Спастическая параплегия – спастика и слабость нижних конечностей, возникающая в результате дегенерации пирамидных путей в области грудного и поясничного отделов спинного мозга, изредка в стволе головного мозга.
Эпидермолиз буллезный – образование пузырей после механических травм кожи.
Полная (общая) цветовая слепота – полное отсутствие цветового зрения.
Рассмотрим, как происходит наследование по группе крови. Распространенное по всему миру разделение людей на 4 группы крови основывается на системе АВ0. А и В – это антигены эритроцитов (агглютиногены). Если у человека они отсутствуют, то его кровь относится к первой группе (0). Если есть только А –ко второй, только В – к третьей, а если и А, и В – к четвертой. Точное определение принадлежности крови к конкретной группе возможно только в лабораторных условиях при помощи специальных сывороток.
По резус-фактору все население земного шара делится на его обладателей (резус-положительные) и тех, у кого этого фактора нет (резус-отрицательные). Отсутствие резуса никак не сказывается на здоровье. Однако у женщины имеется угроза резус-конфликта с ребенком, особенно при повторных беременностях, если в ее крови этот фактор отсутствует, а в крови малыша он есть.
Наследование групп крови и резус-фактора происходит по хорошо изученным законам генетики. От родителей ребенку передаются гены, несущие информацию о наличии или отсутствии агглютиногенов (А, В или 0), а также о наличии или отсутствии резус-фактора. Упрощенно генотипы людей разных групп крови записываются так:
Первая группа крови – 00. Один 0 («ноль») этот человек получил от матери, другой — от отца. Соответственно, и своему потомству человек с первой группой может передать только 0.
Вторая группа крови – АА или А0. Ребёнку от такого родителя может передаться А или 0.
Третья группа крови – ВВ или В0. Наследуется либо В, либо 0.
Четвертая группа крови – АВ. Наследуется либо А, либо В.
Что касается резус-фактора, то он наследуется, как доминантный признак. Это значит, что если он передается человеку хотя бы от одного из родителей, то обязательно проявится.
Если оба родителя отрицательны по резус-фактору, то все дети в их семье также не будут его иметь. Если один родитель имеет резус-фактор, а другой нет – у ребенка резус может присутствовать или отсутствовать. Если оба родителя резус-положительны, то минимум в 75% случаев ребенок также будет положительным. Однако появление в такой семье малыша с отрицательным резусом не нонсенс. Это вполне вероятно, если родители гетерозиготны – т.е. имеют гены, отвечающие как за наличие резус-фактора, так и за его отсутствие. На практике предположить это можно просто – расспросить кровных родственников. Вполне вероятно, что среди них отыщется резус-отрицательный человек.
Таблица вероятностей рождения ребенка с определенной группой крови при соответствующих данных по группам крови родителей:
|
I (00) |
II (A0) |
II (AA) |
III (B0) |
III (BB) |
IV (AB) | |
|
I (00) |
I (00) - 100% |
I (00) - 50% II (A0) - 50% |
II (A0) - 100% |
I (00) - 50% III (B0) - 50% |
III (B0) - 100% |
II (A0) - 50% III (B0) - 50% |
|
II (A0) |
I (00) - 50% II (A0) - 50% |
I (00) - 25% II (A0) - 50% II (AA) - 25% |
II (A0) - 50% II (AA) - 50% |
I (00) - 25% II (A0) - 25% III (B0) - 25% IV (AB) - 25% |
III (B0) - 50% IV (AB) - 50% |
II (A0) - 25% II (AA) - 25% III (B0) - 25% IV (AB) - 25% |
|
II (AA) |
II (A0) - 100% |
II (A0) - 50% II (AA) - 50% |
II (AA) - 100% |
II (A0) - 50% IV (AB) - 50% |
IV (AB) - 100% |
II (AA) - 25% III (B0) - 25% |
|
III (B0) |
I (00) - 50% III (B0) - 50% |
I (00) - 25% II (A0) - 25% III (B0) - 25% IV (AB) - 25% |
II (A0) - 50% IV (AB) - 50% |
I (00) - 25% III (B0) - 50% III (BB) - 25% |
III (B0) - 50% III (BB) - 50% |
II (A0) - 25% III (B0) - 25% III (BB) - 25% IV (AB) - 25% |
|
III (BB) |
III (B0) - 100% |
III (B0) - 50% IV (AB) - 50% |
IV (AB) - 100% |
III (B0) - 50% III (BB) - 50% |
III (BB) - 100% |
III (BB) - 50% IV (AB) - 50% |
|
IV (AB) |
II (A0) - 50% III (B0) - 50% |
II (A0) - 25% II (AA) - 25% III (B0) - 25% IV (AB) - 25% |
II (AA) - 25% III (B0) - 25% |
II (A0) - 25% III (B0) - 25% III (BB) - 25% IV (AB) - 25% |
III (BB) - 50% IV (AB) - 50% |
II (AA) - 25% III (BB) - 25% IV (AB) - 50% |
Стоит помнить, что группа крови, рассчитанная с помощью схем, таблиц или калькуляторов не может считаться окончательной. Точно узнать группу крови своего малыша вы сможете только по результатам лабораторных анализов.
