

юрин физиология растений
.pdfÎäíè èç íèõ (ðèñ. 4.9, 1, 2) связаны с недостатком СО2, который может быть вызван фотосинтезом. Снижение межклеточной концентрации СО2 служит сигналом для открытия устьиц. Вода определяет другой тип связей (рис. 4.9, 3), при ее недостатке в тканях устьица закрываются. Однако оптимизация газового обмена достигается прямыми связями — действием света (рис. 4.9, 5), влажности и ветра (рис. 4.9, 6) и положительной связи фотосинтеза (рис. 4.9, 4) и обеспеченности водой эпидермальных клеток
è(или) клеток мезофилла (рис. 4.9, 3) в значительной мере обусловлено синтезом АБК, а положительная обратная связь (рис. 4.9, 4) определяется продуктами фотосинтеза в клетках мезофилла.
Внеустьичная регуляция транспирации представлена несколькими механизмами. Первый связан с обезвоживанием клеточных стенок, с поверхности которых идет испарение. Это механизм подсушивания. Когда поверхность верхних клеток мезофилла (хлоренхима) начинает подсыхать, то затрудняется движение воды и, таким образом, затрудняется и испарение. В клеточной стенке между микро- и макрофибриллами целлюлозы находятся капиллярные промежутки. Когда много воды и мениски в капиллярах выпуклые, силы поверхностного натяжения малы, то испарение идет быстро. Когда воды мало в клеточной стенке, мениски вогнуты, интенсивность испарения уменьшается.
Второй механизм связан со свойством протоплазмы изменять свою водоудерживающую способность.
На его существование указывает то обстоятельство, что при одной и той же степени открытия устьиц транспирация может сильно меняться. Так, например, показано, что коротковолновые лучи (сине- и ультрафиолетовые) немедленно увеличивают транспирацию за счет изменения структуры белковых. При частичной денатурации способность белков удерживать воду ослабляется и кутикулярная транспирация возрастает.
Во внеустьичной регуляции транспирации, возможно, участвуют эктодермы. Удлиняясь или укорачиваясь, они усиливают или уменьшают контакт клеток эпидермы с окружающей средой
èтем самым могут влиять на скорость испарения воды. Толщина кутикулы на поверхности листьев влияет на интен-
сивность транспирации. Она может изменяться под влиянием внешних факторов. Увеличение толщины кутикулы уменьшает интенсивность транспирации с поверхности листьев.
242
У некоторых растений существуют специфические механизмы внеустьичной регуляции транспирации. Так, например, у некоторых сортов сахарного тростника, когда транспирация превышает поступление воды, происходит свертывание листовых пластинок вдоль средней жилки в трубку и наблюдается снижение транспирации на 10—20 %.
При нехватке воды растения могут терять листья. Опять же, у сахарного тростника может остаться на побеге в условиях засухи один лист. При возобновлении дождей или полива количество листьев быстро увеличивается.
При наступлении зимней засухи в умеренной зоне деревья сбрасывают все листья. Однако листопадные формы деревьев встречаются в тропиках и пустынях.
Существует связь между устьичной и внеустьичной транспирацией. Как при максимально открытых устьицах наиболее эффективен внеустьичный регулятор, так и при закрытых устьицах уменьшение кутикулярной транспирации происходит за счет внеустьичной транспирации.
Роль регуляторных механизмов у растений разных видов неодинакова. Например, более низкая интенсивность транспирации, характерная для оливковых деревьев, связана с устьичным механизмом.
Факторы среды влияют непосредственно на интенсивность транспирации.
Главным фактором, регулирующим транспирацию, является ñâåò. С интенсивностью солнечной радиации хорошо коррелируют температура и влажность среды. Во время максимальной освещенности наблюдается и максимальная интенсивность транспирации. Ночью транспирация в 10 раз меньше, чем днем. Солнеч- ная радиация служит источником энергии, затрачиваемой на транспирацию.
