

- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с. Электронное оглавление
- •10. Генетические функции 19
- •11. Передача информации в клетках 46
- •12. Генетический код 79
- •13. Генетический контроль синтеза днк 115
- •14. Рекомбинация 143
- •15. Регуляция экспрессии генов у прокариот 179
- •16. Регуляция экспрессии генов у эукариот 218
- •17. Генетический анализ развития 260
- •18 .Генетика соматических клеток: картирование генома человека 302
- •19. Количественные признаки 346
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая информация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •"Врожденные ошибки метаболизма" по Гэрроду
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гипотеза "один ген - один фермент"
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Биохимические превращения при метаболизме
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гены и белки
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Структура белков
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Внутригенная комплементация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •11. Передача информации в клетках
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез днк
- •Синтез рнк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез белка
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Специализированный перенос информации Репликация рнк
- •Обратная транскрипция
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансляция днк
- •Запрещенные (неизвестные) варианты переноса информации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Колинеарность генов и полипептидов (прокариоты)
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Колинеарность генов и полипептидов (эукариоты)
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Центральная догма
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •12. Генетический код
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическое изучение кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическое подтверждение существования терминаторных кодонов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Расшифровка кода с помощью биохимических методов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Особенности построения генетического кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Терминаторные кодоны
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Организация нуклеотидной последовательности фагаφ χ74
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетические факторы, влияющие на трансляцию кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический код митохондрии
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •13. Генетический контроль синтеза днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Полимеризация днк в репликативной вилке
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ репликации днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Биохимический анализ репликации днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Инициация синтеза днк в точке начала репликации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез днк у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Точность синтеза днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Исправление ошибок репликации и репарация днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •14. Рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Общая рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Консервативный разрыв и воссоединение
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ рекомбинации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Высокая отрицательная интерференция и генная конверсия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Образование структур Холлидея у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Сайт-специфическая и незаконная рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Интеграция и эксцизия профагаλ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Подвижные генетические элементы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Общая картина метаболизма днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •15. Регуляция экспрессии генов у прокариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Участки днк, контролирующие транскрипцию
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Катаболитная репрессия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Бактериофагλ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Опероны биосинтеза аминокислот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Регуляция экспрессии генов с помощью сайт-специфической рекомбинации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •16. Регуляция экспрессии генов у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Участки днк, контролирующие транскрипцию
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Сплайсинг гяРнк-транскриптов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Транскрипция и структура хроматина
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Согласованная регуляция экспрессии генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Метилирование днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Регуляция гемоглобиновых генов в ходе развития организма
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Контроль экспрессии генов, основанный на перестройках днк
- •Типы спаривания у дрожжей
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Белки клеточной поверхности у трипаносомы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Иммуноглобулиновые гены
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •16. Регуляция экспрессии генов у эукариот 243
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Заключение
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •17. Генетический анализ развития
- •Дифференциальная экспрессия генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Клеточная детерминация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Эмбриогенез мыши
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ развития
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Мутации, затрагивающие эмбриогенез у мыши
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Мутации с материнским эффектом у дрозофилы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гомеозисные мутации уDrosophila
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Определение пола и дозовая компенсация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Определение пола у млекопитающих
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Механизмы генетической регуляции
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •18 .Генетика соматических клеток: картирование генома человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Геном человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гибридизация клеток в культуре
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Отбор клеточных гибридов с помощью методаHat
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Внутрихромосомное картирование генов с помощью хромосомных перестроек
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Микроклетки и изолированные хромосомы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Картирование генов с помощью днк-зондов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гибридизация in situ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая карта человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансфекция и наследственные болезни
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Онкогены
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансфекция генов и рак
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетика соматических клеток растений
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •19. Количественные признаки
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Пенетрантность и экспрессивность
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гены-модификаторы и гены, имеющие эпистатическое действие
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Плейотропия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Непрерывная изменчивость
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Окраска семян у пшеницы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Полигенное наследование
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая и средовая изменчивость
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Наследуемость в различных популяциях
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с. Оглавление
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
Регуляция экспрессии генов с помощью сайт-специфической рекомбинации
Рассмотренные примеры механизмов генетической регуляции основаны на использовании перечисленных в начале этой главы типов регуляторных элементов - белков-регуляторов, низкомолекулярных эффекторов и регуляторных центров. Однако следует отметить, что этим списком не исчерпываются регуляторные возможности прокариот.
