

Дәріс № 4-5. Инженериялық генетика. Генетикалық инженерия әдістерімен алынған, микроағзалардың өнеркәсіптік штамдары.
1. Қажетті генді алу
2. Оны, репликацияға қабілетті, генетикалық элементке (векторға) құру.
3. Реципиент – ағзаға, вектор құрамына кіретін, генді енгізу.
4. Қажетті ген ие болған, жасушаларды сәйкестендіру (скрининг және селекция)
Қазіргі биотехнологияны көбінесе генетикалық инженерия негізіндегі биотехнология ретінде сипаттайды. Шындығында, бұл жасанды жасалған генетикалық бағдарламаларды жүргізудің нәтижесінде, бионысандардың бағытталған түрөзгергіштігіне қолданылатын негізгі жол. Кейде генетикалық инженерияның үш деңгейін ажыратады: 1) гендік – жеке гендерден тұратын, рекомбинантты ДНҚ – ымен тікелей манипуляциялау; 2) хромосомдық – гендердіңүлкен топтарымен немесе тұтас хромосомаларымен манипуляциялар; 3) геномдық - генетикалық материалдың барлығын немесе үлкен бөлігін біржасушадан басқасына тасымалдау. Қазіргі түсінікте генетикалық инженерияға рекомбинантты ДНҚ технологиясы кіреді. Генетикалық инженерия облысындағы жұмыстар төрт негізгі кезеңнен тұрады: 1) қажет генді алу; 2) оны репликацияға қабілетті, генетикалық элементке (векторға) құру; 3) реципиент – ағзаға, вектордың құрамына кіретін, генді енгізу; 4) қажетті генге ие болған, жасушаларды сәйкестендіру (скрининг және селекция). Әрбіркезеңді жеке – жеке қарастырайық. Гендерді алу. Қажетті генді келесі түрмен алуға болады: а) оны ДНҚ – дан бөлумен; б) химиялық - ферментатитвті синтез жолымен; в) РНҚ тәуеді ДНҚ полимеразаның (ревертозаның) көмегімен, оқшауланған матрицалық РНҚ – ның негізінде жаңғыртумен. Гендерді ДНҚ – дан бөлу. Оқшауланған ДНҚ бөлшектеуге ұшыратады. Ол үшін, нуклеотидтердің белгілі тізбектеріне ие болатын, үлескілерде ДНҚ ыдырауын өршітетін, рестрипциялық эндонуклеозаларды пайдаланады. Қазіргі уақытта, 85 әртүрлі нуклеотидті тізбектерді танитын, 400 – ден астам рестриктазалар белгілі. Ыдырау нуклеотидті жұптардың танылатын үлескісінің ортасында жүруі мүмкін, сонда ДНҚ – дың екі тармақшасы да бір деңгейде «қиылады». Түзілген бөлшектер екітармақты (тұйық) ұштарға ие болады. Басқа рестриктазалар ДНҚ – ның тармақшаларын ығыстырумен ыдыратады, сондықтан баспалдақ түзіледі. ДНҚ жіпшелерінің бірібірнешенуклеотидтерге шығыңқы болады. Біржіпшелі (жабысқақ) ұштары түзіледі. Бір рестриктазаның әрекетімен алынған, ДНҚ – дың екі жабысқақ бөлшегі кездессе, онда ұштық тізбектерінің әсерінен өзара әрекетте оңай түседі. Қажет болғанда тұйық ұштары жабысқақтарға айналуы мүмкін. Ол үшін, тұйық ұштарға, жабысқақ ұштар беретін, рестриктазаның тану үлескілерімен екітізбекті тізбектілерді қосады. Жабысқақ ұштарымен нуклеотидті тізбектілік болуы мүмкін: а) алдын – ала сол рестриктазамен өңделген, векторға қосылуы; б) өзара комплементар ұштарды тігу жолымен сызықтық молекуладан сақиналыққа түрленуі мүмкін. Рестриктазалардың көмегімен ДНҚ – дан гендерді әдісі мәнді кемшіліктерге ие болған. Қажет генге сәйкес келетін, сол бірүлескіні ДНҚ – дан кесіп алуға мүмкіндік беретін, рестриктазаларды таңдап алу қиын.Қызықты генмен қатар, ДНҚ бөлшектреі, генді қолдануға кедергілер келтіретін, артық нуклеотидті тізбектіліктерге де ие болады. Рестриктаза нуклеотидті ген тізбектілігінің бөлігін ыдырата алады, нәтижесінде ген функционалдық толық құндылығын жоғалтады.
Эупариотты ағзалардың гендері күрделі құрылысқа ие болған: кодтайтын ақуыздан, мәнді (экзондар) және аралық, мәнсіз үлескілерден (интрондар) тұрады. Осындай ДНҚ – матрицада синтезделген, алғашқы РНҚ түрөзгеріске ұшырайды. Нәтижесінде интрондарға сәйкес келетін, үлескілер жойылады, ал, экзондарға сәйкес келетін үлескілер, қосыла отырып, жетілген матрицалық РНҚ түзеді. Интрондардың болуы трансплантацияланған гендердің қалыпты қызметіне кедергі болып табылады. Рестриктазалармен ДНҚ – ын өңдеу барысында бөлшектердің қоспасы түзіледі. Бұлқоспадан, қажетті генге ие болатын, бөлшектерді бөліп алу – оңай мәселе емес. Бактериялық жасуша шамамен бес мың гендерден, ал эукариоттық жасуша 10 – 20 мың гендерден тұрады. Гендердің химиялық – ферментативті синтезі. Бұләдіс – нативті ДНҚ – дан рестриктазалардың көмегімен гендерді «қиюға» маңызды альтернатива. Әдіс нуклеотидтердің арасында эфир байланыстарының кезеңдік түзілуі есебінен, қысқа (8 – 16 буынды) біртізбекті ДНҚ бөлшектерінің химиялық синтезінен және екі тізбекті поленуклеотидтердің түзілуінен, ДНҚ лигаза арқылы өзара олегонуклеотидтердің тігісінен тұрады.
