

otvety__tablitsa_Vlada
.pdf3. 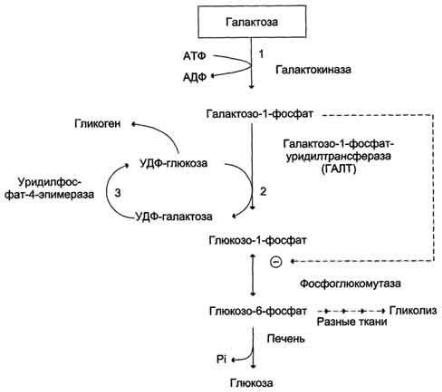
Нарушения обмена галактозы
Дефектный фермент |
Блокируемая |
Клинические проявления |
(частота) |
реакция |
и лабораторные данные |
|
|
|
Галактокиназа |
Галактоза + АТФ → |
Галактоземия, галактозурия, |
(1:500 000) |
Галактозо-1-фосфат + АДФ |
катаракта. Активность |
|
|
фермента в эритроцитах |
|
|
нормальная. |
|
|
|
Галактозо-1- |
Галактозо-1-фосфат + УДФ- |
Галактоземия, галактозурия, |
фосфатуридилтрансфераза |
глюкоза → УДФ-галактоза + |
галактозо-1-фосфатемия, |
(1:40000) |
Глюкозо-1-фосфат |
катаракта. Тенденция к |
|
|
гипогликемии, ком- |
|
|
пенсаторная мобилизация |
|
|
жиров, цирроз печени, |
|
|
нарушения функции почек. |
|
|
Гепатомегалия, задержка |
|
|
психического развития. |
|
|
Активность фермента в |
|
|
эритроцитах снижена. |
|
|
|
Уридилфосфат-4-эпимераза |
УДФ-глюкоза ↔ УДФ- |
Галактоземия, галактозурия. |
(1:1000000) |
галактоза |
Тяжѐлых клинических |
|
|
|

проявлений нет. Описаны единичные случаи заболевания.
368
Таблица 7-7. Некоторые варианты генетического дефекта ГАЛТ
Изменения в структуре |
Проявления |
ГАЛТ |
|
|
|
Асн→Асп |
Признак Дюарта. У гетерозигот при этом варианте |
|
активность фермента составляет 75% от нормальной. |
|
Гомозиготный фенотип Дюарта обычно связан с 50% |
|
потерей активности. Пациенты с синдромом Дюарта |
|
могут быть здоровыми, несмотря на структурную |
|
аномалию ГАЛТ. |
|
|
Глн→Арг |
Проявляется как тяжѐлая галактоземия. Причина - |
|
мутация типа замены нуклеотида 591 в гене фермента. |
|
Активность ГАЛТ составляет 10% от нормы. Эта форма |
|
встречается в 70% случаев заболевания галактоземией |
|
среди европеоидов, частота - 1 :338 886. |
|
|
Сер→Лей |
Заболевание описано у чернокожих пациентов и |
|
названо «чѐрный признак». Галактоземия проявляется |
|
как результат недостаточной активности ГАЛТ в |
|
печени и эритроцитах. Активность ГАЛТ в печени |
|
составляет 10% от нормы. Тем не менее отмечалась |
|
утилизация некоторого количества галактозы, что |
|
объяснялось развитием альтернативного пути. Причина |
|
- мутация типа замены 1 158-го нуклеотида в гене |
|
фермента. |
|
|
Арг→Три |
Тяжѐлая форма галактоземии. Причина - миссенс- |
|
мутация нуклеотида 1025 в гене фермента. Активность |
|
ГАЛТ отсутствует. |
|
|
Лиз→Асн |
Широко распространѐнная мутация при галактоземии. |
|
|
Билет 9, вопрос 2.
Глюконеогенез, синтез глюкозы из глицерина, химизм реакций, значение, регуляция.
Глюконеогенез - процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. Процесс протекает в основном в печени и менее интенсивно в корковом веществе почек, а также в слизистой оболочке кишечника. Эти ткани могут обеспечивать синтез 80-100 г глюкозы в сутки.
Первичные субстраты глюконеогенеза - лактат, аминокислоты и глицерол. Включение этих субстратов в глюконеогенез зависит от физиологического состояния организма.
Лактат - продукт анаэробного гликолиза. Он образуется при любых состояниях организма в эритроцитах и работающих мышцах. Таким образом, лактат используется в глюконеогенезе постоянно.
Глицерол высвобождается при гидролизе жиров в жировой ткани в период голодания или при длительной физической нагрузке.
