

Otvety_po_bkh
.pdfКонкурентным ингибитором данной реакции является МАЛОНОВАЯ кислота, поэтому с активным центром фермента связывается и та, и другая кислота, в зависимости от их соотношения в растворе. Чтобы снять частично или полностью действие конкурентного ингибитора, нужно повысить концентрацию субстрата. При этом весь фермент будет находиться в форме фермент-субстратного комплекса, а доля комплекса фермент-ингибитор будет резко понижаться, поэтому скорость ферментативной реакции может быть максимальной даже в присутствии ингибитора. Многие лекарственные препараты действуют по типу конкурентного ингибитора. При этом они тормозят активность ряда ферментов, необходимых для функционирования бактериальных клеток. Примером является применение СУЛЬФАНИЛА (СА). При различных инфекционных заболеваниях, которые вызываются бактериями, применяются СА препараты. Эти препараты имеют структурное сходство с ПАРААМИНОБЕНЗОЙНОЙ кислотой, которая используется бактериями для синтеза ФОЛИЕВОЙ кислоты, необходимой для роста и размножения бактерий.
Введение СА приводит к ИНГИБИРОВАНИЮ фермента бактерий, которые синтезируют ФОЛИЕВУЮ кислоту. Нарушение синтеза этой кислоты проводит к нарушению роста микроорганизмов и их гибели.
По принципу конкурентных ингибиторов действует целая группа различных препаратов -это АНТИХОЛИНЭСТЕРАЗЫ. Они являются конкурентными ингибиторами фермента ХОЛИНЭСТЕРАЗЫ, катализирующего ГИДРОЛИЗ АЦЕТИЛХОЛИНА. АЦЕТИЛХОЛИН обеспечивает проведение нервного импульса. АНТИХОЛИНЭСТЕРАЗЫ конкурируют с АЦЕТИЛХОЛИНОМ за активный центр фермента ХОЛИНЭСТЕРАЗЫ. В результате этого распад АЦЕТИЛХОЛИНА тормозится, он накапливается в организме, вызывая нарушение проведения нервного импульса.
2.НЕКОНКУРЕНТНОЕ ИНГИБИРОВАНИЕ - это торможение ферментативной реакции, вызванное влиянием ингибитора на каталитическое превращение субстрата. При этом ингибитор не влияет на связывание фермента с субстратом. Неконкурентный ингибитор может связываться либо с каталитическими группами активного центра фермента, либо вне активного центра фермента, но при этом он изменяет КОНФОРМАЦИЮ фермента и затрагивает каталитический участок его активного центра. При неконкурентном ИНГИБИРОВАНИИ, возможно образование тройного комплекса.
В качестве неконкурентного ингибитора выступают ЦИАНИДЫ. Они прочно связываются с ионами железа, которые входят в состав каталитического ГЕМИНОВОГО фермента - ЦИТОХРОМОКСИДАЗЫ. Этот фермент является одним из компонентов дыхательной цепи. Блокирование дыхательной цепи выключает её из работы, что приводит к мгновенной гибели организме. Примером неконкурентного ингибитора являются соли тяжёлых металлов. Они блокируют -SH группы, которые входят в каталитический участок фермента. При этом образуется F-I комплекс. Он способен присоединять субстрат, но дальнейшего превращения субстрата не происходит, т.к. каталитические группы фермента заблокированы. Реакция непродуктивна. Снять действие неконкурентного ингибитора очень сложно, т.к. ионы металлов очень прочно связываются с активным центром фермента. Действие этого ингибитора можно снять только с помощью специальных веществ - РЕАКТИВАТОРОВ.
3.СУБСТРАТНОЕ ИНГИБИРОВАНИЕ - это торможение ферментативной реакции, вызванное избытком субстрата. При этом образуется F-S комплекс, но он не подвергается каталитическим
превращениям, т.к. делает молекулу фермента неактивной. Действие субстратного ингибитора снимается путём уменьшения концентрации субстрата.