Химиялық – ферментативтік синтез нуклоетидтердің минимальді қажетті тізбектілігін дәл жаңғыртуға және ДНҚ бөлшектерінде артық нуклеотидті тізбектіліктердің, сонымен қатар, интрондардың элиминирленуімен байланысқан, мәселелерді болдырмауға мүмкіндік береді. Сонымен қатар,әртүрлі рестриктазалардың реттегішіне тізбектіліктердің т.б. тану үлескілерін гендерге енгізу мүмкіндігі бар.
Гендердің химиялық – ферментативті синтезіне оның нуклеотидті тізбектілігі туралы толық ақпарат қажет, сондықтан әдістің қолданысы осындай ақпаратты алу мүмкіндіктерімен шектелген. Генде нуклеотидтердің тізбектілігі сәйкесті ақуыздың алғашқы құрылымы негізінде жаңғыруы мүмкін. Ген құрылымын талдаудағы триумф – кодталатын ақуызда аминқышқылды қалдықтар тізбегінің және ДНҚ нуклеотидті тізбектілігінің параллель жаңғыруы. Химиялық – ферментативтік синтез әдісімен ас – оператор Е coli, проинсулиннің, инсулиннің А және В тізбектеріндегі, соматостатиндегі гендер алынған.
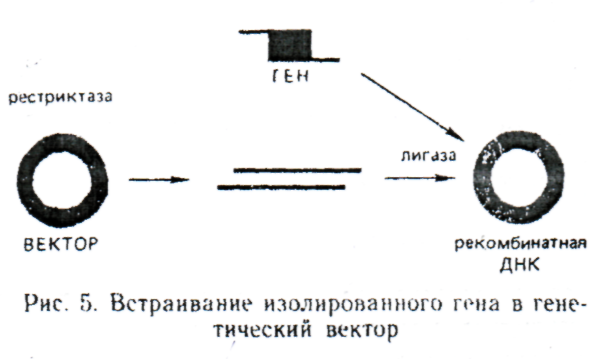
Жасушадан бөлінген матрицалық РНҚ негізінде гендердің ферментативті синтезі. Бұл гендер синтезінің ең танымал әдісі. Кері транскриптаза (ревертаза), мРНҚ – на комплементарлы, ДНҚ жіпшелерінің синтездерін өршітеді. Комплементарлық ДНҚ және кДНҚ деп аталатын, алынған біртізбекті ДНҚ – лып, ДНҚ – полимеразаны немесе ревертазаны қолданумен, ДНҚ – ның екінші жіпшесін синтездлеуге арналған матрица ретінде пайдаланады. Қарастырылатын әдістің артықшылығы, ген интрондарсыз және басқа транскрибцияланбайтын тізбектілерсіз алынады. Сонымен қатар, генді ДНҚ бөлшектерінің қоспасынан іріктегеннен гөрі, мРНҚ – ның қажет түрін жасуша аккумуляциялайтын, жағдайларды жасау оңайырақ. РНҚ тәуелді ДНҚ синтезінде негізделген, әдісті қолдануда 1979ж.адамның өсу гормонының генін алу үлкен жетістік болып табылады. Генді векторға енгізу. Сол немесе өзге тәсілмен алынған, ген ақуыз құрылымы туралы ақпаратты жүзеге асыра алмайды. Геннің әрекетін басқаратын қосымша механизмдер қажет, сондықтан генетикалық ақпаратты жасушаға тасымалдау векторлар құрамында жүзеге асырылады. Векторлар – өз бетінше реплткацияға қабілетті сақиналы молекулалар. Ген вектормен бірге рекомбинантты ДНҚ – ын түзеді. Рекомбинантты ДНҚ – ды құрастыру in Vitro жүзеге асырылады. Вектордың сақиналы молекуласы рестриктазамен айырылады. Алынған сызықты ДНҚ молекуласы, ДНҚ – ны енгізетін ұштарға комплементарлы, жабысқақ ұштарға ие болуы қажет. Енгізілетін геннің және вектордың комплементарлы жабысқақ ұштарын ДНҚ лигазаның көмегімен, біріңғай сақиналы молекула түзумен, қайта тұйықтайды. Векторлардың екі негізгі класын ажыратады: вирустар және плазмидтер. Генетикалық векторлар ретінде вирустарды пайдалану барысында туындайтын, маңызды мәселе – аттеньюация болып табылады. Аттеньюация – вируспен зақымдалған жасушалар аман қалуы және ұрпаққа өзгерген генетикалық бағдарламаны беруі үшін патогенділіктің әлсіреуі. Бүкілағза бойымен басты инфекция қысқа мерзімде дамитындай, жануар немесе өсімдік ұлпасында тарай отырып, жасушадан жасушаға тез тасымалданатын вирустардың қабілеті биотехнология үшін үлкен мәнге ие болған. Вирустардың осындай қасиеті ересек ағзадағы соматикалық жасушалардың генетикалық түрөзгеру мүмкіндігін ашады. Бұл қатынаста барлық он адам ағзасының жасышаларымен жетіспейтін гендерді таситын, вирустарды енгізу жолымен, адамның тұқым қуалайтын ауруларын емдеу жолдары ашылады.
