

Звягина Учебное пособие
.pdfРисунок 16. Схема распада пуриновых нуклеотидов до мочевой кислоты
131
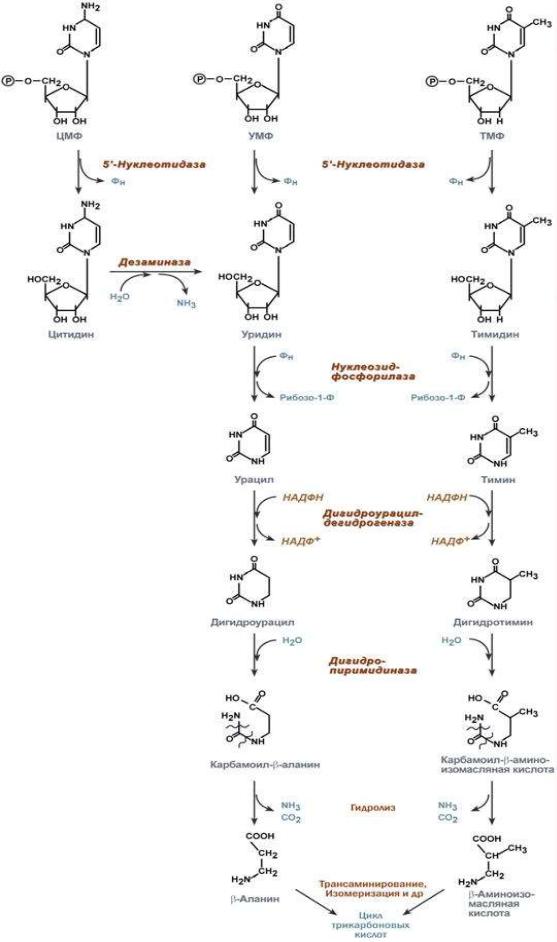
Рисунок 17. Схема распада пиримидиновых нуклеотидов до конечных продуктов
132

Другое заболевание, при котором кристаллы уратов откладываются в почечной лоханке или в мочевом пузыре, известно как мочека-
менная болезнь.
Для лечения подагры и мочекаменной болезни применяются:
ингибиторы фермента ксантиноксидазы. Например, аллопуринол
– вещество пуриновой природы, является конкурентным ингибитором фермента. Действие этого препарата приводит к повышению концентрации гипоксантина. Гипоксантин и его соли лучше растворимы в воде, и легче выводятся из организма.
диетическое питание, исключающее продукты, богатые нуклеиновыми кислотами, пуринами и их аналогами: икра рыб, печень, мясо, кофе и чай.
соли лития, поскольку они лучше растворимы в воде, чем ураты натрия.
Катаболизм пиримидиновых азотистых оснований
Конечными продуктами пиримидинового обмена являются β- аланин и β-аминоизомасляная кислота (аминоизобутират) (рис.17). β- аланин содержится в плазме крови и многих других тканях, он используется в мышцах на образование дипептидов: карнозина и анзерина, часть его может окисляться до СО2 и Н2О.
Синтез пуриновых мононуклеотидов
Для синтеза мононуклеотидов de novo необходимы очень простые вещества: CO2 и рибозо-5-фосфат. Синтез происходит с затратой АТФ. Кроме этого, необходимы заменимые аминокислоты, которые синтезируются в организме, поэтому даже при полном голодании синтез нуклеиновых кислот не страдает.
Особенностью синтеза пуриновых нуклеотидов является то, что циклическая структура пуринового азотистого основания постепенно достраивается на активной форме рибозо-фосфата, как на матрице. При циклизации получается уже готовый пуриновый мононуклеотид.
Пуриновое кольцо строится из СО2, аспарагиновой кислоты, глутамина, глицина и серина. Эти вещества либо полностью включаются
133

в пуриновую структуру, или передают для ее построения отдельные группировки.
5-фосфорибозиламин вовлекается в цепь последовательных реакций, и в результате образуется первый пуриновый нуклеотид – инозинмонофосфорная кислота (ИМФ).
Рисунок 18. Схема синтеза пуриновых нуклеотидов
134