4.АЛЛОСТЕРИЧЕСКОЕ ИНГИБИРОВАНИЕ характерно для ферментов, имеющих четвертичную структуру, молекула которых состоит из нескольких единиц (ПРОТОМЕРОВ). АЛЛОСТЕРИЧЕСКИЕ ферменты могут иметь 2 и более единиц. При этом одна имеет каталитический центр и называется каталитической, а другая - АЛЛОСТЕРИЧЕСКИЙ центр и называется регуляторной. В отсутствии АЛЛОСТЕРИЧЕСКОГО ИНГИБИТОРА субстрат присоединяется к каталитическому центру, и идёт обычная каталитическая реакция. При появлении АЛЛОСТЕРИЧЕСКОГО ИНГИБИТОРА, он присоединяется к регуляторной единице, т.е. к АЛЛОСТЕРИЧЕСКОМУ центру, и изменяет КОНФОРМАЦИЮ центра фермента, в результате этого активность фермента снижается.
16. Характеристика основных видов регуляции активности ферментов в клетках живого организма.
Регуляция скорости ферментативных реакций осуществляется на 3 независимых уровнях:
изменением количества молекул фермента;
доступностью молекул субстрата и кофермента;
изменением каталитической активности молекулы фермента.
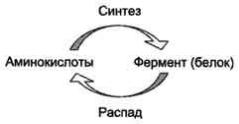
1. Регуляция количества молекул фермента в клетке
Известно, что белки в клетке постоянно обновляются. Количество молекул фермента в клетке определяется соотношением 2 процессов - синтеза и распада белковой молекулы фермента:
Синтез и фолдинг белка - многостадийный процесс. Регуляция синтеза белка может происходить на любой стадии формирования белковой молекулы. Наиболее изучен механизм регуляции синтеза белковой молекулы на уровне транскрипции, который осуществляется определёнными метаболитами, гормонами и рядом биологически активных молекул (см. раздел 4).
Что касается распада ферментов, то регуляция этого процесса менее изучена. Можно только предполагать, что это не просто процесс протеолиза (разрушения белковой молекулы), а сложный механизм, возможно, определяемый на генетическом уровне.
2. Регуляция скорости ферментативной реакции доступностью молекул субстрата и коферментов
Важный параметр, контролирующий протекание метаболического пути, - наличие субстратов, и главным образом - наличие первого субстрата. Чем больше концентрация исходного субстрата, тем выше скорость метаболического пути.
Другой параметр, лимитирующий протекание метаболического пути, - наличие регенерированных коферментов. Например, в реакциях дегидрирования коферментом дегидрогеназ служат окисленные формы NAD+, FAD, FMN, которые восстанавливаются в ходе реакции. Чтобы коферменты вновь участвовали в реакции, необходима их регенерация, т.е. превращение в окисленную форму.
3. Регуляция каталитической активности ферментов
Важнейшее значение в изменении скорости метаболических путей играет регуляция каталитической активности одного или нескольких ключевых ферментов данного метаболического пути. Это высокоэффективный и быстрый способ регуляции метаболизма.
Основные способы регуляции активности ферментов:
аллостерическая регуляция;
регуляция с помощью белок-белковых взаимодействий;
регуляция путём фосфорилирования/дефосфорилирования молекулы фермента;
регуляция частичным (ограниченным) протеолизом.
Аллостерическая регуляция
Аллостерическими ферментами называют ферменты, активность которых регулируется не только количеством молекул субстрата, но и другими веществами, называемыми эффекторами. Участвующие в аллостерической регуляции эффекторы - клеточные метаболиты часто именно того пути, регуляцию которого они осуществляют.
Роль аллостерических ферментов в метаболизме клетки. Аллостерические ферменты играют важную роль в метаболизме, так как они чрезвычайно быстро реагируют на малейшие изменения внутреннего состояния клетки. Аллостерическая регуляция имеет большое значение в следующих ситуациях:
при анаболических процессах. Ингибирование конечным продуктом метаболического пути и активация начальными метаболитами позволяют осуществлять регуляцию синтеза этих соединений;
при катаболических процессах. В случае накопления АТФ в клетке происходит ингибирование метаболических путей, обеспечивающих синтез энергии. Субстраты при этом расходуются на реакции запасания резервных питательных веществ;
для координации анаболических и катаболических путей. АТФ и АДФ - аллостерические эффекторы, действующие как антагонисты;
для координации параллельно протекающих и взаимосвязанных метаболических путей
(например, синтез пуриновых и пиримидиновых нуклеотидов, используемых для синтеза нуклеиновых кислот). Таким образом, конечные продукты одного метаболического пути могут быть аллостерическими эффекторами другого метаболического пути.