Плазмидтер – бактериялардан, саңырауқұлақтардан, өсімдіктерден және жануарлардан табыған, автономды өздігінен репликацияланатын генетикалық бірліктер. Генетикалық инженерияда бактериялық плазмидтер, әсіресе Е coli плазмидтер кең қолданыс тапқан.
Генетикалық векторға оқшауланған генді құру. Бактериалдық плазмидтер, бактериялардың коньюгациясы жолымен жасушадан жасушаға генетикалық ақпаратты тасуға қабілетті коньюгативті және бактериалық трансформация механизмі арқылы бір жасушадан басқасына тасымалданатын коньюгативті емес деп бөлінеді. Дербес тасымалдауға қабілетті плазмида – көмекші болған жағдайда ғана, коньюгация жолымен коньюгативті емес плазмидтердің тасымалы мүмкін болады. Кейбір плазмидтер амплификацияға қабілетті, яғни жасушада көшірмелердің үлкен санын түзеді, бұл гендерді фенотипті өрнектеу дәрежесін күрт жоғарылатады.
Векторларды құру барысында зерттеуші оған рестриктозаларды тану үлескілерін, сондай – ақ оңай танылатын белгілерді кодтайтын гендерді - маркерлерді енгізеді. Осы белгілер бойынша, вектор тасымалдаушылар болып табылатын, жасышаларды іріктеуге болады.
Космидтер – плазмидтер үлкен қызығушылық оятуда - олардың құрамына, ДНҚ – ның қапталуына жауап беретін, IEcoli фагтың ДНҚ cos – үлескісі енгізілген. Осындай плазмидалар генетикалық ақпараттың өте үлкен көлемін тасымалдауға қабілетті, рекомбинантты ДНҚ фагті бөлшектерге қапталуы мүмкін.
Өсімдіктердің генетикалық инженериясына қатысты Rhizobium және Agrobacterium туыстар бактерияларының плазмидтері тиімді. Agrobacteriumtumifaciens – те Ті – плазмидтер болады, ДНҚ плазмидтердің Т – үлескісі кейбір түрлердің өсімдіктерінде геномға құрылуы мүмкін. Ті – плазмидтерде үш ген (онкоген) болады, оның екеуі ауксин синтезінің кезеңдерін кодтайды, ал үшіншісі цитопининнің синтезіне жауап береді. Қалыпты өсімдік жасушаларының өсуі бұл гормондардың тыстан түсуімен реттеледі. Интеграцияланған күйдегі Т – 1 плазмидтер – Т – үлескілерден тұратын жасушалар, қатерлі ісіктерді – өсінділерді түзе отырып, бақылаусыз көбейеді. Плазмидтер оларда онкогендерді қиюмен «қарусыздандыруы» мүмкін. Қажетті өнімді кодтайтын, генді Т – үлеске қою, генетикалық нженерия үшін пайдалы векторға плазмидтердің түрленуіне алып келеді. Бірге опиндер деп аталатын октопиннің және нополиннің – аргинин туындыларының - аномальді аминқышқылдарының синтезін зақымдалған жасушаларда Т – 1 плазмидтер индуцирлейді. Опиндердің синтезге қабілеті пайдалы генетикалық маркер болып табылады. Реципиент ағза жасушаларына гендерді тасымалдау. Плазмидаға құрылған гендерді беру коньюгация немесе тасымалдау жолымен жүзеге асырылады. Егер гендер вирустың геномына құрылса, онда ақпаратты тасудың ең тараған тәсілі тасымалдау болып табылады.
Тасымалдау – жасуша белгілерінің өзгерісін тудыратын реципиент жасушаға плазмидті және бос ДНҚ – ның тасымалдануы. Бұл кезде реципиент хромосомасына немесе қандай да бір хромосомадан тыс генетикалық бірлікке біржіпшелі ДНҚ бөлшегінің рекомбинациясы мен интеграциясы жүреді. Тасымалдануды бактериялар ДНҚ тудыруы мүмкін. Мұны алғаш рет Гриффит пневмококктардан байқаған. Бактерия жасушасына ДНҚ – ның енуі оның өкілетті, яғни сезімтал күйін қажет етеді. Streptococcus және Pneumococcus өкілдерінде, 5 – 10 мД молекулалық салмақпен ақуыздар – өкілеттік факторлары ерекшеленген және тазартылған. Жасушаның өкілеттілігі сондай – ақ сыртқы ортаның шарттарымен де анықталады. Ecoli және В subtillis- те СаСl2 – мен полиэтиленгликольмен ( ПЭГ) жасышаларды өңдеумен тиімді тасымалдауға қол жеткізіледі.
Жасушаға енетін генетикалық материал жасуша ішілік нуклеозалармен шабуылдануы мүмкін. Бұл жағдайда келесілер табысты тасымалдануға ықпал етеді: 1) нуклеозалардың белсенділігін немесе синтезін бәсеңдету және 2) липосомаларға тасымалдайтын ДНҚ қосу – жасанды жарғақшалы лепидті везикулалар.
