

Вторичная структура ДНК 75
3.4 Вторичная структура ДНК
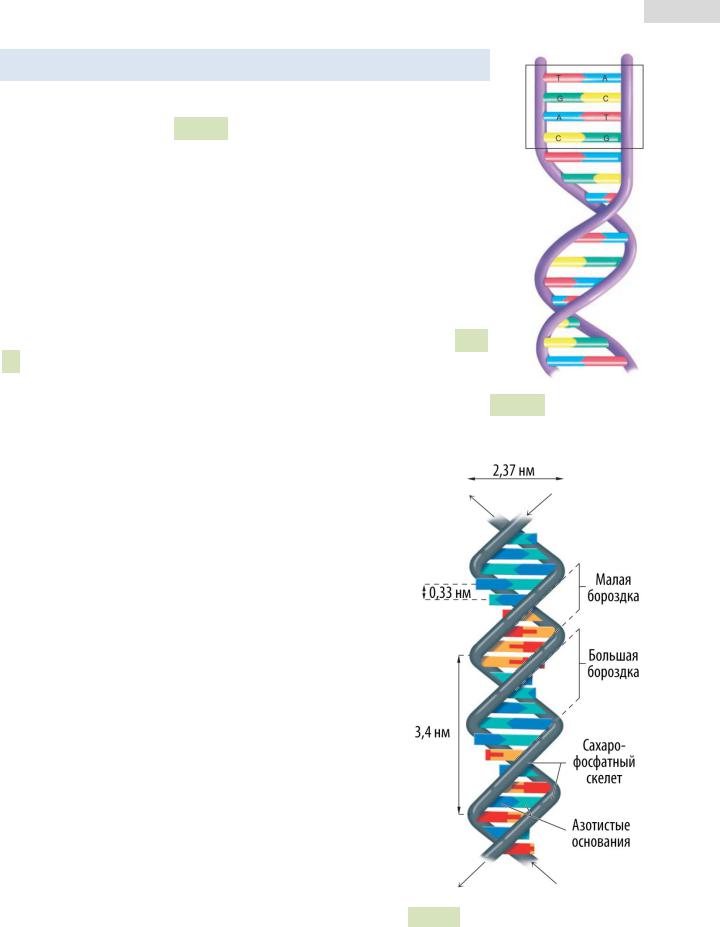
Вторичная структура ДНК представляет собой двухцепочечную спираль (см. Рис. 50 ), из двух длинных полинуклеотидных цепей, закрученных относительно общей оси. Сахарофосфатный скелет обеих цепей обращён наружу от оси спирали, а азотистые основания обращены внутрь спирали.
Две цепи удерживаются вместе с помощью водородных связей между азотистыми основаниями (аденин образует две связи с тимином, а гуанин — три связи с цитозином), гидрофобных связей, стэкинг-взаимодействий и ионных связей. Одна цепь ДНК комплементарна другой, при этом цепи антипараллельны.
Структурные особенности двойной спирали ДНК (см. Рис.
51 ):
1.Две антипараллельные цепи являются правозакрученными вдоль общей оси, диаметр спирали
~23,7 Å (2,37 нм).
2.Плоскости, в которых лежат азотистые основания, образующие водородные связи, почти перпендикулярны оси спирали. В B-ДНК основания занимают центральную часть спирали, а сахарофосфатный скелет закручен и обращён наружу. Только края оснований контактируют с растворителем.
3.Каждое основание имеет приблизительно одну ширину, что придает спирали ДНК практически полную симметрию независимо от состава самих оснований. Основания в парах можно поменять местами без каких-либо последствий для спирали: Г ≡ Ц можно изменить на Ц ≡ Г или А = Т на Т = А.
Рис. 50. Вторичная структура ДНК.
Рис. 51. Вторичная структура ДНК: двойная спираль и её параметры.
75
76 |
Глава 3 |
Нуклеиновые кислоты |
4.Каноническая (идеальная) форма B- ДНК имеет 10 пар оснований в одном шаге (в одном повороте спирале). При этом на каждую пару приходится 36° (если быть точным, то число пар в шаге — 10,4, следовательно, угол составляет 34,6°). Пары оснований уло-
жены в спирали «стопками». Шаг спирали составляет 34 Å (3,4 нм).
ДНК обладает структурной гибкостью и существует в нескольких формах: A-ДНК, B-ДНК (классическая Уотсон-Криковская двойная спираль), Z-ДНК и др. A- и B-ДНК являются правозакрученными спиралями. В B-ДНК азотистые основания располагаются почти перпендикулярно к оси вращения спирали. Z-ДНК представляет собой левозакрученную спираль. Бо́льшая часть ДНК клетки находится в B-форме (см. Приложение 3).
