

Бактериофаги — вирусы,
избирательно поражающие бактериальные
клетки. Чаще всего бактериофаги
размножаются внутри бактерий и вызывают
их лизис.
Как правило, бактериофаг состоит из
белковой оболочки и генетического
материала.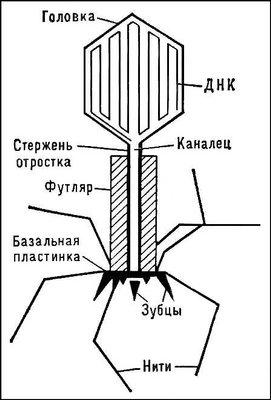
Типичная фаговая частица (вирион) состоит из головки и хвоста. Длина хвоста обычно в 2—4 раза больше диаметра головки. В головке содержится генетический материал — одноцепочечная или двуцепочечная РНК или ДНК с ферментом транскриптазой в неактивном состоянии, окружённая белковой или липопротеиновой оболочкой — капсидом. Нуклеиновая кислота и капсид вместе составляют нуклеокапсид. Хвост, или отросток, представляет собой белковую трубку — продолжение белковой оболочки головки, в основании хвоста имеется АТФаза, которая регенерирует энергию для инъекции генетического материала.
Головка округлой, гексагональной или палочковидной формы диаметром 45—140 нм. Отросток толщиной 10—40 и длиной 100—200 нм. Длина нити нуклеиновой кислоты во много раз превышает размер головки, в которой находится в скрученном состоянии. Отросток имеет вид полой трубки, окружённой чехлом, содержащим сократительные белки, подобные мышечным. У ряда вирусов чехол способен сокращаться, обнажая часть стержня. На конце отростка у многих бактериофагов имеется базальная пластинка, от которой отходят тонкие длинные нити, способствующие прикреплению фага к бактерии. Общее количество белка в частице фага — 50—60%, нуклеиновых кислот — 40—50%.
У фагов отсутствуют механизмы для выработки энергии и рибосомы для синтеза белка.
Вирулентные фаги могут только увеличиваться в количестве посредством литического цикла. Процесс взаимодействия вирулентного бактериофага с клеткой складывается из нескольких стадий: адсорбции бактериофага на клетке, проникновения в клетку, биосинтеза компонентов фага и их сборки, выхода бактериофагов из клетки.
Первоначально бактериофаги прикрепляются к фагоспецифическим рецепторам на поверхности бактериальной клетки. Хвост фага с помощью ферментов, находящихся на его конце (в основном лизоцима), локально растворяет оболочку клетки, сокращается и содержащаяся в головке ДНК инъецируется в клетку, при этом белковая оболочка бактериофага остаётся снаружи. Инъецированная ДНК вызывает полную перестройку метаболизма клетки: прекращается синтез бактериальной ДНК, РНК и белков. ДНК бактериофага начинает транскрибироваться с помощью собственного фермента транскриптазы, который после попадания в бактериальную клетку активируется. Синтезируются сначала ранние, а затем поздние иРНК, которые поступают на рибосомы клетки-хозяина, где синтезируются ранние (ДНК-полимеразы, нуклеазы) и поздние (белки капсида и хвостового отростка, ферменты лизоцим, АТФаза и транскриптаза) белки бактериофага. Репликация ДНК бактериофага происходит по полуконсервативному механизму и осуществляется с участием собственных ДНК-полимераз. После синтеза поздних белков и завершения репликации ДНК наступает заключительный процесс — созревание фаговых частиц или соединение фаговой ДНК с белком оболочки и образование зрелых инфекционных фаговых частиц. Продолжительность этого процесса может составлять от нескольких минут до нескольких часов. Затем происходит лизис клетки, и освобождаются новые зрелые бактериофаги.
Вирулентные бактериофаги развиваются по Литической модели:
- Нуклеиновая кислота фага направляет синтез ферментов фага, используя для этого белоксинтезирующий аппарат бактерии. Фаг тем или иным способом инактивирует ДНК и РНК хозяина, а ферменты фага совсем расщепляют её; РНК фага «подчиняет» себе клеточный аппарат синтеза белка.
- Нуклеиновая кислота фага реплицируется и направляет синтез новых белков оболочки. Образуются новые частицы фага в результате спонтанной самосборки белковой оболочки (капсид) вокруг фаговой нуклеиновой кислоты; под контролем РНК фага синтезируется лизоцим.
- Лизис клетки: клетка лопается под воздействием лизоцима; высвобождается около 200—1000 новых фагов; фаги инфицируют другие бактерии.
Умеренные фаги вызывают латентную инфекцию и ассоциацию генома вируса с бактериальной хромосомой. Умеренные фаги, в отличие от вирулентности, не вызывают гибели бактериальных клеток и при взаимодействии с ней переходят в неинфекционную форму фага, называемую профагом. Профаг— геном фага, ассоциированный с бактериальной хромосомой. Профаг, ставший частью хромосомы клетки, при ее размножении реплицируется синхронно с геномом бактерии, не вызывая ее лизиса, и передается по наследству от клетки к клетке в неограниченном числе поколений. Бактериальные клетки, содержащие в своей хромосоме профаг, называются лизогенными. Профаг в лизогенных бактериях самопроизвольно или под влиянием различных индуцированных агентов может переходить в вегетативный фаг. В результате такого превращения бактериальная клетка лизируется и продуцирует новые фаговые частицы. В ходе лизогенизации бактериальные клетки могут дополнительно приобретать новые признаки, детерминируемые геномом вируса. Такое явление — изменение свойств микроорганизмов под влиянием профага — называется фаговой, или лизогенной, конверсией (проявление вирус-индуцированной трансформации).