Өсімдіктердің және саңырауқұлақтардың, негізінен ашытқылардың тасымалдануын жүргізу үшін, интактілік протопласттарды алу қажет. Тетрциклин антибиотигіне төзімді, ашытқылар осындай тәсілмен алынған. Agrobacterium tumifaciens және өсімдіктер (петуния) протопласттарын бірлесіп өсіру өсімдік жасушаларының ерекше жоғары тиімділікті тасымалдануына алып келеді. СаСl2 және ПЭГ қатысуында фосфатидилсеринхолестеринділипосомаларға қосқанда, өсімдіктердің жасушалары генетикалықвирус материалымен тасымалдануы мүмкін. Тасымалдау генетикалық ақпартты берудің ең әмбебап жолы болып табылады, ол генетикалық инженерия үшін ең үлкен мәнге ие болған. Коньюгация мен трансфекцияны, гендерді тиімді тасуға арналаған арнайы айлабұйымдармен күрделенген тасымалдаудың нұсқалары ретінде қарастыруға болады. Коньюгация жолымен кейбір плазмидалардың тасымалы ғана жүреді. Бұл жағдайда, ақуызды түтікшелер болып табылатын жыныс бүртікшелерімен бактерияларының бір (еркек донорлық) жасушасынан басқа (әйел, реципиенттік) жасушасына ақпарат көшеді.Конъюгативті тасымалдауды дербес жүзеге асыратын, плазмидтер шеңбері шектелгенімен,конъюгативтік емес плазмидтер, көмекші-плазмидтердің қатысуында, конъюгация жолымен берілуі мүмкін.
Трансфекция ретінде, жасушада вирус бөлшектерінің дамуына алып келетін, фаг немесе вирус гендерінің бүкіл жиынтығының берілісін айтады. Генетикалық инженерияда трансфекцияны жүргізу әдістемесіне бактерияларға қосымша сферопласттардың алынуын, жасушадан тыс нуклеазолардан инкубация ортасын тазалау және трансфекция тиімділігін мәнді жоғарлататын, протаминсульфатпен сәйкестікте сол немесе басқа фагтың тазартылған ДНК-ны қосу жатады. Вирустық табиғаттың сәйкес векторларымен өсімдіктер және жануарлар жасушаларының трансфекциясы протопласттар мен тазартылған ДНК-ды пайдалануда да, сондай-ақ бүтін көпжасушалы ағзалардың (бұл жағдайда көбінесе трансфекция туралы емес, инфекция туралы айтады) немесе ДНК-ның вирус бөлшектерімен зақымдануында да жүргізілуі мүмкін.
Қажетті генге (гендерге) ие болған, реципиент - жасушаларды сәйкестендіру. Тасымалдау, конъюгация немесе трансфекция жүргілілген соң, ген-насанаға ие болатын, жасушаларды сәйкестендіру қажет. Генно-инженериялық жобаның табыстылығы көбінесе пайданылған іріктеу әдісінің тиімділігіне тәуелді болады. Гендердің трансплантациясынан кейін, жасушалардың аздаған бөлігі ғана қажетті генге ие болатынын ескерсек, геннноинженериялық өңдеудегі бұл кезеңнің мәні айқындала түседі. Жасушаларды іріктеу екі кезеңде жүреді.
Бірінші кезең – сәйкесті векторға (генді транспланттауға қызмет ететін) ие болатын, жасушаларды іріктеу. Көбінесе бұл іріктеу, вектор белгіленген, гентикалық маркармен жүргізіледі. Антибиотиктерге орнықтылықтың детерминаттары векторда, антибиотикпен ортаға егу барысында, бұл векторға ие болатын, жасушалармен бактериялық популяцияны байытуғв мүмкіндік береді.
Екінші кезең - вектроға ғана емес, нысана - генге де ие болатын, жасушаларды іздеу. Ол үшін әдістердің екі тобын қолданады:
1) Реципиент-жасушалардың ДНК-ын тікелей талдауға негізделген әдістер: а) ДНК-ың нуклеотидті тізбектілігін анықтау; анықталатын генге болжамды ие болатын, жасушалардан, бұл генге ие болатын,үлескілерді іздеу жүргізілетін, вектор ДНК-ын ерекшеленеді; содан кейін нуклеодидті ген тізбектілігінің секвенделуін жүргізеді; б) жасушалардан бөлінген, қажетті ген немесе оған сәйкесті РНК-ы болуы мүмкін болатын шатырмен ДНК-ның гибридтелуі. Алдын-ала оқшауланған ДНК-ын біртізбекті күйге ауыстырады және біртізбекті ДНК - (немесе РНК) шатырымен өзара әрекетке түсіреді. Одан кейін екітізбекті гибридті ДНК молекулаларының болуын анықтайды.
2) Генмен кодталатын, белгіні сәйкестендіруге негізделген әдістер; а) генмен кодталатын, ферменттер синтезінде болатын, қосылысты түзететін жасушалардың немесе нысана – ген трансляциясының және транспкрипциясының өнімін – ақуызды синтездейтін, жасушаларды тікелей іріктеу-осылайша химерлік плазмидалар қоспасымен тасымалданған, жасушалар популяцияларынан гистидин синтездейтін ашытқылырды осылайша іріктеген; б) нысана - генге ие болған жасушалардың ғана өсуін қолдайтын, селективті орталарды пайдалану; мысалы, р-галактозидаза геніне (лактозаны жоюға қажетті ферментке) ие болатын, жасуша-рецепенттер, көміртектің жалғыз көзі ретінде лактозамен ортада бактерия жасушаларын өсіру жолымен, іріктелуі мүмкін; в)иммуннологиялық детекция: егер рекомбинантты ДНК құрамында анықталатын ген транкрибцияланса және тасымалданса, бірақ ағза фенотипіне ешбір әсер көрсетпесе ғана қолданылады; мысалы, егер ген адамның а-интерферонын коттаса, бактерия жасушалары ризаланып, артынан анти денелермен а-интерферонға анти геннін байланысу реакйиясын жүргізеді.
