

Том1_1
.pdf193
повреждающим воздействиям, так как локальные повреждения всегда компенсируются за счет сохранившихся элементов сети. С другой стороны, высокая надежность функционирования РФ обеспечивается тем, что раздражение любой из ее частей отражается на активности всей РФ данной структуры за счет диффузности связей.
Большинство нейронов РФ имеет длинные дендриты и короткий аксон. Существуют гигантские нейроны с длинным аксоном, образующие пути из РФ в другие области мозга, например в нисходящем направлении, ретикулоспинальный и руброспинальный. Аксоны нейронов РФ образуют большое число коллатералей и синапсов, которые оканчиваются на нейронах различных отделов мозга. Аксоны нейронов РФ, идущие в кору большого мозга, заканчиваются здесь на дендритах I и II слоев.
Активность нейронов РФ различна и в принципе сходна с активностью нейронов других структур мозга, но среди нейронов РФ имеются такие, которые обладают устойчивой ритмической активностью, не зависящей от приходящих сигналов.
Вто же время в РФ среднего мозга и моста имеются нейроны, которые в покое «молчат», т. е. не генерируют импульсы, но возбуждаются при стимуляции зрительных или слуховых рецепторов. Это так называемые специфические нейроны, обеспечивающие быструю реакцию на внезапные, неопознанные сигналы. Значительное число нейронов РФ являются полисенсорными.
ВРФ продолговатого, среднего мозга и моста конвергируют сигналы различной сенсорности. На нейроны моста приходят сигналы преимущественно от соматосенсорных систем. Сигналы от зрительной и слуховой сенсорных систем в основном приходят на нейроны РФ среднего мозга.
РФ контролирует передачу сенсорной информации, идущей через ядра таламуса, за счет того, что при интенсивном внешнем раздражении нейроны неспецифических ядер таламуса затормаживаются, тем самым снимается их тормозящее влияние с релейных ядер того же таламуса и облегчается передача сенсорной информации в кору большого мозга.
194
В РФ моста, продолговатого, среднего мозга имеются нейроны, которые реагируют на болевые раздражения, идущие от мышц или внутренних органов, что создает общее диффузное дискомфортное, не всегда четко локализуемое, болевое ощущение «тупой боли».
Повторение любого вида стимуляции приводит к снижению импульсной активности нейронов РФ, т. е. процессы адаптации (привыкания) присущи и нейронам РФ ствола мозга.
РФ ствола мозга имеет прямое отношение к регуляции мышечного тонуса, поскольку на РФ ствола мозга поступают сигналы от зрительного и вестибулярного анализаторов и мозжечка. От РФ к мотонейронам спинного мозга и ядер черепных нервов поступают сигналы, организующие положение головы, туловища и т. д.
Ретикулярные пути, облегчающие активность моторных систем спинного мозга, берут начало от всех отделов РФ. Пути, идущие от моста, тормозят активность мотонейронов спинного мозга, иннервирующих мышцы-сгибатели, и активируют мотонейроны мышц-разгибателей. Пути, идущие от РФ продолговатого мозга, вызывают противоположные эффекты. Раздражение РФ приводит к тремору, повышению тонуса мышц. После прекращения раздражения вызванный им эффект сохраняется длительно, видимо, за счет циркуляции возбуждения в сети нейронов.
РФ ствола мозга участвует в передаче информации от коры большого мозга, спинного мозга к мозжечку и, наоборот, от мозжечка к этим же системам. Функция данных связей заключается в подготовке и реализации моторики, связанной с привыканием, ориентировочными реакциями, болевыми реакциями, организацией ходьбы, движениями глаз.
Регуляция вегетативной деятельности организма РФ проявляется в функционировании дыхательного и сердечнососудистых центров. В регуляции вегетативных функций большое значение имеют так называемые стартовые нейроны РФ. Они дают начало циркуляции возбуждения внутри группы нейронов, обеспечивая тонус регулируемых вегетативных систем.
Влияния РФ можно разделить в целом на нисходящие и восходящие. В свою очередь каждое из этих влияний имеет
195
тормозное и возбуждающее действие.