Лист, поглощая свет, использует на фотосинтез 1—2 %, максимум 5 %, а остальная энергия тратится на испарение воды. Это прямое действие света на транспирацию. Однако свет играет и иную роль, как мы уже отмечали, влияя на открытие и закрытие устьиц.
Как физический процесс (испарение) транспирация зависит от дефицита насыщения воздуха водяными парами, температуры, ветра, величины испаряющей поверхности и др.
243
Потеря воды в условиях, когда солнечная радиация отсутствует, как раз и обусловлена дефицитом насыщения воздуха водяными парами.
Интенсивность транспирации в какой-то мере зависит от наличия ветра. Появление ветра приводит к увеличению транспирации. Однако прямой зависимости между скоростью ветра и величиной транспирации не наблюдается, так как испарение происходит из межклетников, защищаемых от ветра.
Интенсивность транспирации зависит от условий минерального питания. У растений при недостатке азота, фосфора и калия интенсивность транспирации максимальна. Почти такая же интенсивность у растений, получивших калий и фосфор, но при дефиците азота.
Резко уменьшается транспирация при полных сбалансированных минеральных удобрениях. Чем лучше питание, тем ниже транспирация. Увеличение транспирации может свидетельствовать о нарушении минерального питания.
При изменении солнечной радиации, температуры, влажности воздуха дневной ход интенсивности транспирации выглядит следующим образом: слабая в утренние часы транспирация быстро увеличивается по мере восхода солнца, увеличения температуры и уменьшения влажности воздуха, достигает максимума около полудня, затем быстро падает к заходу солнца.
Однако такой дневной ход наблюдается не всегда. В некоторых случаях, несмотря на интенсивную освещенность и высокую температуру, в полдень транспирация уменьшается, и отмечаются двухвершинные кривые. Нужно отметить, что кривые дневного хода транспирации очень разнообразны, что обусловлено тремя факторами: внешние условия данного дня, условия предыдущих дней и наследственность. Взаимодействие этих факторов и определяет реальную интенсивность транспирации.
Растения выработали в процессе эволюции различные приспособления для уменьшения транспирации: восковой налет на поверхности листьев и плодов, погружение устьиц в мезофилл, развитие волосков (опушенность), редукция листовой поверхности.
Сильно уменьшать транспирацию нельзя, так как она определяет подъем воды по растению, поддерживает постоянной температуру растения и т. д.
Взаимосвязь фотосинтеза и водного обмена растений можно представить схемой (рис. 4.10).
244
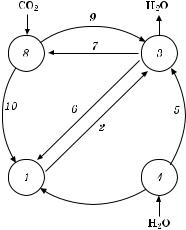
Ðèñ. 4.10. Схема взаимосвязи фотосинтеза и водного обмена растений:
1 — корневое давление; 2 — активное нагнетание воды; 3 — транспирация; 4 — осмотическое поглощение воды клетками корня; 5 — передвижение воды под действием транспирации; 6 — влияние транспирации на корневое давление; 7 — влияние
транспирации на фотосинтез; 8 — фотосинтез; 9 — влияние фотосинтеза на транспирацию; 10 — поступление ассимилятов в корень
Таким образом, водный обмен растений, определяющийся поступлением и расходованием воды, связан с различными физиологическими процессами, особенно с фотосинтезом (рис. 4.10).
4.4. Поступление воды в корень и перемещение по растению
Для нормального функционирования клетки растения должны быть насыщены водой. Состояние насыщения поддерживается с помощью двух взаимосвязанных процессов: поступления и выделения (расходования) воды, которые и составляют водный обмен растений. Соотношение между этими процессами называют водным балансом.
Растение выделяет воду в жидком и парообразном состояниях. Физиологический процесс испарения воды наземными органами
245
растений получил, как уже отмечалось, название транспирации, выделение воды в капельно-жидком состоянии — гуттации.