Генетикалык инженерия және процудент - жана ағзаларды құрастыру. Генетикалық инженерия әдістерінің көмегімен, өте әр түрлі өнімдерді синдездеуге қабілетті, микро ағзалардың жана формалардың, сонымен қатар, өсімдіктен және жануардан алынатын өнімдерді белгіленген жоспар бойынша құрастыруға болады (Н.С.Егоров, В.Д.Самуилов.1985). Бұл кезде микроағзалардың өнімділігін және жоғары өсу жылдамдығын, әртүрлі шикізат түрлерін жоюға қабілеттілігін ескеру қажет биотехнологияның алдындағы кең келешек адам ақуыздарының микробиологиялық смнтез мумкіндігіне жол ашады: осындай тәсілмен соматостатин, интерферондар, инсулин, өсу гармоны алынған.
Жаңа процудент - микроағзаларды құрастыру жолындағы негізгі мәселелер келесілерге негізделген:
1. Өсімдіктен, жануардан және адамзаттан алынған гендер өнімділері оларға бөтен жасуша ішілік ортаға түседі, мұнда микробты протеазалармен ыдырауға ұшырайды. Ерекше тез бірнеше минутта соматостатин секілді қысқа пептидттер гидролизденеді. Микробтың жасушада генноинженерлік ақуыздарды қорғау старатегиясы келесілерге негізделген: а) протеазалар ингибаторларын қолдануға; протеаза ингибиторның синтезіне жауапты, РТ генмен Т4 фагтың ДНК-бөлшегін,негізгі интерферон генін плазмидалар енгізу барысында адамзат интерферонының шығымы 4 есеге өсті (В.И.Таниянин, 1985) ; б) гибридті ақуыз молекуласының құрамында керекті пептидті алуға; в) гендердің амплификациясына (көмірлер санының ұлғаюына); плазмида құрамында адамзат проинсулиннің генін көпреттік қайталау Е.coli жасушасында бұл ақуыз мультимөлшерінің синтезіне алып келеді,ол мономерлік проинсулинге қарағанда,жасуша ішілік протеазалардың әсеріне мәнді түрде орнықты болып шықты.
2. Көп жағдайда трансплантацияланған геннің өнімі егу ортасына босатылмайды және жасуша ішінде жинақталады, бұл оның бөлінуін мәнді қиындатады. Мысалы, Е.coli көмегімен инсулинді алудың қабылданған әдісі жасушалардың ыдырауын және инсулинннің тізбекті тазалануын болжайды. Генетикалық инженерияның сүйікті нысаны Е.coli-ден басқа блонысандарға биотехнологиялардың қайта бағдарлануы да ақталған.Сонымен қатар, Е.coli салыстырмалы аз ағуыздарды эпскреттейді. Сондай-ақ, бұл бактерияның жасуша қабырғасы тосинді зат эндокотиннен тұрады, оны фарманалогиялық мақсаттарда қолданылатын өнімдерден мұқият бөлу қажет. Генетикалық инженерияның нысандары ретінде грамоң бактериялары (Bacillus, staphulococcus, streptomuces, туысының өкілдері) тиімді. Негізінен Bacillus,subtillis дақылды егу ортасына 50-ден астам әр түрлі ақуыздарды бөледі. Оларды қатарына ферменттер,инцептициттер, антибиотиктер кіреді. Сондай-ақ эукароиттық ағзаларда тиімді. Олар артықшылықтар қатарын иеленген, негізінен, ашытқы интерфероны, адамзат ақуызы сияқты, гликолизделген формада синтезделеді (Е.coli жасушаларында синтезделетін интерфероннан ерекшелігі.
3. Көптеген тұқым қуалайтын белгілер бірнеше гендермен котталады, және генноинженерлік өңдеу әрбір гендердің тізбекті трансплантация кезеңінде ие болу керек. Жүзеге асырылған көпгенді жобаның мысалы, шикі мұнайды жоюға қабілеті, Pseudomonas штамын жасау болып табылады. Плазмидалардың көмегімен, октанды, комфораны, ксилолды, нафталинды ыдырататын, ферменттердің гендерімен тізбекті байытылды. Кейбір жағдайларда, бір плазмиданың көмегімен тұтас гендер болктарынын бір уқытта транплантациясы мүмкін болады. Нитрогеназды кешеннің құрылымдық құрамдарына да, сондай-ақ синтездерінің реттелуіне жауап беретін, кем дегенде 17 әр түрлі болуымен ағзаның азотты бекітуге қабілеттілігі анықталады. Қазігі уақытта генетикалық инженерия барлық жандылардың патшалығын игерген. "Бөтен" гендердің фенотиптік өрнектелуі бактерияларда ғана емес, сонымен қатар ашытқыларда, саңырауқұлақтарда, өсімдіктерде, жануарларда да алынған. Гентикалық инженерияның ыңғайлы, жақсы зерттелген және өндірістік құнды нысандары Saccharomyces (шарап, наубайханалық, сыра ашытқылары), Zymomanas (этанол алуға арналған) туыстарының өкілдері, ашытқылар болып табылады. Табысты генноинженерлік өңдеулердің қатарына гепатит вирусының беткі антигенін, а-интерферонды кодтайтын, гендерді ашытқыға енгізу жатады. Антибиотиктердің продуценттері ретінде саңырауқұлақтармен генноинженерлік жұмыстар жүргізіледі. Генетикалық өсімдіктер инженриясының дамытудың магистролді жолдарына келесілер кіреді:
1) басқа өсімдіктерден алынған, гендердің көмегімен қосымша заттармен (зеин, секалин, глутенин, легумин, глиаин, альбулин) өсімдіктерді байыту;
2) ақуызмен байланыстыратын (6-хлорофилл, бисфосфаткарбоксилаза, рибулозо-1,5 гендер негізінде өсімдіктер фотосинтезінің тиімділігін жоғарылату;
3) мысалы, азотты тасымалдауда және қорға жинауда қатысатын, глутамин-синтазаны кодтайтын, гендерді қолданумен, азотты метаболизмді өзгерту;
4) Гербицидтерге, топырақтардың тұздануына, жоғары және төмен температураларға, сыртқы ортаның басқа жайсыз факторларына төзімділікті беру.