Аллостерические эффекторы. Эффектор, вызывающий снижение (ингибирование) активности фермента, называют отрицательным эффектором, или ингибитором. Эффектор, вызьгоаюший повышение (активацию) активности ферментов, называют положительным эффектором, или активатором.
Аллостерическими эффекторами часто служат различные метаболиты. Конечные продукты метаболического пути - часто ингибиторы аллостерических ферментов, а исходные вещества - активаторы. Это так называемая гетеротропная регуляция. Такой вид аллостерической регуляции очень распространён в биологических системах.
Более редкий случай аллостерической регуляции, когда сам субстрат может выступать в качестве положительного эффектора. Такая регуляция называется гомотропной (эффектор и субстрат - одно и то же вещество). Эти ферменты имеют несколько центров связывания для субстрата, которые могут выполнять двойную функцию: каталитическую и регуляторную. Аллостерические ферменты такого типа используются в ситуации, когда субстрат накапливается в избытке и должен быстро преобразоваться в продукт.
Выявить ферменты с аллостерической регуляцией можно, изучая кинетику этих ферментов. Эти ферменты не подчиняются законам Михаэлиса-Ментен, они имеют характерную S-образную кривую зависимости скорости реакции от концентрации субстрата.
Особенности строения и функционирования аллостерических ферментов:
обычно это олигомерные белки, состоящие из нескольких протомеров или имеющие доменное строение;
они имеют аллостерический центр, пространственно удалённый от каталитического активного центра;
эффекторы присоединяются к ферменту нековалентно в аллостерических (регуляторных) центрах;
аллостерические центры, так же, как и каталитические, могут проявлять различную специфичность по отношению к лигандам: она может быть абсолютной и групповой. Некоторые ферменты имеют несколько аллостерических центров, одни из которых специфичны к активаторам, другие - к ингибиторам.
протомер, на котором находится аллостерический центр, - регуляторный протомер, в отличие от каталитического протомера, содержащего активный центр, в котором проходит химическая реакция;
аллостерические ферменты обладают свойством кооперативности: взаимодействие аллостерического эффектора с аллостерическим центром вызывает последовательное кооперативное изменение конформации всех субъединиц, приводящее к изменению
конформации активного центра и изменению сродства фермента к субстрату, что снижает или увеличивает каталитическую активность фермента (рис. 2-30);
регуляция аллостерических ферментов обратима: отсоединение эффектора от регуляторной субъединицы восстанавливает исходную каталитическую активность фермента;
аллостерические ферменты катализируют ключевые реакции данного метаболического пути.
Локализация аллостерических ферментов в метаболическом пути. Скорость метаболических процессов зависит от концентрации веществ, использующихся и образующихся в данной цепи реакций. Такая регуляция представляется логичной, так как при накоплении конечного продукта он (конечный продукт) может действовать как аллостерический ингибитор фермента, катализирующего чаще всего начальный этап данного метаболического пути:
Фермент, катализирующий превращение субстрата А в продукт В, имеет аллостерический центр для отрицательного эффектора, которым служит конечный продукт метаболического пути F. Если концентрация F увеличивается (т.е. вещество F синтезируется быстрее, чем расходуется), ингибируется активность одного из начальных ферментов. Такую регуляцию называют отрицательной обратной связью, или ретроингибировани-ем. Отрицательная обратная связь - часто встречающийся механизм регуляции метаболизма в клетке.
В центральных метаболических путях исходные вещества могут быть активаторами ключевых ферментов метаболического пути. Как правило, при этом аллостерической активации подвергаются ферменты, катализирующие ключевые реакции заключительных этапов метаболического пути:
В качестве примера можно рассмотреть принципы регуляции гликолиза - специфического
(начального) пути распада глюкозы (рис. 2-31). Один из конечных продуктов распада глюкозы - молекула АТФ. При избытке в клетке АТФ происходит ретро-ингибирование аллостерических ферментов фосфофруктокиназы и пируваткиназы. При образовании большого количества фруктозо-1,6-бисфосфата наблюдают аллостерическую активацию фермента пируваткиназы.
Благодаря такой регуляции осуществляется слаженность протекания метаболического пути распада глюкозы.