Умеренные фаги, неспособные ни при каких условиях переходить из профага в вегетативный фаг (образовывать зрелые фаговые частицы), называются дефектными, чаще это происходит в результате нарушения стадии сборки вирусных частиц. Некоторые умеренные фаги называются трансдуцирующими,поскольку с их помощью осуществляется один из механизмов генетической рекомбинации у бактерий - трансдукции.
Взаимодействие фагов с клеткой (бактерией) строго специфично, т.е. бактериофаги способны инфицировать только определенные виды и фаготипы бактерий.
Основные этапы взаимодействия фагов и бактерий.
1.Адсорбция (взаимодействие специфических рецепторов).
2.Внедрение вирусной ДНК (инъекция фага) осуществляется за счет лизирования веществами типа лизоцима участка клеточной стенки, сокращения чехла, вталкивания стержня хвоста через цитоплазматическую мембрану в клетку, впрыскивание ДНК в цитоплазму.
3.Репродукция фага.
4.Выход дочерних популяций.
Очень важным свойством бактериофагов является их специфичность: бактериофаги лизируют культуры определённого вида, более того, существуют так называемые типовые бактериофаги, лизирующие варианты внутри вида, хотя встречаются поливалентные бактериофаги, которые паразитируют в бактериях разных видов.
Жизненный цикл
Умеренные и вирулентные бактериофаги на начальных этапах взаимодействия с бактериальной клеткой имеют одинаковый цикл.
-
Фаг приближается к бактерии, и хвостовые нити связываются с рецепторными участками на поверхности бактериальной клетки.
-
Хвостовые нити изгибаются и «заякоривают» шипы и базальную пластинку на поверхности клетки; хвостовой чехол сокращается, заставляя полый стержень входить в клетку; этому способствует фермент лизоцим, который находится в базальной пластинке; таким образом нуклеиновая кислота (ДНК или РНК) вводится внутрь клетки.
-
Нуклеиновая кислота фага кодирует синтез ферментов фага, используя для этого белоксинтезирующий аппарат хозяина.
-
Фаг тем или иным способом инактивирует ДНК и РНК хозяина, а ферменты фага совсем расщепляют её; РНК фага подчиняет себе клеточный аппарат.
-
Нуклеиновая кислота фага реплицируется и кодирует синтез новых белков оболочки.
-
Новые частицы фага, образовавшиеся в результате спонтаной самосборки белковой оболочки вокруг фаговой нуклеиновой кислоты; под контролем РНК фага синтезируется лизоцим.
-
Лизис клетки: клетка лопается под воздействием лизоцима; высвобождается около 200-1000 новых фагов; фаги инфицируют другие бактерии.
-
Стадии 1-7 по времени занимают около 30 минут; этот период называется латентным периодом.
-
Умеренные бактериофаги после деления клетки находятся в состоянии профага (Лизогенный путь).
Одной из областей использования бактериофагов является антибактериальная терапия, альтернативная приёму антибиотиков. Бактериофаги применяются в генной инженерии в качестве векторов, переносящих участки ДНК, возможна также естественная передача генов между бактериями посредством некоторых фагов (трансдукция). Поскольку размножение бактериофага возможно только в живых клетках, бактериофаги могут быть использованы для определения жизнеспособности бактерий.
По признаку специфичности выделяют поливалентные бактериофаги, лизирующие культуры одного семейства или рода бактерий, моновалентные (монофаги) - лизирующие культуры только одного вида бактерий, а также отличающиеся наиболее высокой специфичностью - типовые бактериофаги, способные вызывать лизис только определенных типов (вариантов) бактериальной культуры внутри вида бактерий.
ПРЕИМУЩЕСТВА БАКТЕРИОФАГОВ ПЕРЕД АНТИБИОТИКАМИ:
-
бактериофаги способны уничтожать бактерии, устойчивые к антибиотикам, т.к. они действуют лишь на определенные бактерии;
-
свободно проникают в ткани организма человека и животного не нарушая баланса высшего организма;
-
постоянно эволюционируют;
-
не вызывают побочных эффектов;
-
не подавляют рост нормофлоры, не ослабляют иммунитет;
-
не развивают устойчивость (привыкание) бактерий;
-
сочетаются с любыми лекарственными препаратами, оказывают положительное действие на становление иммунитета;
-
не подавляют и не нарушают действия человеческого организма.
Практическое использование бактериофагов.
1.Для идентификации (определение фаготипа). Фагодиагностика:
а) фагоиндикация (метод индикации (обнаружения) патогенных бактерий в исследуемом материале при помощи специальных (индикаторных) фагов. РНТФ (реакция нарастания титра фага)
Компоненты:
- исследуемый материал
- определенное количество частиц индикаторного фага
Инкубируют, фильтруют через бактериальный фильтр и в фильтрате подсчитывают количество фаговых частиц. Увеличение числа фаговых частиц свидетельствует о присутствии в материале соответствующих бактерий. Подсчет фаговых частиц в фильтрате проводят методом титрования по Грациа.