Сонымен қатар, өсімдіктер адамының ақуыздарын-инсулинді, интерферонды, өсу гармонын алу үшін қолданыла алады.
Генетикалық өсімдіктер инженериясы жасушалардың ядролық генонымен ғана емес, сонымен қатар, митохондриялардың және хлоропласттардың генонымен де манипуляцияларға ие болады. Өсімдіктің азотты тыңайтқыштарға мұқтаждығын жою үшін, хлоропластты геномға азотфикация генін енгізген өте тиімді. Біздің уақытта гентикалық векторлардың және бұл векторлардан тұратын, өсімдіктердің биотехнологтардың қадағалауынан шығып кету мүмкіндігі қауіп төндіреді. Біріншіден, генноинженерлік дәнді дақыл өсімдіктерінің арам шөптерге түрлену қаупі айтылады. Басқа өсімдіктерге залал келтіре отырып,табиғатта тез таралуға қажетті белгілердің, "арам шөптіліктің" кешені - тұқымдарды себудің тиімді механизмдері, сыртқы ортаның жайсыз факторларына бейімделу, т.б. – бір немесе бірнеше гендерді трансплантациясына байланысқан гербинидтерге төзімділік егу айналымдарында мәселелер тудыруы мүмкін: белгіленген егу алаңында егілген гербицидтерге төзімді өсімдік, оны алмастырған ауылшаруашылық дақылына қатысты, келесі жылда арамшөп болып табылады, оған гербицидтердің күші жетпейді.
Екінші қауіп-гентикалық түр өзгерістен туындаған, биохимиялық өзгерістер өсімдікте тақамдық немесе азықтық құндылықтың жойылуына және тіпті улы болуына алып келуі мүмкін.
№ 6 Дәріс. Жасушалық инженерия.
1. Гибридті жасушаларды алу кезеңдері.
2. Жасушалардың қосылу әдістерінің мүмкіншіліктері.
3. Гибридті технология.
4. Өсімдік сұрыптары мен микроорганизмдер штамдарының жаңа түрлерін шығару және бар түрлерін жақсарту.
Жасушалық инженерияның негізі - соматикалық жасушаларды гибридтеу болып табылады, яғни жыныссыз жасушалардың бірігіп біреуге айналуы.
Гибридті жасушаларды алу кезеңдері. Жасушалардың қосылуына плазмалық мембраналардың жақындасуы әсер етеді. Оған табиғи мемранадағы зарядтың болуы бөгет жасайды, яғни теріс зарядталған ақуыздар мен майлар тобы.
Мембрананың ауыспалы электрлі немесе магнитті өріспен деполяризациялануы, жасушалардың қосылуына - катиондардың көмегімен мембрананың теріс зарядының бейтараптандырылуы әсер етеді.
Қосылып кетудің алдында өсімдік, саңырауқұлақ және бактериялық жасушалар жасуша қабырғасынан босатылады, соның салдарынан протопласттар пайда болады.
Лизоцим (бактериялық жасушаларға арналған), ұлудың зимолиазасын (саңырауқұлақ жасушаларына), саңырауқұлақтардан шығарылатын целлюлазалар кешенін, гемицеллюлаза және пектиназаны (өсімдік жасшаларына арналған) қолдана отырып жасуша қабырғасын ферментативті гидролизге ұшыратады.
Протоплатардың ісіну және бұзылуын ортаның осмостығын ұлғайту арқылы алдын алады.
Алынған гибридті жасушалардың скринингі үшін әртүрлі әдістер қолданылады: 1) фенотиптік белгілерді есепке алу; 2) тек ата-аналарының жасушаларының геномы қосылған гибридтер ғана тіршілігін сақтайтын таңдаулы шарттар жасау.
Жасушалардың қосылу әдісінің мүмкіндіктері.
1. Жекеленген пішін түрлерін филогенитикалық шағылыстыру мүмкіндігі.
2. Бір ата-анадан толық гендер жиынын, ал екінші ата-анадан дербес жиынды тасушы ассиметриялық гибридтерді алу. Мұндай гендер организмдер жасушалары қосылған кезде жиі кездеседі.Ассиметриялық гибридтер гендерді толық тасушы симметриялық гибридтерге қарағанда орнықтырақ, жемістірек және өмірсүргіш келеді.
3. Үш немесе одан да көп ата-аналық жасушаларды қосу арқылы гибридтерді алу. Осындай гибридті жасушалардан регенерантты өсімдіктерді (саңырауқұлақтар) өсіру мүмкіндігі бар.
4. Әртүрлі даму программасын тасушы жасушаларды гиридизациялау. Яғни - әртүрлі ұлпалар немесе органдар жасушаларының қосылуы, қалыпты жасушалардың жасушалармен қосылуы - яғни қатерлі өзгеру нәтижесінде даму программасы өзгергентүрлер.Осыдан гибридомды жасушалар немесе гибридомалар пайда болады,олар қалыпты ата-ана жасушасынан әртүрлі пайдалы қосылысты синтезге кабілетін алады, ал қатерліден - тез әрі тоқтамсыз өсуді алады.