Регуляция каталитической активности ферментов белок-белковыми взаимодействиями. Некоторые ферменты изменяют свою каталитическую активность в результате белок-белковых взаимодействий. Рассмотрим 2 механизма активации ферментов с помощью белок-белковых взаимодействий:
активация ферментов в результате присоединения регуляторных белков;
изменение каталитической активности ферментов вследствие ассоциации или диссоциации протомеров фермента.
Активация ферментов в результате присоединения регуляторных белков. Этот тип регуляции можно рассмотреть на примере активации фермента аденилатциклазы, локализованной
вплазматической мембране клетки.
Активный центр аденилатциклазы локализован на цитоплазматической стороне плазматической мембраны. Активированная аденилатциклаза катализирует реакцию образования из АТФ циклического 3',5'-АМФ (цАМФ) - вторичного, внутриклеточного посредника действия гормонов (см. схему ниже).
В мембране аденилатциклаза функционирует в комплексе с другими белками:
рецептором гормона, выступающего во внеклеточную среду и взаимодействующего с гормонами;
с G-белком, занимающим промежуточное положение между рецептором и ферментом аденилатциклазой. G-белок - олиго-мерный белок, состоящий из 3 субъединиц - α, β, γ. α- Субъединица имеет центр связывания и расщепления ГТФ. Поэтому этот белок называется ГТФ-связывающим белком, или G-белком;
в результате связывания гормона с рецептором происходит изменение конформа-ции G- белка, уменьшение его сродства к молекуле ГДФ, с которой он связан в отсутствие гормонального сигнала, и увеличение сродства к ГТФ. Присоединение ГТФ вызывает

конформационные изменения в G-белке и диссоциацию его на субъединицы: субъединицу α, связанную с ГТФ (α-ГТФ), димер βγ;
α-ГТФ имеет высокое сродство к аденилатциклазе, его присоединение приводит к активации последней, поэтому α-ГТФ - регуляторный белок, а данный механизм активации аденилатциклазы называют активацией ферментов в результате присоединения регуляторных белков.
Регуляция каталитической активности ферментов ассоциацией/диссоциацией протомеров Протеинкиназы - группа ферментов, катализирующих перенос остатка фосфорной кислоты с АТФ на специфические ОН-группы аминокислотных остатков белков (вызывают фосфорилирование белков). Механизмы активации различных протеинкиназ неодинаковы. В качестве примера регуляции каталитической активности ферментов ассоциацией или диссоциацией протомеров можно привести регуляцию активности фермента Протеинкиназы А.
Протеинкиназа А (цАМФ-зависимая) состоит из 4 субъединиц 2 типов: 2 регуляторных
(R)и 2 каталитических (С). Такой тетрамер не обладает каталитической активностью. Регуляторные субъединицы имеют участки связывания для циклического 3',5'-АМФ (цАМФ), по 2 на каждую субъединицу. Присоединение 4 молекул цАМФ к 2 регуляторным субъединицам приводит к изменению конфор-мации регуляторных протомеров и к диссоциации тетрамерного комплекса, при этом высвобождаются 2 активные каталитические субъединицы. Такой механизм регуляции обратим. Отщепление молекул цАМФ от регуляторных субъединиц приведёт к ассоциации регуляторных и каталитических субъединиц Протеинкиназы А с образованием неактивного комплекса.
115
Регуляция каталитической активности ферментов путём фосфорилирования/дефосфорилирования
В биологических системах часто встречается механизм регуляции активности ферментов с помощью ковалентной модификации аминокислотных остатков. Быстрый и широко распространённый способ химической модификации ферментов - фосфорилирование/дефосфорилирование. Модификации подвергаются ОН-группы фермента. Фос-форилирование осуществляется ферментами протеинкиназами, а дефосфорилирование - фосфопротеинфосфатазами. Присоединение остатка фосфорной кислоты приводит к изменению
конформации активного центра и его каталитической активности. При этом результат может быть двояким: одни ферменты при фосфорилировании активируются, другие, напротив, становятся менее активными (рис. 2-33).