АМФ и ГМФ превращаются далее в нуклеозиддифосфаты с помощью нуклеозидмонофосфаткиназ, функция этих ферментов заключается в том, чтобы поддерживать равновесие фонда нуклеотидов.
ГДФ далее превращается в ГТФ при участии фермента нуклеозиддифосфаткиназы и молекулы АТФ. Превращение АДФ в АТФ происходит, в основном, за счет окислительного фосфорилирования.
В некоторых тканях есть альтернативный способ синтеза – реутилизация (повторное использование) пуриновых азотистых оснований, которые образовались при распаде нуклеотидов.
Ферменты, катализирующие реакции реутилизации, наиболее активны в быстроделящихся клетках (эмбриональные ткани, красный костный мозг, раковые клетки), а также в тканях головного мозга. На схеме видно, что фермент гуанингипоксантинФРПФтрансфераза
обладает более широкой субстратной специфичностью, чем аденинФРПФтрансфераза – помимо гуанина, может переносить и гипоксантин - образуется ИМФ. У человека встречается генетический дефект этого фермента – «болезнь Леша-Нихана». Для таких больных характерны выраженные морфологические изменения в головном и костном мозге, умственная и физическая отсталость, агрессия, аутоагрессия.
135

Синтез пиримидиновых мононуклеотидов.
В отличие от пуринов, при биосинтезе пиримидинов de novo вначале образуется пиримидиновое кольцо, а лишь затем к нему присоединяется рибозо-5-фосфат. Источниками атомов для пиримидинового кольца являются глутамин, аспартат и СО2. Синтез начинается с образования карбамоилфосфата:
Глутамин + СО2 + 2 АТФ Карбамоилфосфат + Глутамат + 2
КарбамоилфосфатсинтетазаI
АДФ + Фн.
Из мононуклеотидов посредством фосфаткиназ синтезируются ди- и трифосфаты
ЦТФ образуется из УТФ в следующей рекции:
136
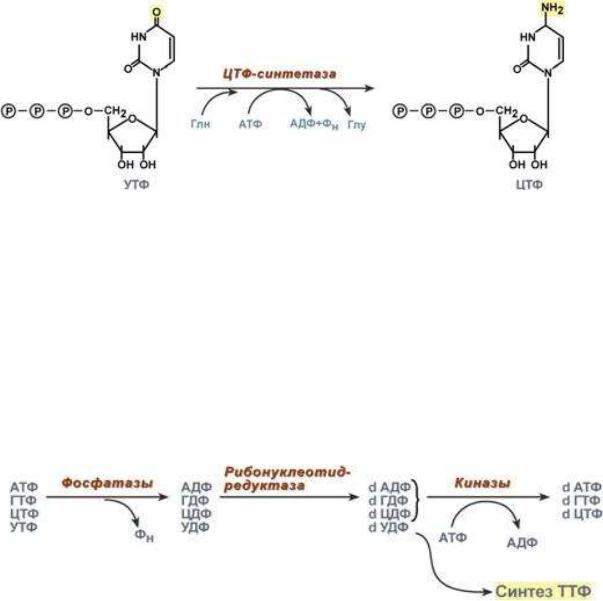
дТМФ синтезируется из дезоксиформы УМФ Ключевой фермент синтеза пиримидиновых нуклеотидов — ас-
партаткарбамоилтрансфераза.
Регуляция: избыток пиримидиновых нуклеотидов ингибирует ключевой фермент, а избыток пуриновых — активирует.
Образование дезоксирибонуклеотидов
Особенностью обмена пуринов и пиримидинов является то, что они могут образовывать не только рибонуклеотиды, но и дезоксирибонуклеотиды, которые необходимы клетке для синтеза ДНК.
В самом начале процесса происходит потеря рибонуклеозидтрифосфатами одной фосфатной группы и образуются АДФ, ГДФ, ЦДФ, УДФ.
Во второй реакции фермент рибонуклеозид-редуктаза восстанавливает АДФ, ГДФ, ЦДФ, УДФ до дезоксирибонуклеозиддифосфатов dАДФ, dГДФ, dЦДФ, dУДФ. Донором водорода для восстановления рибозы является белок тиоредоксин, его SH-группы окисляются кислородом рибозы и образуется вода. Последующее восстановление тиоредоксина в рабочее состояние обеспечивается за счет НАДФН2.
137