Гибридомді технология. Қазіргі уақытта клеткалы инженерияның негізгі бағыты - гибридомды алу болып табылады. Негізгі мақсат - қатерлі ісік жасушасы мен алынған гибридомды жасушы сызығының қосылып бағалы заттар шығаратын жасушаның тіршілігін мәңгілік ету. Гибридомді технология көп жағдайда жануар жасушасына қолданыдады, оның көмегімен гормондар және қанның ақуызды факторларының шектеусіз көбеетін продуценттерін алуды жоспарлайды. Иммундық жүйенің қатерлі ісік клеткалары (миолема) мен сол жүйенің қаліпті жасушалары - лимфоциттер қосылуынан пайда болған өнім - өте үлкен практикалық мағынасы бар гибридомалар болып табылады.
Адам немесе жануар организміне бөгде зат - бактерия, вирус, «бөгде» жасуша немесе күрделі органикалық қосылыс түскенде - лимфоциттер түскен бөгде затты залалсыздандыру үшін күш жұмылдырады. Атқаратын қызметтеріне қарай лимфоциттердің бірнеше популяциясын ажыратады.
Т-лимфоциттер, олардың ішінде ағзаға енген бөгде затқа шабуылдап белсенділігін төмендететін Т-киллерлер ерекшеленді, және В-лимфоциттер, басты мақсаты иммунды ақуыз (иммуноглобулин) өндіру, олар бөгде затты беткі қабатымен байланыстыру арқылы залалсыздандырады, басқаша айтқанда В-лимфоциттер бөгде затқа антидене болып табылатын иммундық ақуыз бөліп шығарады.
Т-лимфоцит-киллер мен ісік жасушасы қосылып тоқтамсыз көбейетін жасушалар клонын береді. Осындай Т-киллер гибридом клондарын науқас оганизміндегі қатерлі ісік жасушаларымен күресу үшін қолдану жоспарланып отыр.
В-лимфоцит пен миолем жасушасы қосылуының нәтижесінде В-гибридомді клондар пайда болады. Олар антидене продуценті ретінде кең қолданыс тапқан, антидене сияқты антигенге көзделген, В-лимфоциттен пайда болған клондармен синтезделетін - яғни моноклинальді антидене.
Моноклинальді антиденелер қасиеттері бойынша біртекті,олар біркелкі уәкілге бірдей туысқандыққа ие болады және жалғыз-ақ қарсы гендік детерминанттармен жалғастырылады.
Адамның иммундық жасушалары негізінде буданды алудың технологиясын игеру түбегейлі қиындықтармен байланысты:адамның гибридомалары баяу өседі, салыстырмалы түрде тұрақсыз. Кемшіліктері:олар кептірілген күйде тұрақсыз және кәдімгі антиденелердің қоспасында сақтаудың таңдамалы шарттарында орнықты келетін антиденелер кездеседі.
Миелом жасушалары мен иммундық лимфоциттер неізінде будандарды алудың жалпы үлгісі келесі кезеңдерден тұрады.
1. Гибридомды жасушалардың келесі селекциясы барысында тіршілігін жоятын мутантты ісік жасушаларын алу.
2. Лимфоциттерді алу - берілген антигенге антидене продуцентін алу. Жануарды (тышқан, сирек егеуқұйрықты, қоян) құрсақ қуысына, көктамырішілік немесе тері астына енгізумен иммунитеттейді. Адамның гиридомаларын алу күрделі және көп сатылы күрделі процедура табылтын ұлпа культурасында адамның лимфоциттерін иммунитеттейді.
3. Лимфоциттердің ісік жасушаларымен қосылуы:қосушы агент болып полиэтиленгликоль, сирек түрде лизолецитин немесе Сендай вирусы, және де қуаттылығы жоғары электр өрісі қолданылады.
4. Гибридомды жасушалар скринингі. Нуклеотидтер синтезінің маңызды жолдарын жауып тастайтын аминоптериннен тұратын селекциялық орта НАТ қолданылады, және биосинтездің қосымша жолдарының алғызаттары болып табылатын гипоксантин мен тимидин де қолданылады. Бұл ортада ата-аналық миелом жасушалары биосинтездің қосыша жолдар ферменттері бойымен генетикалық ақаулы сияқты тіршілігін жояды. Миелом жасушаларымен қосылмаған ата-аналық лимфоциттер де тіршілігін жояды, себебі олар организм сыртында өмір сүре алмайды. Гибридомды жасушалар шексіз өсуге және қосымша жолдар бойынша нуклеотидтер синтезіне бейім, сондықтан олар культурада жинақталады.
5. Гибридомды жасушалардың берілген антигенге моноклональді антидене шығару қабілетін тексеру. Ол үшін иммуносорбенттер әдісі қолданылады. Гибридомды жасушалары бар сұйықтық үлгісін сақтаушыда нық бекітілген антигені бар реакцияға енгізеді. Антиген - антидене кешенін айырып тану үшін - пайдаланылатын антиденеге жануардың осы иммуноглобулиндерін иммундау арқылы екінші антиднелерді алады (мысалы, тышқаның иммуноглобулиндерін ешкінің организміне енгізеді). Бұл екінші антиденелер ковалентті түрде қандай да бір ферментпен байланысады (мысалы, сілтілік фосфатазмен немесе пероксидазамен). Егер гибридомадан өндірілген антидене шынында берілген антигенді байланыстырса, онда оған екінші ферментпен қосылған антеденені қоссақ ол мынандай кешен пайда болуына жағдай жасайды - антиген-моноклональді антидене - антидене моноклональді антиденеге - фермент.
6. Моноклональді антидене өндіруіне байланысты тексеруден өткен және иммундық жүйесінің тұрақтылығына байланысты күнделікті бақылаудын өткен гибридомды жасушаларды клондау.
7. Гибридомаларды жаппай өсіру,шығару және шығарылатын антиденеледі қойылту және тазарту.