Более тонкую работу генетического контроля можно проследить на примере подбора группы крови для переливания. В зависимости от состояния антигенов (содержатся в эритроцитах) и антител (локализуются в плазме) человек относится к одной из четырех групп, обозначаемых 0 (I), А (II), В (III), и АВ (IV), см. таблицу 1.
Таблица 1
|
Группа крови |
0 (I) |
А (II) |
В (II) |
АВ (IV) |
|
Антиген А |
- |
+ |
- |
+ |
|
Антиген В |
- |
- |
+ |
+ |
|
Антитела α |
+ |
- |
+ |
- |
|
Антитела β |
+ |
+ |
- |
- |
Антитела α вызывают агглютинацию (слипание и разрушение) эритроцитов, несущих антиген А, антитела β – агглютинацию эритроцитов, несущих антиген В. Т.о., в крови человека не должно содержаться одновременно антигенов и антител типа А, иначе его кровь свернется. Аналогичным образом не могут совмещаться антигены В и антитела β. Если же кровь группы А переливают человеку, кровь которого относится к группе В, то агглютинация произойдет главным образом потому, что кровь группы В содержит антитела α, и в меньшей степени за счет антител β, вводимых с переливаемой кровью, т.е. 1) антигены содержатся в кровяных тельцах (эритроцитах), а антитела – в плазме; 2) переливаемые эритроциты не должны агглютинироваться плазмой реципиента.
Люди с группой крови 0(I) особенно ценны в качестве доноров, так как ни одна сыворотка не вызывает агглютинацию их эритроцитов. Однако эти люди – плохие реципиенты, так как их сыворотка агглютинирует эритроциты всех остальных типов: А, В и АВ. Таким образом, людям, принадлежащим по крови к группе 0, можно переливать только кровь людей, относящихся к этой же группе, иначе они погибнут.
Люди с группой крови АВ представляют собой противоположную крайность: им можно переливать кровь любой группы, не опасаясь вредных последствий, но они плохие доноры, поскольку их эритроциты агглютинируются сывороткой крови групп А, В и 0.
Считается, что гены, от которых зависит группа крови, оказывают плейотропное действие на различные органы и скорее отражают приспособленность к разным условиям внешней и генотипической среды. Основанием для подобного вывода служит то обстоятельство, что люди с некоторыми группами крови более других подвержены определенным заболеваниям. Например, люди с группой крови А склонны к заболеванию раком желудка, тогда как принадлежащие к группе 0 чаще других заболевают язвой двенадцатиперстной кишки. Различия в группах крови связаны также с различиями в жизнеспособности, причем характер этого влияния у мужчин и женщин неодинаков. Сходное влияние оказывают различия в группах крови и на способность к деторождению.
Знание того, к какой группе крови системы АВ0 мы принадлежим, имеет первостепенное значение при переливании крови, а также при определении родства и в судебно-медицинской практике. Большей частью люди относятся либо к группе резус-отрицательных, либо к группе резус-положительных. Генетическая основа групп крови системы резус сложна; известно не менее 12 различных аллелей для резус-признаков.
В системе АВ0 антитела возникают спонтанно, естественным путем, а резус-антитела – лишь в результате иммунизации.
С несовместимостью групп крови матери и ребенка связана так называемая гемолитическая болезнь новорожденных, которая проявляется гемолизом (разрушением) эритроцитов плода или ребенка в ответ на агрессию со стороны антител матери. Несовместимость крови матери и плода, обусловленная резус-конфликтом может приводить к тяжелым энцефалопатиям или смерти.
Действие патологических генов имеет свои сроки реализации и точки приложения. Дебют наследственных болезней происходит в разное время – от периода новорожденности до старости. В настоящее время произведено картирование 1000 патологических генов, составлены генетические карты с локализацией гена в определенном участке хромосомы. Многие гены клонированы, т.е. расшифрована их белковая структура и определен механизм их действия.
ИЗМЕНЧИВОСТЬ.
Если наследственность обеспечивает сходство морфологических характеристик, механизмов развития и жизнедеятельности, то структурно-функциональное разнообразие особей каждого вида зависит от изменчивости, то есть изменчивостью можно назвать фундаментальное свойство живого приобретать новые признаки в ходе онтогенеза.