После образования dАДФ, dГДФ, dЦДФ фосфорилируются, а dУДФ используется для синтеза тимидилового нуклеотида.
Обмен гемпротеинов. Распад гемоглобина. Образование билирубина и других желчных пигментов. Обезвреживание билирубина. Понятие о свободном («непрямом») и связанном («прямом») билирубине.
Распад гемоглобина происходит в селезенке и печени. Сначала разрывается связь между I-м и II-м кольцами под действием фермента - оксигеназы. После ее действия образуется пигмент зеленого цвета - вердоглобин. В его составе сохраняется и железо и белок. В дальнейшем вердоглобин распадается на составные части: белок, железо и пигмент-биливердин. Железо образует комплекс с белком трансферрином, и этот комплекс поступает в ткани, где оно снова может быть использовано для синтезов или депонироваться в виде комплекса с белком ферритином.
138
Билирубин плохо |
растворим в воде, поэтому он адсорбируется |
на альбумине крови. |
Такой комплекс называется свободным или не- |
прямым билирубином. Он свободный, потому что не связан ни с каким другим веществом ковалентными связями; непрямой, потому что не дает прямой реакции с диазореактивом Эрлиха (для определения этого вида билирубина необходимо предварительно осадить альбумины).
Впечени непрямой билирубин подвергается обезвреживанию путем связывания с глюкуроновой кислотой. В реакции расходуется УТФ. При этом к молекуле билирубина присоединяются 2 молекулы глюкуроновой кислоты (образование билирубиндиглюкуронида) под действием фермента - УДФ - глукуронозилтрансферазы. Это веще-
ство малотоксично и хорошо растворяется в воде, называется прямым связанным билирубином, т.к. определяется в прямой реакции с диазореактивом и, ковалентно связан с глюкуроновой кислотой. В таком виде билирубин поступает в желчь и вместе с ней выводится из организма.
При определении содержания билирубина используют диазореактив, который дает розовую окраску с билирубином и по ее интенсивности судят о концентрации билирубина в крови.
Вкрови человека примерно 75% непрямого билирубина от его общего количества. Общее содержание билирубина в норме (прямой+непрямой) от 8 до 20 мкмоль/л.
Билирубин - это конечный продукт распада гемоглобина у человека. Однако, поступив с желчью в кишечник, он превращается под действием ферментов микрофлоры кишечника. Сначала от него отщепляется глюкуроновая кислота, затем происходит частичное восстановление и образуется вещество, называемое мезобилиноген, а затем стеркобилиноген. Стеркобилиноген в основном выводится с калом и на воздухе (в кале) превращается в стеркобилин, а мезобилиноген частично всасывается из кишечника по системе v.portae, т.е. попадает в печень и там разрушается. Поэтому его нет у здоровых людей в крови. Небольшая часть стеркобилиногена тоже всасывается из кишечника через систему геморроидальных вен, т.е. попадает сразу в большой круг кровообращения, минуя печень, поэтому часть его выводится с мочой.
Значит, стеркобилиноген является компонентом мочи в норме.
ВНОРМЕ:
Кровь:
139
Билирубин (в основном непрямой) - в норме (8-20 мкмоль/л) Гемоглобин - в норме (мужчины - 130-145 г/л; женщины - 110-130 г/л Моча: светло-желтого цвета, стеркобилиноген - в норме, мезобилиногена нет.
Кал: нормальной окраски, стеркобилиноген - в норме
Нарушения обмена билирубина. Диагностическое значение определения билирубина в крови и моче.
При некоторых заболеваниях билирубин начинает накапливаться в слизистых оболочках, коже, склерах. Человек «желтеет». Этот симптом называется желтуха и наблюдается при желчнокаменной болезни, гемолизе, инфекционном или токсическом гепатитах. Определение билирубина в моче, в крови и в кале позволяет выявить причины желтухи.
Гемолитическая желтуха. Наблюдается при усилении распада эритроцитов. Билирубина образуется больше, чем в норме и скорость экскреции тоже увеличивается. Непрямой билирубин повышается в крови. В моче билирубина нет, а содержание стеркобилина в кале и в моче повышено.
Печеночная желтуха (паренхиматозная). Наблюдается при повреждении гепатоцитов (например, при вирусном гепатите). Билирубин не обезвреживается. Желчные пигменты поступают не только в кишечник, но и в кровь. Т.е. в крови повышается содержание непрямого билирубина.
Обтурационная желтуха (механическая). Наблюдается при закупорке желчных протоков (например, при желчнокаменной болезни). Желчь продолжает вырабатываться, но поступает не только в желчные протоки, а также в кровь. В крови повышается уровень билирубина, в основном - прямого билирубина. Наблюдается билирубинурия. Содержание стеркобилина в моче и в кале снижено (кал становится светлым, а моча темнеет за счет прямого билирубина).
I.Вопросы для самоподготовки:
1.Фонд свободных аминокислот.
2.Классификация протеолитических ферментов.
3.Функции и регуляция активности тканевых протеиназ.
4.Дезаминирование аминокислот, его виды.
5.Трансдезаминирование и прямое окислительное дезаминирование аминокислот, биологическая роль этих процессов.
140