


Приобретенный (специфический) иммунитет 173
один нуклеотид в одном цикле деления, что в 106 раз больше, чем частота мутаций в обычных клеточных генах.
Итак, разнообразие антител объясняется следующими механизмами:
1.Комбинаторное соединение сегментов генов;
2.Связывание цепей: после стандартной V(D)J-рекомбинации неиспользуемые сегменты удаляются, это приводит к появлению двунитевых разрывов между сегментами. Эти разрывы должны быть закрыты. Часто это требует удаления или вставки нуклеотидов, что и формирует многообразие;
3.Комбинаторное соединение тяжелой и лёгкой цепей;
4.Соматическая гипермутация в V-областях.
ДПервичный иммунный ответ
Первичный иммунный ответ заключается в том, что недифференцированные B-лимфоциты («наивные» B-клетки), встречаясь с антигеном, активируются и проходят стадии деления и дифференцировки в плазматические клетки и B-клетки памяти. Этот процесс ещё называют лаг-фазой (когда антитела ещё не продуцируются); продлиться он может от нескольких дней до месяцев. Затем плазматические клетки продуцируют антитела IgM (и небольшое количество IgG), которые способствуют эффективному уничтожению антигена в организме. B-клетки памяти сохраняются в крови и участвуют во вторичном иммунном ответе. В целом, от момента встречи B- лимфоцита с антигеном до продукции достаточного количества антител плазматическими клетками проходит 3-14 дней. При этом у пациента развиваются симптомы заболевания, поскольку антигену удаётся вызвать повреждение тканей.
Е Вторичный иммунный ответ
Вторичный иммунный ответ обеспечивается B-клетками памяти, когда антиген, вызвавший первичный иммунный ответ, снова попадает в организм. Клетки памяти начинают быстро делиться и формировать плазматические клетки, а те, в свою очередь, продуцируют антитела IgG (и небольшое количество IgM) к данному антигену, и его удаётся быстро уничтожить. Обычно вторичный иммунный ответ занимает от нескольких часов до нескольких дней и протекает гораздо быстрее первичного.
Ж Активация гуморального иммунитета
Теперь рассмотрим подробно процессы, происходящие в рамках гуморального иммунитета. Итак, все антигены можно разделить на:
—Тимус-зависимые антигены (требуют участия Т-хелперов): большинство антигенов;
—Тимус-независимые антигены (не нуждаются в Т-хелперах): бактериальные полисахариды, липополисахариды, высокополимерные белки.
173

174 |
Глава 9 |
Иммунитет и антитела |
Соответственно, выделяют два пути активации B-лимфоцитов: тимус-зависи- мый и тимус-независимый путь. Оба пути требуют передачи двух сигналов на B- лимфоцит:
—Тимус-зависимый: сигнал 1 (от взаимодействия антигена с мембранными антителами B-лимфоцитов IgM и IgD) и сигнал 2 (от взаимодействия CD40-белка B-лимфоцита и CD-40L-лиганда Т-хелпера).
—Тимус-независимый: сигнал 1 (от взаимодействия антигена с мембранными антителами B-лимфоцитов IgM и IgD) и сигнал 2 (от взаимо-
действия Толл-подобного рецептора B-лимфоцита и другого фраг-
мента антигена, например, липополисахаридов бактерий или бактериальной ДНК).
Рассмотрим подробно тимус-зависимый путь активации B-лимфоцитов (см. Рис. 106 ▼).
1.Представление антигена:
—Антиген, попадая в организм, «узнается» и поглощается антиген-
представляющими клетками (АПК) — B-лимфоцитами, макро-
фагами и дендритными клетками. В узнавании антигенов участ-
вуют Толл-подобные рецепторы и B-клеточные рецепторы (BCR,
только у B-лимфоцитов).
В нашем случае антиген взаимодействует с мембранными иммуноглобулинами IgM и IgD B-лимфоцита — так передаётся сиг-
нал 1.
—Внутри B-лимфоцита (или другой АПК) вакуоль с патогеном сливается с лизосомами и подвергается перевариванию, а фрагменты белков патогена «выставляются» на поверхности этих клеток в комплексе с белками MHC класса II (англ. MHC, major histocompatibility complex — главный комплекс гистосовместимости). Это явление получило название «представления» антигена.
—B-лимфоцит (или другая АПК) мигрирует в лимфатические узлы. С ним встречаются «наивные» (недифференцированные) Т-хел- перы. На поверхности Т-хелперов находится комплекс Т-клеточ- ного рецептора (TCR) и CD3-маркера, который взаимодействует с комплексом фрагмента антигена и белков MHC класса II на поверхности АПК.