Жизнь возникла в Мировом океане. С выходом растений на сушу появилась опасность их гибели от высыхания. Ведь концентрация водяных паров в воздухе, даже в условиях влажного климата, всегда меньше, чем в межклетниках листа, поэтому неизбежно происходит движение воды из тела растения в окружающую среду, при этом диффузия молекул воды от поверхности листа в воздух в 1500 раз быстрее, чем их поступление из почвы в корень.
Большая потеря воды растением обусловлена ее большой листовой поверхностью. Одно растение кукурузы расходует за вегетационный период 200 литров воды, а сахарный тростник — в два раза больше. Растения вынуждены формировать большую листовую поверхность, чтобы получать необходимое количество СО2 для фотосинтеза, несмотря на его малое содержание (0,045 %) в атмосфере.
Например, восьмилетняя яблоня может ассимилировать за день до 50 г СО2 при интенсивном фотосинтезе. Это количество СО2 она поглощает из 300 000 литров воздуха.
Необходимость расходовать воду в больших количествах связана еще и с тем, что поглощение солнечных лучей растениями должно привести к повышению температуры, которая может вызвать коагуляцию белков. Испаряя воду, растение понижает температуру своего тела.
Для большинства растений подсыхание смертельно, поэтому расход воды должен соотноситься с приходом. Только мхи и лишайники могут выдерживать длительное отсутствие воды и переносить это время в состоянии высыхания.
Поэтому как только растения вышли из воды на сушу, начался долгий процесс по выработке приспособлений для поддержания водного баланса: во-первых, для быстрого поступления, во-вторых, для уменьшения траты наземными органами; в-третьих, для экономически выгодного транспорта по стеблю.
При анализе соотношения между поступлением и расходованием воды возможны три случая: поступление больше расходования, равно или меньше. В последнем случае возникает водный дефицит. В полдень водный дефицит может достигать 5—10 и даже 25 %. Одно из условий нормального функционирования наземных растений — поддержание условий без длительного и глубокого водного дефицита. Для этого необходима хорошо развитая корневая система, обеспечивающая поступление воды с большой скоростью.
246
Поступление воды из почвы в корень
Поступление воды из почвы — более сложный процесс, чем простое всасывание воды растением, опущенным в водный раствор корнями. В почве мы имеем ряд сил, противодействующих этому всасыванию, которые можно назвать водоудерживающими силами.
Почва — многофазная система, состоящая из четырех основных компонентов: твердых минеральных частиц, органического вещества (гумуса), почвенного раствора и почвенного воздуха.
Минеральные частицы и гумус образуют почвенную структуру, вода и воздух заполняют полости этой структуры.
Способность почвы удерживать воду зависит от ее состава и свойств. Определенное количество воды входит в состав минеральных компонентов почвы и недоступно растениям (гигроскопическая вода). Разнообразные глинистые минералы и гетерогенные гумусовые вещества, которые представляют собой коллоиды, могут также удерживать значительное количество гидратационной воды. Такая вода условно называется связанной (поеночной) и тоже труднодоступна. Вода, находящаяся в капиллярах почвы (капиллярная вода), легко всасывается корневыми волосками и поверхностными клетками сосущей зоны корня. Такая вода счи- тается свободной.
Кроме того, в почве присутствует раствор, а не чистая вода; поэтому сам раствор имеет сосущую силу. При равновесии концентраций в почве и в сосудах ксилемы в общем случае поступление воды должно остановиться.
Чтобы вычислить количество недоступной почвенной влаги, поступают следующим образом. Растения выращивают в сосуде с непроницаемыми для воды стенками (стеклянный или металли- ческий), и после того как растения хорошо разовьются, почву перестают поливать и оставляют в притемненном месте до завядания растения. Завядание указывает, что доставка воды в корни прекратилась. То количество воды, которое остается в почве к моменту завядания, и будет недоступной для растения (мертвый запас). Количество недоступной воды получило название коэффициента завядания, или влажности завядания. Следовательно, коэффициент завядания характеризует влажность, при которой устойчивое завядание только начинается.