Изменение активности фермента, вызванное фосфорилированием, обратимо. Отщепление остатка фосфорной кислоты осуществляется ферментами фосфопротеинфосфатазами. Активность протеинкиназ и фосфопротеинфосфатаз регулируется гормонами, что позволяет быстро изменять активность ключевых ферментов метаболических путей в зависимости от условий внешней среды. Антагонистичные по функции гормоны противоположным образом влияют на фосфорилирование/дефосфорилирование ферментов, вызывая противоположные эффекты изменения метаболизма клетки.
Например, под действием глюкагона (в период между приёмами пищи) в клетках происходит уменьшение синтеза энергетического материала - жира, гликогена и усиление его распада (мобилизация), вызванного фосфо-рилированием ключевых ферментов этих процессов. А под действием инсулина (во время пищеварения), наоборот, активируется синтез гликогена и

ингибируется его распад, так как взаимодействие инсулина с рецептором активирует сигнальный путь, приводящий к дефосфорилированию тех же ключевых ферментов.
Регуляция каталитической активности ферментов частичным (ограниченным) протеолизом
Некоторые ферменты, функционирующие вне клеток (в ЖКТ или в плазме крови), синтезируются в виде неактивных предшественников и активируются только в результате гидролиза одной или нескольких определённых пептидных связей, что приводит к отщеплению части белковой молекулы предшественника. В результате в оставшейся части белковой молекулы происходит конформационная перестройка и формируется активный центр фермента.
Рассмотрим механизм частичного протеолиза на примере активации протеолитического фермента трипсина. Трип-синоген, синтезируемый в поджелудочной железе, при пищеварении по протокам поджелудочной железы поступает в двенадцатиперстную кишку, где и активируется путём частичного протеолиза под действием фермента кишечника энтеропептидазы. В результате отщепления гексапептида с N-конца формируется активный центр в оставшейся части молекулы. Следует напомнить, что трипсин относят к семейству "сериновых" протеаз - активный центр фермента содержит функционально важный остаток Сер.
Частичный протеолиз - пример регуляции, когда активность фермента изменяется необратимо. Такие ферменты функционируют, как правило, в течение короткого времени, определяемого временем жизни белковой молекулы. Частичный протеолиз лежит в основе активации протеолитических ферментов, белков свёртывающей системы крови и фибринолиза, белков системы комплемента, а также пептидных гормонов.
17.Обмен веществ и энергии. Характеристика основных этапов обмена веществ. Общий и
специфические пути катаболизма. Окислительное декарбоксилирование пирувата.
Обмен веществ - необходимое условие жизни, но он протекает и вне живой природы. Обмен веществ всегда связан с обменом энергии. Системы, обменивающиеся с окружающей средой веществом и энергией, называются открытыми (живой организм). Закрытые системы обмениваются только энергией. В соответствии со 2 законом термодинамики, открытые системы являются более устойчивыми. Отличительной особенностью обмена веществ в живой природе от обмена веществ в неживой системе является способность образовывать дочерние структуры из поступивших веществ. Обмен веществ - это процессы взаимоотношения организма с внешней средой, представляющие совокупность химических реакций, которым подвергаются различные вещества с момента их поступления в организм до момента их выделения в виде конечных продуктов. Основные компоненты живых систем - белки, жиры, углеводы. В организм человека должно поступать: Белков - 100 г. в сутки, Жиров - 100 г. в сутки, Углеводов - 400 г. в сутки.
За сутки при обмене этих веществ образуется 2000-3000 ккал энергии. ЭТАПЫ ОБМЕНА ВЕЩЕСТВ.
1.Поступление веществ из внешней среды посредством питания и дыхания. 2.Превращение веществ в организме - МЕЖУТОЧНЫЙ обмен 3.Выделение конечных продуктов.
Распад БЖУ до ПИРУВАТА идёт индивидуально для каждой группы органических соединений и носит название специфического пути катаболизма. С образованием ПИРУВАТА начинается общий путь катаболизма, идентичный для обмена всех питательных веществ.

Пируват подвергается реакции окислительного декабоксилирования. Этот процесс катализируется комплексом ферментов - пируватдекарбоксилазным комплексом (ПДК) – 3 фермента, с участием в процессе 5 коферментов:
1 .Декарбоксилаза (ДК) (кофермент - ТДФ).
2.Ацетилтрансфераза (АТ) (включают коферменты - ЛК, HSKоA). 3.Дегидрогеназа (ДГ) (включают коферменты - НАД, ФАД).