Изменчивость выражается в бесконечном разнообразии признаков и свойств у особей различной степени родства. При классификации типов и форм изменчивости подчеркиваются те или иные стороны этого универсального свойства. Учитывая причины и характер изменений, обычно выделяют два основных типа изменчивости. Наследственная, или генотипическая, изменчивость обусловлена изменениями в генетическом материале (генотипе), которые передаются из поколения в поколение. Изменения в генотипе могут быть вызваны мутациями – изменениями в структуре генов и хромосом или изменениями числа хромосом в хромосомном наборе. При мутационной изменчивости возникают новые варианты (аллели) генов, причём мутации происходят сравнительно редко и внезапно. Другая форма генотипической изменчивости – т.н. комбинативная изменчивость, в основе которой лежит перекомбинация (перегруппировка) хромосом и их участков при половом размножении (в процессе мейоза и оплодотворения). В результате набор генов, а следовательно, и признаков у потомков всегда отличается от набора генов и признаков у родителей. Комбинативная изменчивость создаёт новые сочетания генов и обеспечивает как всё разнообразие организмов, так и неповторимую генетическую индивидуальность каждого из них.
Ненаследственная, или модификационная, изменчивость – способность организмов изменяться под действием различных факторов окружающей среды (температуры, влажности и т.п.). Этот тип изменчивости не связан с изменениями в генотипе и не наследуется. Однако пределы модификационной изменчивости любого признака – т.н. норма реакции – задаются генотипом. Степень варьирования признака, т.е. широта нормы реакции, зависит от значения признака: чем важнее признак, тем у́же норма реакции. Модификационная изменчивость носит групповой характер – изменения (модификации) возникают у всех особей популяции, которая подвергается влиянию определённого внешнего воздействия. Другая её особенность – обратимость: обычно модификации сразу или постепенно исчезают при устранении вызвавшего их фактора. Ч. Дарвин различал определённую (или групповую) и неопределённую (или индивидуальную) изменчивость, что по современной классификации совпадает соответственно с ненаследственной и наследственной изменчивостью. Следует помнить, однако, что это разделение в известной степени условно, т.к. пределы ненаследственной изменчивости определяются генотипом.
Наряду с наследственностью изменчивость – фундаментальное свойство всех живых существ, один из факторов эволюции органического мира. Различные способы целенаправленного использования изменчивости (разные типы скрещиваний, искусственные мутации и др.) лежат в основе создания новых пород домашних животных и сортов культурных растений.
(Источник: «Биология. Современная иллюстрированная энциклопедия.» Гл. ред. А. П. Горкин; М.: Росмэн, 2006.)
Взаимодействие неаллельных генов (гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки) обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов.
Выделяют три формы взаимодействия неаллельных генов:
комплементарность;
эпистаз;
полимерия.
Комплементарное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9:6:1, 9:3:4, 9:7, иногда 9:3:3:1.
Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлинённую. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдёт расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлинённые.
Подавление (ингибирование) действия одной аллельной пары генов геном другой, не аллельной им пары, называется эпистазом. Различают доминантный и рецессивный эпистаз. Если обычное аллельное доминирование можно представить в виде формулы А>а, То явление эпистаза выразится формулой А>В (доминантный эпистаз) или А>В (рецессивный эпистаз), когда доминантный или рецессивный ген одной аллельной пары не допускает проявления генов другой аллельной пары.
Гены, подавляющие действие других, не аллельных им генов, называются Эпистатичными, а подавляемые — Гипостатичными. Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию. При эпистазе фермент, образующийся под контролем одного гена, полностью подавляет или нейтрализует действие фермента, контролируемого другим геном.
Разберем эпистатическое действие генов на примере наследования окраски зерна у овса (рис. 28). У этой культуры были установлены доминантные гены, определяющие черную и серую окраску зерна. Обозначим один из них буквой А, А второй — В. При этом можно представить себе скрещивание, в котором родительские формы имели генотипы A Abb (черносемянный) и АаВВ (серосемянный). В генотипе растения первого поколения (АаВB) Содержатся доминантные гены и черной окраски А, И серой окраски В. Так как ген А Эпистатичен по отношению к гену В, Он не дает ему проявиться, и все гибриды F1 Будут черносемянными. В F1 Произойдет расщепление в отношении 12 черных : 3 серых: 1 белый. Такой результат расщепления легко понять, если представить себе отношение 12:3:1 как видоизменение типичного для дигибридных скрещиваний отношения 9:3:3:1.
В девяти сочетаниях присутствуют оба доминантных гена А И В, Но ген серой окраски В Не может проявляться, и они дают черносемянные растения. В трех сочетаниях (AAbbAabb, Aabb) Ген черной окраски семян А Также обусловит развитие черносемянных растений. Эта группа по фенотипу будет совершенно сходна с первой, и, следовательно, из каждых 16 растений 12 будут черносемянными. В трех сочетаниях (ааВВ, ааВB, ааВB) Доминантный ген В При отсутствии эпистатичного гена А Может проявить доминантное действие по отношению к своему рецессивному аллелю b, И разовьются растения с серыми семенами. Один генотип (Aabb) Представляет собой новую комбинацию, в которой проявится белая окраска зерна, так как отсутствуют оба доминантных гена.