В этот момент в почве имеется еще некоторое количество воды, хоть и малое, но в какой-то степени доступное для растения. В этих условиях даже сильно подвядшее растение оживает,
247
если его начать поливать. Полная гибель наступает только при влажности значительно ниже, чем коэффициент завядания.
При поступлении воды в сухую почву она впитывается очень быстро. Затем скорость просачивания воды в нижние горизонты становится медленной. Когда скорость нисходящего тока воды резко понижается, влажность почвы достигает уровня, который называется полевой влагоемкостью.
Под доступной для растения почвенной влагой имеется в виду количество воды, которое накапливается в почве от уровня влажности устойчивого завядания до полевой влагоемкости.
Почву можно рассматривать как резервуар, количество воды в котором то увеличивается, то уменьшается. Если после дождя почва находится в состоянии полевой влагоемкости, то ее водный потенциал близок к нулю; вода легко поступает в корни. По мере подсыхания почвы ее водный потенциал снижается.
Водный потенциал максимален в почве, несколько меньший в клетках корня и самый низкий в клетках, прилегающих к эпидермису листа (табл. 4.2).
Таблица 4.2
Величины водного потенциала в системе «почва — растение — воздух»
Компоненты системы |
Водный потенциал, бар |
|
|
Почвенный раствор |
–0,5 |
|
|
Корневой волосок |
–0,9 |
|
|
Корень (кора) |
–2,0 |
|
|
Стебель |
–5,0 |
|
|
Ëèñò |
–15,0 |
|
|
Воздух |
–1000,0 |
|
|
Первая причина возникновения градиента водного потенциала — активное поступление солей и перемещение из одной клетки в другую. Вторая — живые клетки паренхимы центрального цилиндра выделяют растворимые органические и минеральные вещества в сосуды и таким образом поддерживают градиент водного потенциала в сосудах. Поступая в сосуды ксилемы, они образуют осмотический градиент.
Химический потенциал воды, в которой эти ионы находятся, ниже потенциала воды в разбавленном солевом растворе внешней среды. Поэтому вода должна перемещаться по градиенту водного потенциала в ксилему корня.
248
Сегодня выдвинута гипотеза, подтвержденная экспериментально, о том, что в корнях есть специализированный механизм активного накачивания воды (водная помпа), работа которой не зависит от поступления ионов.
Действительно, для поступления воды в корень необходима энергия, поэтому этот процесс зависит от скорости аэробного дыхания корней — основного источника АТФ. Таким образом, одним из главных факторов, обусловливающих поступление воды, является О2. Отсюда условия, способные подавить дыхание, резко уменьшают поступление воды. В качестве примера рассмотрим следующее явление: прошел сильный дождь, в низком месте собралось много воды, а растение завядает. Почему? Избыток воды в почве вытеснил из нее воздух, поступление кислорода в корень затруднено, дыхание подавлено. Сразу же наблюдается торможение поступления воды в корневую систему. Этот пример объясняет, почему на затопленных водой почвах растения развиваются плохо и даже гибнут.
Затопление приводит к уменьшению количества О2, к увели- чению концентрации СО2 в почве, который повреждает мембраны корневых волосков; снижается поступление воды, что подтверждается торможением выделения пасоки.
Поступление воды в корень зависит и от температуры почвы. Например, на холодных болотистых почвах, несмотря на большое количество воды, растения испытывают недостаток воды, поскольку при низких температурах подавляется дыхание и нарушается снабжение корней энергией. У растений в этих условиях формируется ксероморфная структура (мелкие клетки, много устьиц и т. д.), характерная для сухих мест.
Состояние растений, при котором вода не может поступать, несмотря на ее большое количество в окружающей среде, получи- ло название физиологической засухи.
Различные поллютанты также могут влиять на поступление воды, в частности через подавление дыхания.