—Взаимодействие этих двух комплексов запускает сложный каскад внутриклеточных сигнальных путей. Сближение АПК и Т-хелпера обеспечивается специфическими белками: интегринами LFA-1 (Т- клетки) и ICAM (АПК).
2.Проверка антигена:
—После передачи сигнала на Т-хелпер наступает следующая фаза представления антигена — проверка. Т-хелпер должен быть «уверен» в том, что ему был представлен именно чужеродный антиген. Это обеспечивается взаимодействием особых белков CD28 (на поверхности Т-хелпера) и СD80/CD86 (на поверхности АПК).
3.Пролиферация:

Приобретенный (специфический) иммунитет 175
—Как только вторая стадия пройдена, Т-хелперы секретируют ин- терлейкин-2 (и рецептор к нему — CD25 или интерлейкин-2R) во внеклеточное пространство, который сразу же действует на секретировавшую его клетку (аутокринный механизм). Кроме того, ин- терлейкин-2 воздействует на соседние клетки Т-хелперов, вызывая их пролиферацию.
—Клетки, активированные таким образом, становятся Т-хел-
перами 0 и секретируют интерлейкин-2, интерлейкин-4 и интер-
ферон γ (ИФН-гамма).
—Затем Т-хелперы 0 дифференцируются в Т-хелперы 1 или Т-хел- перы 2 в зависимости от цитокинов, которые воздействуют на них.
—Интерферон γ стимулирует продукцию Т-хелперов 1, а интерлей- кин-4 и интерлейкин-10 ингибируют её.
—Интерлейкин-4 стимулирует продукцию Т-хелперов 2, а интерферон γ ингибирует её.
4.Взросление:
—Пройдя много циклов деления, предшественники Т-хелперов дифференцируются в эффекторные Т-хелперы, Т-хелперы памяти и регуляторные Т-хелперы.
175
176 |
Глава 9 |
Иммунитет и антитела |
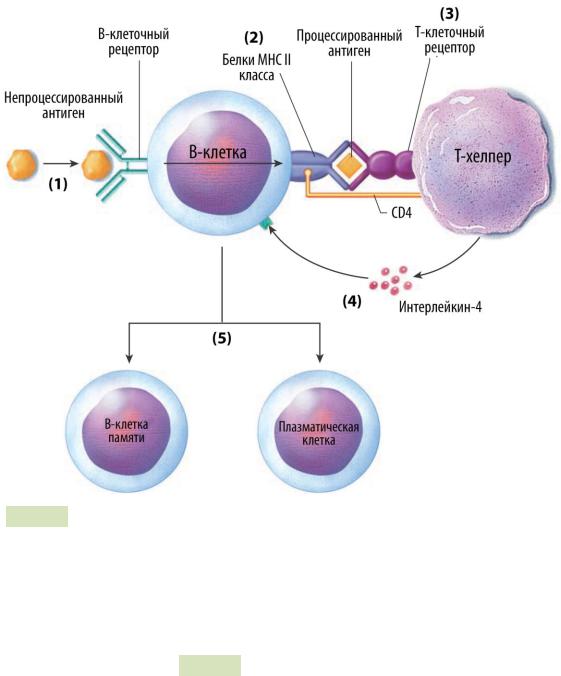
Рис. 106. Активация и пролиферация B-клеток. 1) Контакт с антигеном. 2 и 3) Взаи-
модействие B-клетки с T-хелпером. 4) Выделение цитокинов (в т.ч. ИЛ-2 Т-хелпером).
5)Пролиферация В-клеток.
5.Активация B-клеток:
—Т-хелперы взаимодействуют с B-лимфоцитами с помощью своих мембранных белков: CD40 (у B-лимфоцита) и CD-40L (у Т-хел- пера) (см. Рис. 106 ▲).
—В активации B-лимфоцитов участвует интерлейкин-4 — так передаётся сигнал 2.
—После того, как оба сигнала переданы, B-лимфоциты пролифери-
руют и дифференцируются в плазматические клетки и B-клетки памяти. Это происходит в герминативных центрах вторичных лимфоидных органов (лимфоузлы и селезёнка). Кроме того, происходит переключение изотипа антител B-лимфоцитов на IgG,
IgA и IgE.
—Плазматические клетки начинают синтезировать до 2000 молекул антител в секунду. Антитела обеспечивают:
Нейтрализацию — связывание антигена, препятствующее его вредной активности в организме;

Приобретенный (специфический) иммунитет 177
Опсонизацию — «помечают» антигены для фагоцитирования;
Активацию комплемента по классическому пути (см.
Тему 9.8).
—B-клетки памяти продолжают циркулировать в кровяном русле. Участвуют во вторичном иммунном ответе и способны быстро реагировать на повторное появление антигена в организме.