Аминокислоты образуются в результате распада мышечных белков и включаются в глюконеогенез при длительном голодании или продолжительной мышечной работе.
Большинство реакций глюконеогенеза протекает за счѐт обратимых реакций гликолиза (реакции 9, 8, 7, 6, 5, 4, 2) и катализируется теми же ферментами. Однако 3 реакции гликолиза (реакции 1, 3, 10) термодинамически необратимы. На этих стадиях реакции глюконеогенеза протекают другими путями.
Необходимо отметить, что гликолиз протекает в цитозоле, а часть реакций глюконеогенеза происходит в митохондриях.
Регуляция глюконеогенеза
Гормональная активация глюконеогенеза осуществляется глюкокортикоидами, которые увеличивают синтез пируваткарбоксилазы, фосфоенолпируват-карбоксикиназы, фруктозо- 1,6-дифосфатазы. Глюкагон стимулирует те же самые ферменты через аденилатциклазный механизм путем фосфорилирования.
Также имеется метаболическая регуляция, при которой аллостерически активируется пируваткарбоксилаза при помощи ацетил-SКоА, фруктозо-1,6-дифосфатаза при участии АТФ.

Ферменты обратимых реакций гликолиза и глюконеогенеза: 2 - фосфоглюкоизомераза; 4 - альдолаза; 5 - триозофосфатизомераза; 6 - глицеральдегидфосфатдегидрогеназа; 7 - фосфоглицераткиназа; 8 - фосфоглицератмутаза; 9 - енолаза. Ферменты необратимых реакций глюконеогенеза: 11 - пируваткарбоксилаза; 12 - фосфоенолпируваткарбоксикиназа; 13 - фруктозо-1,6-бисфосфатаза; 14 -глюкозо-6- фосфатаза. I-III -субстратные циклы.
Рассмотрим более подробно те реакции глюконеогенеза, которые отличаются от реакций гликолиза и происходят в глюконеогенезе с использованием других ферментов. Рассмотрим процесс синтеза глюкозы из пирувата.
1. Образование фосфоенолпирувата из пирувата - первая из необратимых стадий глюконеогенеза
Образование фосфоенолпирувата из пирувата происходит в ходе двух реакций (рис. выше, реакции 11, 12), первая из которых протекает в митохондриях. Пируват, образующийся из лактата или из некоторых аминокислот, транспортируется в матрикс митохондрий и там карбоксилируется с образованием оксалоацетата.
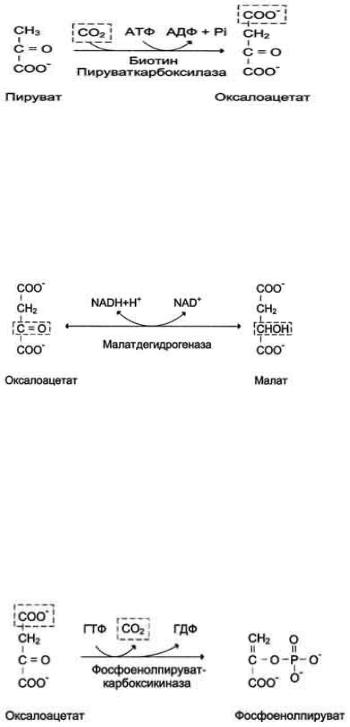
Пируват-карбоксилаза, катализирующая данную реакцию, - митохондриальный фермент, коферментом которого является биотин. Реакция протекает с использованием АТФ.
Дальнейшие превращения оксалоацетата протекают в цитозоле. Следовательно, на этом этапе должна существовать система транспорта оксалоацетата через митохондриальную мембрану, которая для него непроницаема. Оксалоацетат в митохондриальном матриксе восстанавливается с образованием малата при участии NADH (обратная реакция цитратного цикла).
Образовавшийся малат затем проходит через митохондриальную мембрану с помощью специальных переносчиков. Кроме того, оксалоацетат способен транспортироваться из митохондрий в цитозоль в виде аспартата в ходе малат-аспартатного челночного механизма.
В цитозоле малат вновь превращается в оксалоацетат в ходе реакции окисления с участием кофермента NAD+. Обе реакции: восстановление оксалоацетата и окисление малага катализируют малатдегидрогеназа, но в первом случае это митохондриальный фермент, а во втором - цитозольный. Образованный в цитозоле из ма-лата оксалоацетат затем превращается в фосфоенолпируват в ходе реакции, катализируемой фосфоенолпируваткарбоксикиназой - ГТФ-зависимым ферментом.