Все ферменты этого комплекса располагаются в пространстве, поэтому промежуточные метаболиты передаются от одного активного центра к другому и все вместе формируют мультиферментный комплекс. Образующийся ацетилКоА далее вступает в цитратный цикл.
18. Современные представления о биологическом окислении. Компоненты дыхательной цепи и их характеристика.
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О БИОЛОГИЧЕСКОМ ОКИСЛЕНИИ.
1. Биологическое окисление, как и окисление вообще, есть процесс переноса электронов. То вещество, что отдаёт электроны, окисляется, то, что принимает, восстанавливается. Если акцептором электронов является кислород, то такой процесс называется ТКАНЕВЫМ ДЫХАНИЕМ. Биологическое окисление предполагает ДЕГИДРИРОВАНИЕ с образованием воды.
Если водород взаимодействует с кислородом с образованием воды вне организма, то это сопровождается взрывом.
2.Биологическое окисление - это процесс многоступенчатый - многоступенчатая передача электронов с постепенным выделением энергии, что исключает взрыв.
3.Биологическое окисление - это процесс, требующий много ферментов. Т.о. биологическое окисление - это многоступенчатый процесс транспорта электронов, осуществляемый комплексов ферментов. Этот комплекс ферментов называется ЭЛЕКТРОНТРАНСПОРТНОЙ ЦЕПЬЮ (ЭТЦ), или ЦЕПЬЮ ПЕРЕНОСА ЭЛЕКТРОНОВ (ЦПЭ), или дыхательной цепью. ЭТЦ - это своеобразный КОНВЕЕР по переносу электронов и протонов от субстрата к кислороду.
КОМПОНЕНТЫ ДЫХАТЕЛЬНОЙ ЦЕПИ.
1.НИКОТИНЗАВИСИМЫЕ ДГ, т.е. содержащие КОФЕРМЁНТЫ - НАД, НАДФ 2.ФЛАВИНЗАВИСИМЫЕ ДГ, т.е. содержащие КОФЕРМЁНТЫ - ФМН, ФАД. З.УБИХИНОН (Ko-Q).
4.ЦИТОХРОМЫ: в, с, c1., а, а3.
Почти все эти компоненты, за исключением первого, встроены во внутреннюю мембрану МИТОХОНДРИЙ. В печени таких дыхательных цепей до 5000, а в сердце - до 20000. СТРОЕНИЕ КОМПОНЕНТОВ ДЫХАТЕЛЬНОЙ ЦЕПИ.
1.В НАД и НАДФ рабочей частью является витамин РР - НИКОТИНАМИД.

НАД*2Н + 2е = НАДН+Н 2.В ФАД и ФМН рабочей частью является ФЛАВИИ (компонент витамина В2)
ФАД + 2Н + 2е = ФАДН2
3.УБИХИНОН легко переходит в восстановленную форму KOQ +2Н +2е =KOQ*H2 4.ЦИТОХРОМЫ - это ГЕТЕРОПРОТЕИНЫ. Их белковой частью является ГЕМ, структура которого представляет собой 4 ПИРРОЛЬНЫХ кольца и атом железа, который легко меняет валентность. Также могут включать медь.
ФЕРМЕНТЫ ДЫХАТЕЛЬНОЙ ЦЕПИ.
1 .ДГ субстратов находятся в цитоплазме клетки, могут быть в МАТРИКСЕ МИТОХОНДРИЙ. 2.НАДН-ДГ(ФМН).
3.KOQ
4.Q*H2 - ДГ (ЦИТОХРОМЫ в, с 1). 5.ЦИТОХРОМ с.
6.ЦИТОХРОМОКСИДАЗА участвует в передаче электронов на кислород (включает ЦИТОХРОМЫ а,
а3).
ФУНКЦИОНИРОВАНИЕ ДЫХАТЕЛЬНОЙ ЦЕПИ.
Полная ЭТЦ - взаимодействие субстрата с НАД. Укороченная ЭТЦ - взаимодействие субстрата с ФАД и последующий транспорт электронов и протонов сразу на КОФЕРМЕНТ Q,
Порядок компонентов дыхательной цепи обусловлен величиной их red-ox потенциалов. Он изменяется от -0,32В до +0,81В -0,32 характерно для НАДН2 +0,81 характерно для О2.