ЗАктивация клеточного иммунитета
Клеточный иммунитет обеспечивается цитотоксическими Т-клетками (Т- киллерами) и служит для уничтожения патогенов, находящихся внутри клеток организма: всех вирусов, некоторых бактерий, грибов и паразитов. Кроме того, клеточный иммунитет участвует в ряде аллергических реакций, в борьбе с опухолями и отторжении трансплантированных клеток.
После попадания в клетку вирус запускает процесс собственной репликации. Затем новые вирусные частицы покидают клетку и заражают другие. Находясь внутри клетки, вирусам удаётся избежать гуморального иммунитета и действия антител. Чтобы остановить репликацию вируса, необходимо уничтожить клетку, в которую ему удалось проникнуть. На первой стадии заражения инфицированные клетки обнаруживаются и уничтожаются естественными киллерами (NK-клетками). Тем временем происходит активация и деление Т-киллеров, которые способны с бо́льшей эффективностью уничтожать инфицированные клетки.
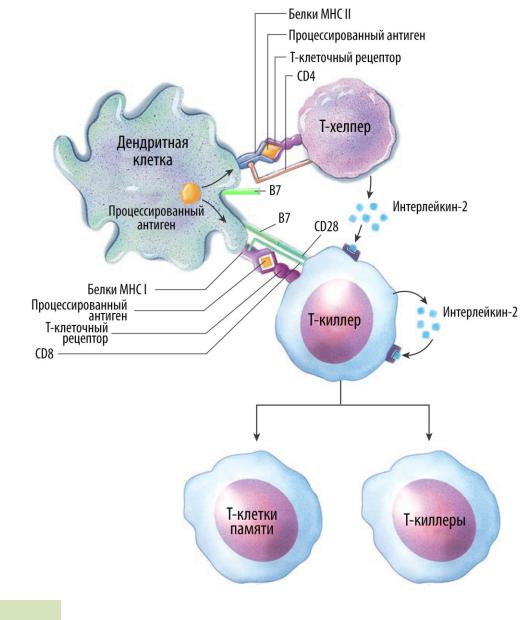
1.Дендритные клетки, самые эффективные АПК, поглощают вирусную частицу или становятся её «жертвой» (см. Рис. 107 ▼).
2.Происходит переваривание вирусной частицы (антигена) и «презентация» фрагмента антигена на поверхности дендритной клетки в комплексе с белками MHC класса I. С этим комплексом взаимодействует Т- киллер с помощью своего Т-клеточного рецептора и белка CD8. Помимо этого, белок B7 дендритной клетки взаимодействует с белком CD28 Т-киллера.
3.Почти одновременно с этим комплекс фрагмента антигена и белков MHC класса II на поверхности дендритной клетки взаимодействует с Т-
клеточным рецептором и белком CD4 Т-хелпера.
4.Т-хелпер начинает секретировать интерлейкин-2, интерлейкин-10, γ - интерферон (запускает свою же продукцию и секрецию другими Т-хел-
перами) и трансформирующий фактор роста-β. Интерлейкин-2 по
паракринному механизму воздействует на Т-киллер. Сам Т-киллер тоже выделяет интерлейкин-2, который сразу же воздействует на саму выде-
лившую его клетку (аутокринный механизм).
5.Т-киллеры проходят много циклов клеточного деления и дифференци-
руются в зрелые Т-киллеры и Т-клетки памяти (участвуют во вторич-
ном иммунном ответе).
6.Почти все клетки организма экспрессируют на своей поверхности белки MHC класса I. Если вирусу удалось проникнуть в клетку, то фрагменты его белков будут «выставлены» на поверхности клетки в комплексе с
177
178 |
Глава 9 |
Иммунитет и антитела |
белками MHC класса I. Зрелый Т-киллер взаимодействует с этим комплексом с помощью Т-клеточного рецептора и белка CD8. У здоровых клеток организма в мембране нет антигенов в комплексе с белками MHC класса I, поэтому Т-киллеры на них не реагируют, однако стоит фрагменту антигена появиться на мембране клетки, как Т-киллер активируется и секретирует перфорины и гранзимы.
а. Перфорин, по структуре напоминающий белок комплемента C9, образует в мембране клетки литическую пору.
б. Гранзимы проникают в клетку через образованные каналы и запускают апоптоз.
Если вириону удаётся «выйти» из клетки через поры до апоптоза, он становится мишенью фагоцитарных клеток и гуморального иммунитета (встречается с антителами, специфичными к нему, если соответствующие специфичные к нему популяции плазматических клеток и B-клеток памяти уже существуют).
Рис. 107. Активация и пролиферация T-клеток.