Восходящие влияния РФ на кору большого мозга повышают ее тонус, регулируют возбудимость ее нейронов, не изменяя специфику ответов на адекватные раздражения. РФ влияет на функциональное состояние всех сенсорных областей мозга, следовательно, она имеет значение в интеграции сенсорной информации от разных анализаторов.
РФ имеет прямое отношение к регуляции цикла бодрствование—сон. Стимуляция одних структур РФ приводит к развитию сна, стимуляция других вызывает пробуждение. Г. Мэгун и Д. Моруцци выдвинули концепцию, согласно которой все виды сигналов, идущих от периферических рецепторов, достигают по коллатералям РФ продолговатого мозга и моста, где переключаются на нейроны, дающие восходящие пути в таламус и затем в кору большого мозга.
Возбуждение РФ продолговатого мозга или моста вызывает синхронизацию активности коры большого мозга, появление медленных ритмов в ее электрических показателях, сонное торможение.
Возбуждение РФ среднего мозга вызывает противоположный эффект пробуждения: десинхронизацию электрической активности коры, появление быстрых низкоамплитудных β-подобных ритмов в электроэнцефалограмме.
Bremer F., (1938) показал, что если перерезать мозг между передними и задними буграми четверохолмия, то животное перестает реагировать на все виды сигналов; если же перерезку произвести между продолговатым и средним мозгом (при этом РФ сохраняет связь с передним мозгом), то животное реагирует на свет, звук и другие сигналы. Следовательно, поддержание активного анализирующего состояния мозга возможно при сохранении связи с передним мозгом.
Реакция активации коры большого мозга наблюдается при раздражении РФ продолговатого, среднего, промежуточного мозга. В то же время раздражение некоторых ядер таламуса приводит к возникновению ограниченных локальных участков возбуждения, а не к общему ее возбуждению, как это бывает при раздражении других отделов РФ.
196
РФ ствола мозга может оказывать не только возбуждающее, но и тормозное влияние на активность коры мозга.
Нисходящие влияния РФ ствола мозга на регуляторную деятельность спинного мозга были установлены еще И. М. Сеченовым (1862). Им было показано, что при раздражении среднего мозга кристалликами соли у лягушки рефлексы отдергивания лапки возникают медленно, требуют более сильного раздражения или не появляются вообще, т. е. тормозятся.
Г. Мэгун (1945—1950), нанося локальные раздражения на РФ продолговатого мозга, нашел, что при раздражении одних точек тормозятся, становятся вялыми рефлексы сгибания передней лапы, коленный, роговичный. При раздражении РФ в других точках продолговатого мозга эти же рефлексы вызывались легче, были сильнее, т. е. их реализация облегчалась. По мнению Мэгуна, тормозные влияния на рефлексы спинного мозга может оказывать только РФ продолговатого мозга, а облегчающие влияния регулируются всей РФ ствола и спинного мозга.
3.7.4.1. Роль ретикулярной формации ствола мозга в механизмах коркового электрогенеза
Проведенными исследованиями (Лобасюк Б.А., 2005), выявлено, что при отведении ЭКоГ от препарата изолированного переднего мозга, который можно рассматривать как модель мозга организма, находящегося в патологическом коматозном состоянии, по сравнению с условиями интактности мозга отмечались общее снижение амплитуд ритмов ЭКоГ и реорганизация этих ритмов. Последняя выражалась в уменьшении индексов бета-2- и тетаритмов и увеличении соответствующих индексов бета-1- и дельта-ритмов. Показано (Русинов В. С.,. Гриндель О. М, Болдырева Г. Н., 1988), что у больных с очаговыми поражениями диэнцефальных отделов мозга спектральная характеристика ЭЭГ смещается в сторону преобладания низких частот. Данный сдвиг, по мнению авторов, свидетельствует об утрате корой в подобных условиях её
197
ведущей роли и переходе к регуляции массовой электрической активности мозга филогенетически более старыми образованиями – в значительной степени лимбической системой.