В двойной спирали различают большую и малую бороздки — это расстояния между сахарофосфатными остовами обеих цепей в двойной спирали.
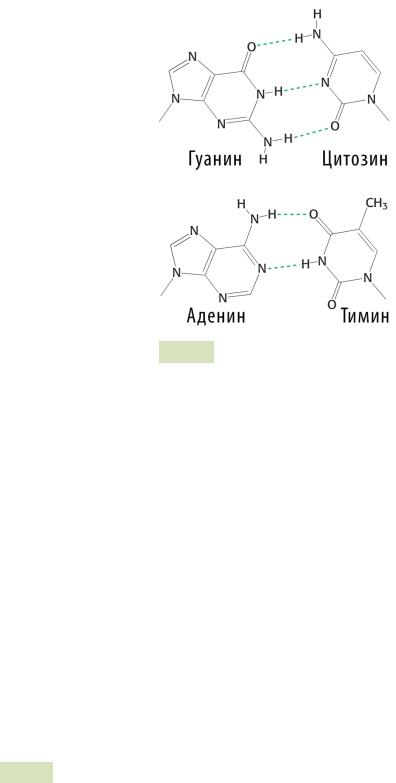
Рис. 52. Уотсон-Криковские пары комплементарных осно-
ваний: пунктиром отмечены водородные связи между атомами
H, O и N.
Длину ДНК измеряют в килоба́зах, т.е. тысячах пар нуклеотидов: 1 кб = 1 000 пар нуклеотидов. Геном одной клетки человека (т.е. все 46 хромосом клетки) имеет длину 3,2 × 106 кб (или 3,2 × 109 пар нуклеотидов).
Химические связи, стабилизирующие вторичную структуру ДНК:
1.Стэкинг-взаимодействия: пары оснований, уложенные в спирали «стопками» удерживаются силами Ван-дер-Ваальса. И несмотря на то, что связи между 2 парами оснований крайне слабые, их большое количество во всей молекуле ДНК является важным фактором её стабилизации.
2.Гидрофобные связи: гидрофобные азотистые основания «погружены» внутрь спирали и взаимодействуют друг с другом, это придаёт спирали дополнительную стабильность.
3.Водородные связи образуются между азотистыми основаниями цепей. Между аденином и тимином — 2 водородных связи, а между гуанином и цитозином — 3 (см. Рис. 52 ).
4.Электростатические взаимодействия: электростатическое отталкива-
ние между отрицательно заряженными остатками фосфорной кислоты дестабилизирует спираль ДНК, однако эти эффекты минимизированы присутствием ионов Mg2+ и катионных белков (богатых лизином и аргинином).

Денатурация и ренатурация ДНК 77
3.5 Денатурация и ренатурация ДНК
ДНК в клетке термодинамически стабильна и спирализована, однако некоторые её области подвергаются «плавлению» (т.е. двойная спираль разматывается) изза происходящих процессов репликации, транскрипции, репарации и рекомбинации. Полное расплетение цепей и их разделение называют денатурацией. Денатурация происходит только in vitro (см. Рис. 53 ).
Двухцепочечная ДНК может быть денатурирована воздействием высоких температур или хаотропных агентов, таких как мочевина или гуанидин хлорид.
При медленном повышении температуры двойная спираль ДНК денатурирует постепенно, за счет нарушения стэкинг и водородных связей. Две цепи постепенно расплетаются и наконец полностью диссоциируют (разделяются). Температура, при которой половина молекулы ДНК подверглась денатурации, называется
точкой плавления Tm.
Ренатурацией называется процесс восстановления нативной кон-
формации ДНК. Ренатурация происходит самопроизвольно при охлаждении раствора, в котором содержатся диссоциировавшие цепи ДНК.
ДНК и РНК могут образовывать гибриды по принципу комплементарности (например, матричная цепь ДНК и транскрибированная с неё пре-мРНК). Кроме того, если смешать в одном растворе денатурированные цепи ДНК различных видов, они тоже будет образовывать гибриды, но в силу того, что полностью нуклеотидные последовательности этих цепей некомплементарны (ввиду генетической изменчивости и разнообразия), такие гибриды будут считаться неполными: помимо спирализованных участков будут образовываться «вздутия», не имеющие спиральной укладки.
Степень денатурации можно измерить с помощью абсорбции ультрафиолетовых лучей. Одноцепочечные ДНК при длине волны 260 нм поглощают 12-40% света. Разумеется, области, богатые парами A = T быстрее подвергаются плавлению, чем Г ≡ Ц участки.