Так, бактерии рода Salmonella обладают жгутиками, используемыми для передвижения. Эти жгутики содержат один основной структурный белок - флагеллин; однако бактерии обладают двумя структурными генами, которые кодируют два иммунологически различных типа флагеллинов. Два соответствующих белка обозначают HI и Н2. Структурные гены этих белков не являются тесно сцепленными. В клетках всегда экспрессируется только какой-либо один из них. Ген Н2 тесно сцеплен и экспрессируется синхронно с геном rН1, продукт которого служит репрессором гена HI. Таким образом, при экспрессии пары H2, rН1 синтезируется только флагеллин Н2, поскольку репрессор rН1 предотвращает экспрессию гена HI. Напротив, при подавлении экспрессии H2 и rН1 происходит синтез флагеллина HI. Явление альтернативной экспрессии
Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
200 Экспрессия генетического материала
|
Рис. 15.24. Доказательство наличия спонтанно инвертируемой последовательности в регуляторной области оперона Н2-rН1. А. Плазмиду, содержащую клонированную регуляторную область оперона, размножают для получения большой популяции плазмидных молекул, которые затем линеаризуют обработкой рестриктазой EcoRI (плазмида содержит единственный сайт для EcoRI). Далее проводят денатурацию ДНК и отжиг. Б. Образовавшиеся гетеродуплексные молекулы ДНК анализируют с помощью электронного микроскопа. Ообразный участок негомологии, обнаруживаемый в составе некоторых гетеродуплексных молекул ДНК, свидетельствует о спонтанной инверсии, имеющей место в ходе роста плазмидной популяции. (По Zieg J. et al., 1977. Science 196, 170.) [Copyright 1977 by the American Association for thé Advancement of Science.] |
|

Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
15. Регуляция экспрессии генов у прокариот 201
|
|
|
Рис. 15.25. Модель фазовой вариации, основанная на представлении об инверсии нуклеотидной последовательности, содержащей промоторную область оперона Н2-rН1. (По Zieg J. et al, 1977. Science, 196, 170.) |
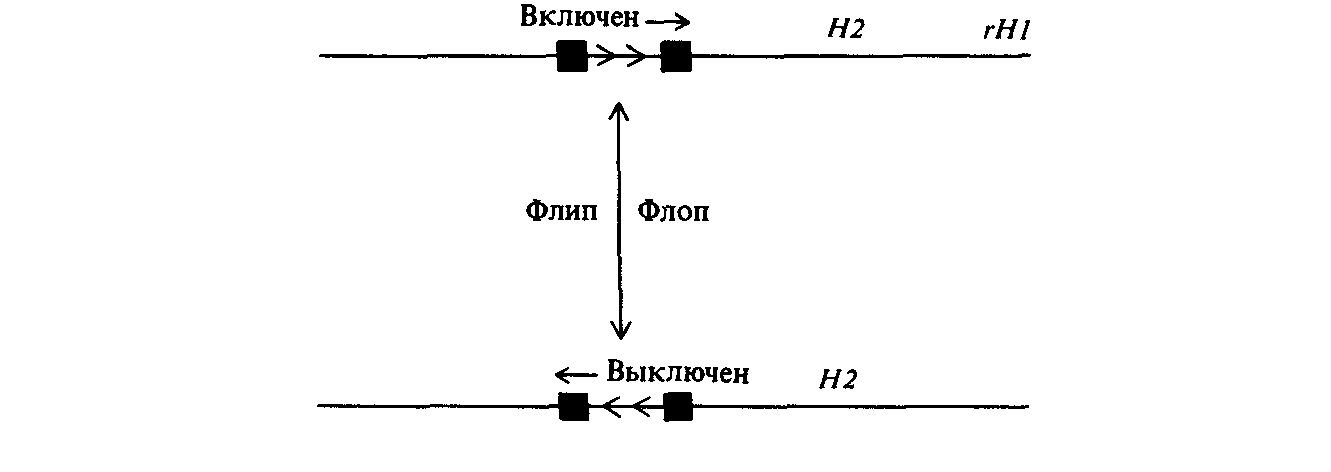
генов H1 и Н2 получило название вариации фаз. В культуре, выращенной из клетки, находившейся в какой-либо одной из двух фаз, всегда обнаруживаются клетки в обеих фазах. Частота перехода клеток из одной фазы в другую для различных штаммов Salmonella варьирует в пределах от 1 на 105 до 1 на 103 клеток за генерацию. Вариация фаз для бактерий Salmonella, вероятно, является одним из способов избежать иммунологического отторжения в ходе инфекции.
Контроль вариации фаз зависит от состояния промотора, направляющего транскрипцию генов H2 и rН1. Это было показано с помощью клонирования соответствующей регуляторной области на плазмидном векторе с использованием методов, описанных в гл. 9. При выращивании клеток E.coli, содержащих плазмиду со встроенной регуляторной областью, можно получить огромную популяцию идентичных молекул. Плазмидную ДНК очищают и линеаризуют с помощью рестриктазы Eco RI (рис. 15.24). Денатурация и отжиг цепей ДНК приводит к образованию гетеродуплексов, содержащих цепи из различных исходных плазмидных молекул. Некоторые из этих гетеродуплексных молекул содержат негибридизующийся участок протяженностью около 800 п. н. (рис. 15.24). Появление этого участка, как оказалось, вызвано тем, что в некоторых плазмидах произошла переориентация участка протяженностью 800 п. н. Это наблюдение позволило предложить модель механизма вариации фаз, согласно которой промотор генов Н2 и rН1 может находиться в одной из двух ориентации («флип» или «флоп»). В одной из них транскрипция с этого промотора приводит к образованию белков Н2 и rН1. При другой ориентации транскрипции этих генов не происходит (рис. 15.25).
Было показано, что инверсия промоторной области у Salmonella направляется системой сайт-специфической рекомбинации. По обе стороны от инвертируемого участка находятся идентичные противоположно ориентированные сегменты последовательности ДНК по 14 п. н. Наряду с промотором генов Н2 и rН1 этот участок содержит также ген (hin), продукт которого направляет сайт-специфическую рекомбинацию между 14-членными повторами. В результате рекомбинации происходит инверсия всего этого участка ДНК. Фаги Р1 и Ми также кодируют белки сайт-специфической рекомбинации, которые могут направлять инверсию сегментов ДНК в их собственных геномах. Оказалось, что эти белки могут заменить дефектный белок Hin в мутантах Salmonella hin -.