Полимерия — взаимодействие неаллельных множественных генов, однонаправленно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммарного действия нескольких генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление в F2 по фенотипу при дигибридном скрещивании происходит в соотношении 1:4:6:4:1, а в целом соответствует третьей, пятой (при дигибридном скрещивании), седьмой (при тригибридном скрещивании) и т.п. строчкам в треугольнике Паскаля.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление в F2 по фенотипу при дигибридном скрещивании — 15:1.
Пример полимерии — наследование цвета кожи у людей, который зависит (в первом приближении) от четырёх генов с кумулятивным эффектом.
Хромосомная теория наследственности Т.Моргана.
Установленные Г.Менделем принципы независимого наследования и комбинирования признаков проявляются только тогда, когда гены, определяющие эти признаки, находятся в разных хромосомах (относятся к разным группа м сцепления).
Т.Морганом и его группой (1910-1916 гг.) была сформулирована хромосомная теория наследственности. Они подтвердили, что гены находятся в хромосомах. Гены, локализованные в одной хромосоме, образуют группу сцепления и передаются потомству совместно. Групп сцепления столько, сколько пар хромосом, или число групп сцепления равно числу хромосом в гаплоидном наборе. У женщин 23 группы сцепления, т.к. у них 23 пары гомологичных хромосом, а у мужчин 24 группы сцепления, т.к. у них 22 пары аутосом, X-, Y-хромосомы – две отдельные группы сцепления.
Следует подчеркнуть, что гены, локализованные в одной хромосоме, сцеплены не абсолютно. В профазе мейоза гомологичные хромосомы конъюгируются (сближаются), происходит кроссинговер, в результате которого несестринские хроматиды обмениваются гомологичными (одинаковыми) участками. Кроссинговер может происходить в разных участках хромосомы. Чем дальше друг от друга расположены локусы в одной хромосоме, тем выше вероятность кроссинговера между ними. Результаты опытов над мухами дрозофилы показали, что если, кроссинговер практически не происходит, то гены, расположенные в одной хромосоме, обнаруживают полное сцепление, т.е. наследуются совместно. Если кроссинговер происходит, наблюдают неполное сцепление.
В результате анализа опытов Т. Морган сформулировал закон: гены в хромосоме расположены линейно и наследуются сцеплено. Сила сцепления зависит от расстояния между генами, которое измеряется в процентах кроссинговера: 1% кроссинговера равен 1 морганиде. Процент кроссинговера для разных пар генов колеблется от долей единицы до пятидесяти. Уже при расстоянии в 50 морганид признаки наследуются практически независимо, несмотря на то, что гены локализуются в одной хромосоме.
Частота кроссинговера может сильно увеличиваться под влиянием как внутренних (видовая принадлежность, возраст, гормональный фон и др.), так и внешних факторов (температура, радиация, химические мутагены, некоторые лекарственные препараты и др.).
Основные положения хромосомной теории наследственности.
Гены располагаются в хромосомах, различные хромосомы содержат неодинаковое число генов, набор генов каждой из негомологичных хромосом уникален.
Гены в хромосоме расположены линейно, каждый ген занимает в хромосоме определенный локус (место).
Гены, расположенные в одной хромосоме, образуют группу сцепления и вместе (сцеплено) передаются потомкам, число групп сцепления равно гаплоидному набору хромосом.
Сцепление не абсолютно, т.к. в профазе мейоза может происходить кроссинговер и гены, находящиеся в одной хромосоме разобщатся. Сила сцепления зависит от расстояния между генами в хромосоме: чем больше расстояние, тем меньше сила сцепления, и наоборот. Расстояние между генами измеряется в процентах кроссинговера. 1% кроссинговера соответствует одной морганиде.
Влияние генотипической среды и факторов внешней среды на проявление признаков.
Признаки проявляются под влиянием генотипической среды и условий внешней среды. При наличии гена, детерминирующего признак, степень выраженности его различна. Это называется экспрессивностью.
Экспрессивность – степень выраженности признака.
Пенетрантность – это частота фенотипического проявления гена среди носителей этого гена. Если среди ста носителей гена признак проявляется у всех – это 100 % пенетрантность. Если среди ста носителей гена признак проявляется только у некоторых – это неполная пенетрантность. Пенетрантность может отсутствовать, т.е. признак не проявляется.
Знание пенетрантности имеет значение для определения генотипа людей родственники которых имеют наследственные заболевания.