


46 |
Глава 2 |
Ферменты |
—Гипоксантин + H2O + O2 Ксантин + H2O2
—Ксантин + H2O + O2 Мочевая кислота + H2O2
б. Фермент катализирует две реакции с одним субстратом. Пример:
изоцитратдегидрогеназа сначала окисляет изоцитрат, а затем декарбоксилирует его:
—Изоцитрат + NAD+ → α-Кетоглутарат + CO2 + NADH + H+
2.6Модели образования фермент-субстратного комплекса
Субстратная специфичность фермента связана с расположением аминокислотных радикалов активного центра в пространстве. Именно они формируют особый «карман», с которым связывается субстрат в ходе реакции. Существует две модели, объясняющие связывание субстрата с ферментом.
А Модель «ключ-замок»
Предложена Эми́лем Фишером́ в 1890 г. Согласно его модели, субстрат строго соответствует активному центру фермента, как ключ — замку. Субстрат связывается с ферментом с помощью гидрофобных связей, электростатических взаимодействий, водородных связей и др. Даже малое несоответствие пространственной конфигурации химических групп субстрата расположению радикалов аминокислот в активном центре может привести к стерической некомплементарности и к возникновению сил отталкивания.
Б Модель индуцированного соответствия
Создана Дэ́ниелом Кошланд́ ом в 1958 году. В ней сказано, что комплементарность субстрата ферменту — лишь часть сложной картины взаимодействий между ферментом и субстратом: при связывании субстрата с активным центром происходит изменение конформации молекулы фермента. То есть фермент «подстраивается» под субстрат, как «перчатка под руку».
2.7 Термодинамика ферментативных реакций
Направление химической реакции определяется изменением стандартной свободной энергии (ΔG°) — разностью между энергиями субстратов и продуктов реакции.
Если ΔG отрицательно, то реакция протекает самопроизвольно и сопровождается уменьшением свободной энергии. Такие реакции называют экзергони́ческими. Если при этом абсолютное значение ΔG велико, то реакция идёт практически до конца, и её можно рассматривать как необратимую.
Если ΔG положительно, то реакция будет протекать только при поступлении свободной энергии извне; такие реакции называют эндергони́ческими. Если абсолютное значение ΔG велико, то система устойчива, и реакция в таком случае практически не осуществляется. При ΔG, равном нулю, система находится в равновесии.
Термодинамика ферментативных реакций 47
В биологических системах термодинамически невыгодные (эндергонические) реакции могут протекать лишь за счёт энергии экзергонических реакций. Такие реакции называют энергетически сопряжёнными. Многие из этих реакций происходят при участии аденозинтрифосфата (АТФ), играющего роль сопрягающего фактора.
Для примера рассмотрим реакцию глюкозы и кислорода. Они способны реагировать с образованием углекислого газа и воды:
Глюкоза + O2 → 6CO2 + 6H2O
Однако глюкоза химически стабильна на воздухе (реакция не идёт), несмотря на то, что эта реакция термодинамически вероятна, иными словами спонтанна, поскольку изменение свободной энергии Гиббса для этой реакции G° = −2880 кДж/моль = 689 ккал/моль. То, что эта реакция термодинамически выгодна, не означает, что она мгновенна. Чтобы произошла эта реакция, необходим
приток энергии извне.
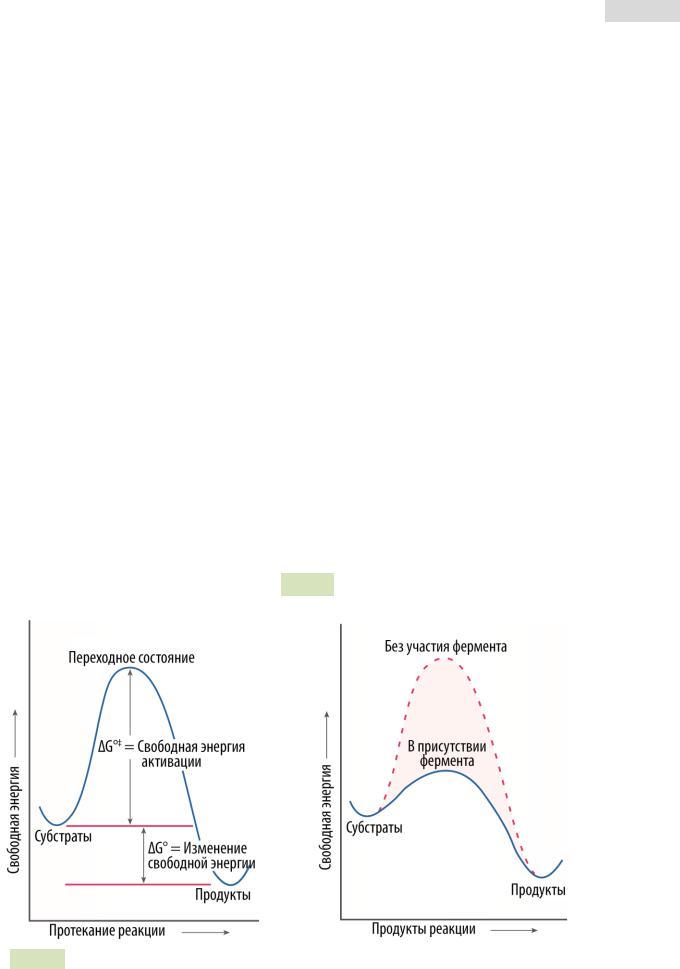
Количество энергии, необходимое для того, чтобы началась реакция, называется энергией активации. Ферменты катализируют реакции, снижая энергию активации G°‡ (или активационную энергию) — энергию, необходимую для того, чтобы реакция началась и было достигнуто переходное состояние. Важной особенностью ферментов является их более высокое сродство к соединениям, уже находящимся в переходном состоянии.
Итак, ферменты ускоряют химические реакции, но не меняют константу равновесия реакции, т.е. изменение свободной энергии. Скорость реакции зависит от свободной энергии активации, т.е. энергии, требуемой для того, чтобы началась реакция. Ферменты снижают активационную энергию, делая реакцию более вероятной и, следовательно, чаще протекающей, отсюда и ускорение реакции. Все вышесказанное проиллюстрировано на графиках (см. Рис. 33 ▼).
Рис. 33. График изменения свободной энергии реакции (слева) и снижение энергии активации ферментом (справа).
47