19.Пути синтеза АТФ в клетках, клеточная локализация процессов синтеза АТФ, примеры реакций. Молекулярные механизмы окислительного фосфорилирования (теория
Митчелла).
ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИИАНИЕ.
В дыхательной цепи создаются условия для синтеза АТФ, т.е. выделяется достаточное количество энергии. Процесс образования АТФ из АДФ и Н3РО4 за счёт энергии переноса электронов дыхательной цепи называется ОКИСЛИТЕЛЬНЫМ ФОСФОРИЛИРОВАНИЕМ. СУБСТРАТНОЕ ФОСФОРИЛИРОВАНИЕ - это процесс образования АТФ из АДФ и Н3РО4 за счёт энергии распада какого-либо субстрата. В дыхательной цепи выделяются 3 пункта, где может образоваться АТФ:
1.НАД KOQ
2.ЦИТ. в. ЦИТ. с З.ЦИТ. а. ЦИТ. A3 НАДН2 — 3 АТФ ФАДН2 — 2 АТФ
Процесс освобождения АТФ с транспортом электронов в дыхательной цепи называется СОПРЯЖЕНИЕМ ОКИСЛЕНИЯ И ФОСФОРИЛИРОВАНИЯ. Но может быть разобщение ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ (свободное окисление), т.е. окисление идёт, а ФОСФОРИЛИРОВАНИЯ нет, вся энергия выделяется в виде тепла. Это обуславливает ПИРОГЕННЫЙ эффект ряда лекарственных веществ.
Лекция № 9. Биологическое окисление (продолжение).
1 .МЕХАНИЗМ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ.
2.АЛЬТЕРНАТИВНЫЕ ПУТИ БИОЛОГИЧЕСКОГО ОКИСЛЕНИЯ.
3.СВОБОДНО-РАДИКАЛЬНОЕ ОКИСЛЕНИЕ.
МЕХАНИЗМ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ.
Теория ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ выдвинута английским учёным П. МИТЧЕЛОМ в 1961 г. и названа ХЕМИООСМОТИЧЕСКОЙ ТЕОРИЕЙ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ. Он объяснил процесс синтеза АТФ с биохимической позиции, но его взгляды не получили признания. Однако в последующем его теория подтвердилась, и через 17 лет он был удостоен Нобелевской премии.
Основные положения теории:
1.Мембрана МИТОХОНДРИЙ не проницаема для протонов.
2.Образуется протонный потенциал в процессе транспорта электронов и протонов. 3.Обратный транспорт протонов в МАТРИКС сопряжен с образованием АТФ.
Процесс транспорта электронов происходит во внутренней мембране. Первые реакции окисления происходят в матрице. Протоны переносятся в межмембранное пространство, а электроны продвигаются по дыхательной цепи. В процессе работы дыхательной цепи внутренняя мембрана со стороны матрицы заряжается отрицательно, а со стороны межмембранного пространства положительно. Следовательно, возникает разность потенциалов, градиент концентрации ионов, и, соответственно, градиент РН. Т.о. РН со стороны матрицы будет менее кислая. Во время дыхания создаётся ЭЛЕКТРО-ХИМИЧЕСКИЙ градиент: концентрационный и разности потенциалов. Электрический и концентрационный градиент составляет ПРОТОНДВИЖУЩУЮ силу, которая даёт силу для синтеза АТФ. На определённых участках внутренней мембраны есть протонные каналы, образованные АТФ-СИНТЕТАЗОЙ. Протоны могут проходить обратно в матрицу, при этом образующаяся энергия идёт на синтез АТФ.
УСЛОВИЯ ОБРАЗОВАНИЯ АТФ.
1.Целостность мембраны - непроницаемость её для протонов.
2.Наличие специальных каналов.
3.Движение протонов в матрицу сопровождается выделением энергии, используемой для синтеза АТФ.
Вопрос о том, что позволяет протонам переходить в межмембранное пространство остаётся не вполне ясным.
Основные компоненты ЭТЦ представляют собой интегральные белки и фиксированные в мембране: 1.НАДН-ДГ.
2.QН2-ДГ.
3.ЦИТОХРОМОКСИДАЗА. 4.KOQ не связан с белками.