Необходимость поглощать большое количество воды даже в условиях ее лимита, например в условиях засухи, приводит к тому, что растение формирует огромную корневую систему. В результате корни проникают в почву на большую глубину. У пшеницы длина корней достигает 90 см, у люцерны — 120 см. Если считать, что глубина пахотного горизонта составляет 20—25 см, то большая часть корневой системы размещается ниже. Размер корневой системы характеризуется не только глубиной их проникновения в почву. Большое значение имеет общая поверхность.
249
Вода в почве двигается очень медленно: на протяжении месяца она продиффундирует не больше чем на 30 см. Перемещение кончика корня в почве опережает движение воды. Таким образом, не вода движется к корню, а корень к воде в процессе роста. Рост является первой и важнейшей особенностью корня как органа, поглощающего воду.
В засушливых условиях формируется корневая система в 3—4 раза большая, чем во влажных.
Разветвление и быстрый рост помогают корню двигаться к воде, но, с другой стороны, вода — необходимое условие роста. Как видно уже не на клеточном, а на организменном уровне, мы сталкиваемся с примером обратной связи, которая лежит в основе регуляторных процессов.
Как специализированный орган, поглощающий воду, корень владеет еще одним важным свойством — положительным гидротропизмом, при недостатке воды растущие части корней выгибаются в стороны более влажных участков почвы.
Рост корней обычно опережает рост наземных органов. Это очень важная особенность, ведь корень должен обеспечить потребности формирующегося растения в воде.
Однако клетка любого органа, которая не насыщена водой, тоже может поглощать воду, как только она будет приведена с ней в соприкосновение. Поэтому и листья, особенно подвядшие, при погружении в воду довольно энергично ее всасывают; с другой стороны, несмотря на кутикулу, вода может поступать через поверхность листа. Показано, что только сухая кутикула почти непроницаема для воды; при смачивании она набухает и делается проницаемой, поэтому смоченные дождем или росой листья могут поглощать до 25 % падающей на них воды. Это имеет практи- ческое значение при орошении растений дождеванием.
Говоря о перемещении воды по растению, выделяют транспорт в тканях одного органа, который называют ближним (радиальный), и транспорт между отдельными органами, дальний. Между ними есть существенная разница. Ближний транспорт идет по неспециализированным тканям, а для дальнего в растениях имеются специальные проводящие ткани.
Радиальный транспорт воды
Перед тем как перейти к разговору о радиальном транспорте воды, рассмотрим в общих чертах строение поперечного среза корня однодольных и двудольных растений (рис. 4.11).
250
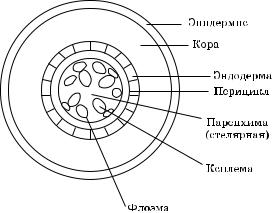
Ðèñ. 4.11. Схематическое изображение поперечного среза корня
В обоих типах растений имеется центральный цилиндр (стела), в котором находятся сосудистые пучки и паренхима (стелярная). При помощи сосудистых пучков (флоэма) поддерживается связь между разными частями растения и местами синтеза углеводов. По сосудистым пучкам (ксилема) поступает из корня в побег вода и растворенные вещества. Если перемещаться к периферии, то далее идет перицикл — однорядный слой клеток.
Затем идет эндодерма — тоже однорядный слой клеток, особенно хорошо приспособленный к тому, чтобы служить барьером, отделяющим проводящие ткани от коры. После того как растяжение клеток заканчивается, в радиальных и поперечных оболоч- ках эндодермы выявляется прослойка материала, которая дает положительную реакцию на лигнин. Это поясок Каспари. Пояски Каспари сильно ограничивают движение воды, ионов в клеточ- ных оболочках; они эффективно блокируют апопластический транспорт.
Первым барьерную функцию эндодермы выявил Д. Пристли в 1920 г.
Большую часть корней занимают клетки коры. На их долю (вместе с эпидермисом) приходится 86—90 % площади. Клетки коры вытянуты параллельно главной оси, тонкий слой цитоплазмы (1—5 мкм) окружает в них центральную вакуоль, которая занимает около 90 % объема протопласта. Вторичные изменения в
251