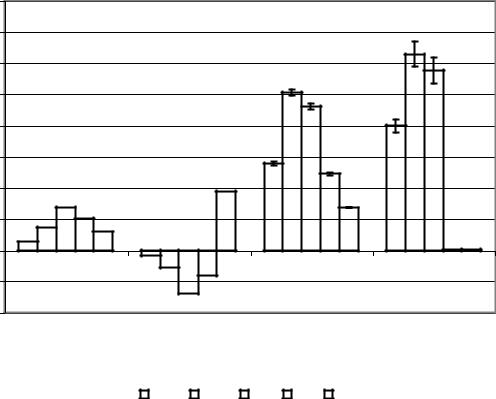
Следует отметить, что в условиях наших опытов уменьшение индекса длительности тета-ритма и, соответственно, увеличение индекса длительности дельтаритма при отведении от препарата ИПМ по сравнению с таковым в условиях ИМ были более выраженными в правом полушарии, чем в левом. ЭКоГ, отводимая от препарата ИПМ, характеризовалась бóльшими значениями индексов длительности альфа-ритма на отрезках, как десинхронизации, так и синхронизации по сравнению с картиной, наблюдаемой в условиях интактности мозга. Переход от десинхронизации к синхронизации ЭКоГ при отведении от интактного мозга (рис. 17) сопровождался сменой выраженной левосторонней латерализации амплитуд бета-2-, бета-1-, альфа- и тета-ритмов правосторонней и увеличением левосторонней латерализации амплитуды дельта-ритма. Полученные результаты в определенной степени согласуются с представлениями о возможности смены доминантности полушарий в разные фазы сна и об относительном повышении активности правого полушария во время быстрого и левого – во время медленного сна.
198
%
40
35
30
25
20
15
10
5
0
-5
-10
int_desin |
int_sin |
si_des |
si_sin |
|
группы опытов - ритмы ЭКоГ |
||
|
B2A B1A AA |
TA |
DA |
Рис. 17. Диаграммы коэффициентов функциональной межполушарной асимметрии амплитуд ритмов 2, 1, и ЭКоГ, отводимой от интактного мозга и препарата изолированного мозга, int_desin – интактный десинхронизация, int_sin – интактный синхронизация, si_desin – препарат изолированного переднего мозга десинхронизация, si_sin - препарат изолированного переднего мозга синхронизация.
Переход от десинхронизации к синхронизации при отведении от препарата ИПМ сопровождался увеличением левосторонней латерализации амплитуд бета-2-, бета-1- и альфа-ритмов и уменьшением левосторонней латерализации амплитуд тета- и дельта-ритмов.
С учетом описанных выше данных можно высказать предположение, что структуры ствола (в частности, РФ) и мозжечка наряду с собственно корковыми механизмами в значительной степени обеспечивают управление ФМПА в условиях интактности мозга. Следует подчеркнуть, что показатели ФПМА в условиях ИПМ не только были положительными, т.е. проявлялась отчетливая латерализация электрической активности в левое полушарие, но и при этом их
199
модули в два–пять раз превышали аналогичные величины, характерные для ИМ. Указанная особенность, возможно, связана с тем обстоятельством, что связи полушарий со срединными образованиями мозга разного уровня организованы различным образом. Кора левого полушария имеет более развитые связи со стволовыми структурами, тогда как правого – с диэнцефальными (Брагина Н. Н., Доброхотова Т. А., 1988)
Полученные результаты также могут свидетельствовать о том, что влияния из ствола и мозжечка в некоторой степени нивелируют собственную ФМПА полушарий; такое заключение согласуется с представлениями о генерализованном преобладании ингибиторных процессов в левом полушарии, а активационных – в правом.
3.7.5.Промежуточный мозг
Кпромежуточному мозгу относятся:
1)зрительный мозг (thalamencephalon);
2)подбугровая (субталамическая) часть, гипоталамус (hypothalamus).
В зрительном мозге различают:
а) зрительный бугор (thalamus opticus),
б) надбугровую область (epithalamus), в) забугровую область (metathalamus).
3.7.5.1. Таламус
Зрительный бугор имеет яйцевидную форму. Задний полюс зрительного бугра утолщен и слегка вытянут, его называют подушкой (pulvinar thalamus). В зрительных буграх находятся многочисленные, свыше 120 ядерных образований, в которые поступает афферентация из различных отделов головного мозга (коры головного мозга, мозжечка, центров экстрапирамидной системы, миндалевидного комплекса).