77
78Глава 3 Нуклеиновые кислоты
3.6Третичная структура ДНК
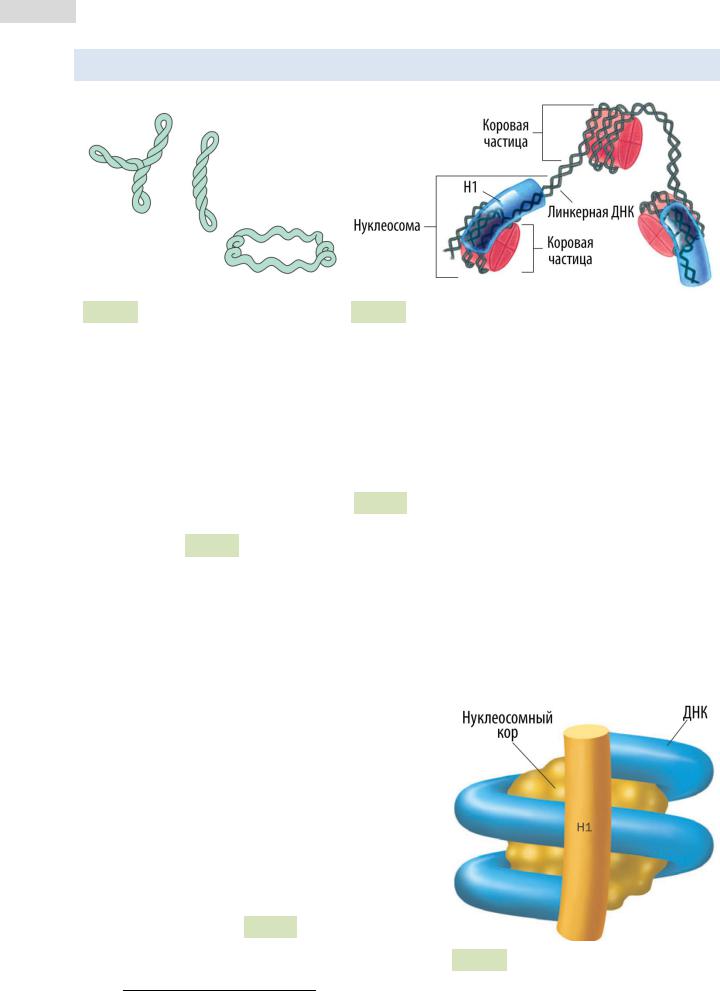
Рис. 54. Отрицательные супер- Рис. 55. Строение нуклеосомной нити витки в кольцевой бактериальной ДНК (слева) и их постепенное раскручивание (справа)
Третичная структура ДНК — это пространственная форма, которую принимает молекула ДНК по мере своего сворачивания и компактизации. Третичная структура ДНК про- и эукариот различается. Прокариотическая ДНК связана лишь с небольшим количеством белков, среди которых есть и гистоноподобные. Она обычно свёрнута в суперспираль округлой формы (см. Рис. 54 ▲). У эукариот степень упаковки ДНК гораздо выше, поскольку их цепи связаны с гистоновыми белками, образующими нук-
леосо́мы (см. Рис. 55 ▲). Нуклеосома — это комплекс нуклеосомного ко́ра и накру-
ченного на него участка ДНК. Каждый нуклеосомный кор состоит из 4 пар гисто́нов (всего 8 белковых субъединиц): H2A, H2B, H3 и H4. Гистоны — положительно заряженные белки (из-за обилия остатков аргинина и лизина), взаимодействующие с отрицательно заряженной ДНК. Комплекс гистонов и ДНК стабилизирован водород-
ными связями, ионными связями (или солевыми мостиками), диполь-дипольными взаи-
модействиями1 (связь между положительно заряженным концом одной молекулы и отрицательно заряженным концом другой).
Вокруг одной нуклеосомной частицы ДНК делает 2,5 оборота (1,75 × 2). Длина участка ДНК, связанного с нуклеосомным кором, составляет 146 пар нуклеотидов. Фрагмент спирали ДНК, находящийся между двумя нуклеосомными частицами и связывающий их, называют линкерным (линкерная ДНК). Гистон H1 в состав нуклеосомного кора не входит, однако связан с молекулой ДНК, которая оборачивается вокруг нуклеосомного кора, в области её «вхождения» и «выхода» из кора (см. Рис. 56 ). Роль H1 заклю-
чается в поддержании плотности упаковки ДНК Рис. 56. Гистон H1 и нуклеосома.
1 См. Таблица 2 (стр. 12) и Рис. 127 (стр. 191)