5.ЦИТОХРОМ с - не фиксирован к мембране. Выдвигается теория Q-цикла транспорта протонов.

2Н + 2е + KOQ KOQ*H2
KOQ*H2 KOQ + 2Н + 2е - на наружной поверхности внутренней мембраны.
Т.о. в соответствии с ХЕМООСМОТИЧЕСКОЙ теорией МИТЧЕЛА окисление НАДН2 и ФАДН2 в дыхательной цепи создаёт сначала ЭЛЕКТРОНО-ХИМИЧЕСКИЙ протонный потенциал, градиент концентрации ионов на внутренней мембране, а обратный транспорт протонов через мембрану сопряжен с ФОСФОРИЛИРОВАНИЕМ, т.е. образованием АТФ.
АЛЬТЕРНАТИВНЫЕ ПУТИ БИОЛОГИЧЕСКОГО ОКИСЛЕНИЯ.
В организме возможен и ОКСИГЕНАЗНЫЙ путь биологического окисления. Он не относится к процессам, сопровождающимся выделением энергии, он не снабжает клетку энергией. Ферменты этого пути включают кислород и субстрат. Этот путь характерен для ДЕГИДРАТАЦИИ различных метаболитов, чаще всего чужеродных.
Стадии ОКСИГЕНАЗНОГО ПУТИ:
1.Связывание кислорода с активным центром фермента.
2.Восстановление кислорода и перенос его на субстрат. Выделяют 2 типа ОКСИГЕНАЗ:
1.ДИОКСИГЕНАЗЫ - ферменты, включающие в субстрат молекулу кислорода.
А + О2 = АО2 В живых тканях этот процесс практически не встречается.
2.МОНООКСИГЕНАЗЫ - они катализируют включение в субстрат 1 атома кислорода, др. атом кислорода восстанавливается до воды. Для реакций катализируемых МОНООКСИГЕНАЗАМИ необходим КОСУБСТРАТ - донор электронов.
А-Н + О2 + ZH2 А-ОН + Z + Н2О Где А-Н - субстрат
ZH2 - КОСУБСТРАТ
А-ОН - окисленный субстрат.
В организме есть несколько видов МОНООКСИГЕНАЗ и прежде всего МИКРОСОМАЛЬНЫЕ МОНООКСИГЕН АЗЫ, содержащие ЦИТОХРОМ Р-450.Т.к. образуется -ОН группа, то реакции называются ещё реакциями ГИДРОКСИЛИРОВАНИЯ. МИКРОСОМАЛЬНАЯ система участвует в деградации многих умеренно токсических соединений, лекарственных веществ. Восстановленным КОСУБСТРАТОМ в этих реакциях является НАДФ*Н2. Этот путь окисления иногда называют ГИДРОКСИЛАЗНЫМ ЦИКЛОМ.
СВОБОДНОЕ РАДИКАЛЬНОЕ ОКИСЛЕНИЕ.
Свободные радикалы - это молекулярные частицы, у которых на внешней оболочке имеется не спаренный электрон. Они могут образовываться: при окислении (отрывании атома водорода)
при восстановлении О2 + е О2-
Атомарный кислород имеет на внешнем электронном уровне 2 неспареных электрона. Он не слишком активный, но может образовывать высоко активные формы.
О2+4е + 4Н 2Н2О Этот процесс, в тканях, идёт постепенно, с переносом 1е на каждом этапе.
О2 + е = 02- - СУПЕРОКСИДНЫЙ АНИОН 2-02 + 2е = 022- - ПЕРОКСИДНЫЙ АНИОН НО, - ГИДРОКСИЛ РАДИКАЛ
Н2О2, О2-, О22-, ОН, - активные формы кислорода (АФК)
Они образуются в организме при различных физиологических и патологических процессах. Все свободные радикалы в организме классифицируют:
1.Первичные радикалы (О2, N,O). Они образуются в результате ферментативных реакций. Они являются физиологическими. Способствуют образованию РАДИКАЛОБРАЗНЫХ молекул, к которым относятся НООН, они вызывают образование вторичных радикалов.
2.Вторичные радикалы (ОН, ЛИПИДНЫЕ радикалы – L,, LO,, LOO,) Их образование происходит с участием железа (11). Это патологические продукты.