Зрительный бугор - это коллектор всех видов чувствительности. Уолкер, один из специалистов по функциональной организации зрительных бугров, писал:
200
"Таламус является посредником, в котором сходятся все раздражения от внешнего мира и, видоизменяясь здесь, направляются к подкорковым и корковым центрам таким образом, чтобы организм смог адекватно приспособиться к постоянно меняющейся среде. Таламус, как видно, таит в себе тайну того, что происходит в коре головного мозга".
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер). Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая — в разные области коры большого мозга.
1.Предложено несколько классификаций ядер зрительных бугров, в основу которых положены различные принципы. Уолкер (1938) разделил ядра таламуса на шесть групп.
В 1927 году Хасслер на основании характера связей ядер зрительных бугров с корой головного мозга выделил две системы ядер:
1.Паллиоталамические ядра. К ним относятся ядра, имеющие двусторонние связи с корой головного мозга (передняя ядерная группа, медиальная ядерная группа, латеральная ядерная область, медиальные коленчатыетела, латеральные коленчатые тела, подушка).
2.Трункоталамические ядра. Эти ядра не имеют прямых связей с корой головного мозга (центральное, или перивентрикулярное, серое вещество таламуса, интраламинарная группа и ретикулярная зона).
В 1976 году Адрианов на основании эволюционных и физиологических признаков выделил три группы ядер:
1. Диффузная таламическая система (неспецифические
201
ядра). Данная группа ядер представляет филогенетически древнюю часть таламуса и состоит из интраламинарной группы ядер средней линии. Диффузная таламическая система характеризуется следующими признаками:
а) получает афференты из филогенетически древней экстралемнисковой системы, спинного мозга, бульбарных отделов ретикулярной формации;
б) не имеет прямого выхода к коре головного мозга. Выход в кору мозга осуществляется через ретикулярное ядро таламуса.
2.Группа специфических (релейных) ядер. Данная группа подразделяется на сенсорные и несенсорные релейные ядра.
Релейные ядра:
а) получают афференты от лемнисковых систем (спинальная, тригеминальная, слуховая, зрительная), от структур мозжечка, гипоталамуса, полосатого тела;
б) имеют прямой выход в кору головного мозга; в) характеризуются топографической и функциональной
разграниченностью проекций к определенным областям коры головного мозга;
г) имеют соматотопическую организацию.
3.Ассоциативные ядра таламуса. Данная группа ядер характеризуется следующими признаками:
а) не имеет прямого афферентного входа из периферии; б) афференты этих ядер берут начало в других ядрах
таламуса; в) имеется развитая система связей данной группы ядер с
ассоциативными полями коры головного мозга; г) данная группа ядер осуществляет наиболее сложные
интегративные процессы по сравнению с другими ядрами таламуса.
Как отмечено выше, таламус является коллектором всех видов чувствительности. Это объясняется тем, что в таламус входит 12 различных афферентных систем:
а) зрительная афферентная система; б) слуховая афферентная система; в) вкусовой тракт; г) спинно-таламический тракт; д) спинно-кортикальный тракт; е) спинно-цервико- таламический тракт; ж) дорсальный спинно-церебеллярный
тракт; з) вестибуло-таламический тракт; и) церебелло-
202
таламический тракт; к) пучок лентикулярной петли; л) внутренний лапидарный тракт; м) наружный палидарный тракт.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
Вмедиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Многочисленность афферентных входов зрительных бугров приводит к тому, что любой афферентный импульс, прежде чем поступит в кору головного мозга, непременно достигнет зрительных бугров. При этом следует подчеркнуть, что зрительные бугры не являются только проводниками афферентации в кору мозга, в зрительных буграх совершаются сложные интегративные процессы. Только после соответствующей трансформации на уровне зрительных бугров афферентация поступает в кору головного мозга.
Основной эфферентный выход из таламуса представлен системой волокон, которая проводит афферентацию из зрительных бугров в кору мозга.
Воснове интегративной деятельности таламуса лежит многоуровневый принцип организации интегративных процессов.
Первый уровень интеграции - это взаимодействие процессов на синаптическом уровне.
Второй уровень интеграции - это интрануклеарный уровень, выражающийся в объединении значительной группы нейронов таламического ядра с помощью внутриядерных
тормозных |
интернейронов. |
Тормозный |
интернейрон |