2. Гидролиз фруктозо-1,6-бисфосфата и глюкоза-6-фосфата
Отщепление фосфатной группы из фруктозо-1,6-бисфосфата и глюкозо-6-фосфата - также необратимые реакции глюконеогенеза. В ходе гликолиза эти реакции катализируют специфические киназы с использованием энергии АТФ. В глюконеогенезе они протекают без участия АТФ и АДФ и ускоряются не киназами, а фосфатазами - ферментами, принадлежащими к классу гидролаз. Ферменты фруктозо-1,6-бисфосфатаза и глюкозо-6- фосфатаза катализируют отщепление фосфатной группы от фруктозо-1,6-бисфосфата и глюкозо-6-фосфата. После чего свободная глюкоза выходит из клетки в кровь.
Синтез глюкозы из глицерола
Глицерол образуется при гидролизе триацил-глицеролов, главным образом в жировой ткани. Использовать его могут только те ткани, в которых имеется фермент глицеролкиназа, например печень, почки. Этот АТФ-зависимый фермент катализирует превращение глицерола в α-глицерофосфат (глицерол-3-фосфат). При включении глицерол-3-фосфата в глюконеогенез происходит его дегидрирование NAD-зависимой дегидрогеназой с образованием дигидроксиацетонфосфата, который далее превращается в глюкозу.
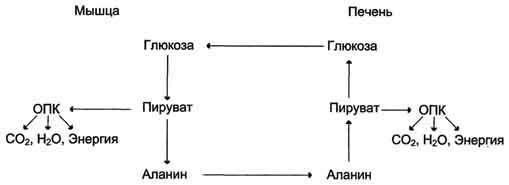
Глюкозо-аланиновый цикл
Билет 10. Вопрос 2.
Синтез глюкозы из лактата и цикл Кори. (и процесс глюконеогенеза – все то же самое, что в предыдущем вопросе, кроме синтеза глюкозы из глицерола).
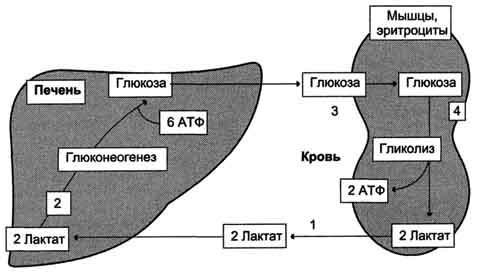
Лактат, образованный в анаэробном гликолизе, не является конечным продуктом метаболизма. Использование лактата связано с его превращением в печени в пируват. Лактат как источник пирувата важен не столько при голодании, сколько при нормальной жизнедеятельности организма. Его превращение в пируват и дальнейшее использование последнего являются способом утилизации лактата.
Лактат, образовавшийся в интенсивно работающих мышцах или в клетках с преобладающим анаэробным способом катаболизма глюкозы, поступает в кровь, а затем в печень. В печени отношение NADH/NAD+ ниже, чем в сокращающейся мышце, поэтому лактатдегидрогеназная реакция протекает в обратном направлении, т.е. в сторону образования пирувата из лактата. Далее пируват включается в глюконеогенез, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами. Эту последовательность событий называют "глюкозо-лактатным циклом", или "циклом Кори" (рис. ниже). Цикл Кори выполняет 2 важнейшие функции: 1 - обеспечивает утилизацию лактата; 2 - предотвращает накопление лактата и, как следствие этого, опасное снижение рН (лактоацидоз). Часть пирувата, образованного из лактата, окисляется печенью до СО2 и Н2О. Энергия окисления может использоваться для синтеза АТФ, необходимого для реакций глюконеогенеза.
11-- 4(1) 1(2) 4(3)
12-- 5(1) 2(2) 5(3)
13--6(1) 3(2) 6(3)
14--7(1) 4(2) 7(3)
15--8(1) 5(2) 8(3)
16--9(1) 6(2) 9(3)
17--10(1) 7(2) 10(3)
18--4(1) 8(2) 4(3)
19--5(1) 9(2) 5(3)
20--6(1) 10(2) 6(3)
21--7(1) 1(2) 7(3)
22--8(1) 2(2) 8(3)
23--9(1) 3(2) 9(3)
24--10(1) 4(2) 10(3)
25--4(1) 5(2) 4(3)
26--5(1) 6(2) 5(3)
27--6(1) 7(2) 6(3)
28--7(1) 8(2) 7(3)
29--8(1) 9(2) 4(3)
30--4(1) 10(2) 7(3)