Өсімдіктер түрлері мен микроорганизмдер штаммаларының жаңа түрлерін шығару және бар сұрыптарын мейлінше жақсарту.
Жасушалық инженерияның маңызды және песпективалы бағыты -өсімдік түрлері мен микроорганизмдер штаммаларының жаңа түрлерін шығару және бар түрлерін жақсарту. Бірақта протопластардың қосылуы негізінде өсімдіктердің жаңа үрін шығару үшін екі негізгі кедергіден өту керек: а) протопластардың жасушалық қабырғамен қапталған жасушаға айналуы; б)жасушадан тұтас өсімдік алу. Бұл екі негізгі қосымша кедергіден басқа қосымша қиыншылықтар бар. Тұтас дарақтың регенерацияға мүмкіншілігі көрсетіліп, екі ата-аналық протопластта өсімдіктер түріне жатса да қосылған жасушалар осындай регенерацияға қабілетін жиі жоғалтып алады.
Жасушалық ассосациялар. Әртүрлі организмдер жасушаларының ассосациясын жасап шығару жөнінде қарқынды түрде зерттеу жүргізілуде. Жасанды симбиоз құру мақсатында екі немесе оданда көп организм жасушаларының араласқан культурасын пайдаланып, яғни серіктестердің қолайлы өмір сүруіне байланысты зерттеулер. Белгілі табиғи симбиоз ретінде құрамында саңырауқұлақ және балдыр жасушалары араласқан қынаны (немесе цианобактерия) айтуға болады, «цианобактерияның инфузориясы» жүйесі(соңғылары инфузорияның цитоплазмасында өмір сүреді және оны оттегімен қамтамасыз етеді)
Дәріс № 7. Биологиялық объектілердің популяциялық тұрақтылығы.
Бионысананы жұмыс жағдайында ұстап тұру, оның құнды қасиеттерін сақтау маңызды биотехнологиялық мәселе болып табылады.
Биореактор, культивирленген микроорганизмдер немесе өсімдіктер мен жануарлар клеткаларының үлкен популяциясымен қамтылған. Бұл популяцияда 1010 және одан аса жасушалар бар, олар өз кезегінде жер бетіндегілерден миллион есе асады. Бұл жағдайларда екіталай генетикалық мәселеге негізделген процестер үлкен рөл атқарады. Сонымен төменгі жиілікті спонтанды мутациялар (1 мутация 106-108жасушаға) биореакторда мутантты түрлердің тез жиналып қалуына әкеледі. Әсіресе мынадай жағдайда егер де осындай түрлер тез өсу жылдамдығымен сипатталса. Жасушалық популяцияда бейімделген жасушалар азырақ бейімделгендерді шығарып тастауы автоселекция деп аталады, яғни бұл көбінесе селекционердің қатысуынсыз және оның жоспарынан тыс болатын процесс. Бактерияларда 3 форманың (K,S,M) жиі өзара өтуімен сипатталатын фазалық диссоциация көрсеткіштік үлгі болып қызмет етеді. Бұл формалар жасушалық қабырға мен капсуланың құрылысымен өзгешеленеді. Сонымен қатар көптеген физиолого-биохимиялық қасиеттерімен, оның ішінде биологиялық активті заттар синтезіне қабілеттілік те бар.
Жануарлар және өсімдіктер жасушаларын ағзадан тыс культивирлегенде ұқсас мәселелер пайда болады. Жасуша белгілерінің культивирлеу процесі кезінде өзгеруі оның көпклеткалы организмнен қоректік ортасы бар ыдысқа ауыстырған кезде пайда болған қауырт өзгеріспен байланысты.
Үлкен жасушалық популяцияда өтетін мутанттау мен селекцияның спонтанды процестерінің жақсы жақтары болуы мүмкін:
Мақсаты биомасса алу болып табылатын өндірістік процестерде автоселекция продуктивті және көп бейімделгіш формалар басымдығына әкеледі.
Егер зерттеуші келесі селекционды жұмысқа бастапқы формадан өзгешеленетін формалар алуға тырысса, бұл процестерде биотехнологиялық маңызды қасиеттер жоғалса, онда бұл жаман жағы болады. Әдетте генетикалық және жасушалық инженерия негізінде алынатын штаммдар тұрақсыз болып келеді.
Маңызды штамм-продуценттерді сақтау мәселесі мынадай екі жағдайда біріншілік маңыздылық алады: олардың ұзақ сақталуы кезінде және ол штаммдардың зертханалық культиваторлардан өндірістік биореакторға көшірілуі кезінде (масштабты өту және масштабтану).
Микроорганиздер культураларының халықтық әдістері олардың жиі ауыстырылатын бай қоректік ортада өсірілуіне әкелді. Осы кезде пайда болатын мутанттар мен автоселекцияға жағымды жағдайлар көбінесе штамм-продуценттердің жоғалуына әкеп соқтырады. Басқа әдістер пайда болмағанша бұл әдіс қана қол жетімді. Популяцияда қажет емес мутанттардың жиналуының алдын-алуға болады, егер де әр қайта отырғызудан соң жасушалар клондарының функционалды активтілігін тексеретін скрининг өткізсе.
Гибридпен клондау кезінде жасушаның маңызды қасиеттерін сақтау үшін қандай жолдар қарастырылған?
Жасушаларды ұзақ уақыт бойы маңызды қасиеттерін жоғалтпай сақтауға болады, егер де онда болып жатқан барлық өмірлік процестерді тоқтатса,оның ішінде генетикалық қайта құрылуды да. Бұл кезде культура – анабиоз сияқты жағдайға түседі және сақтау әдістері анабиозға жақындау сатысы бойынша жіктеледі:
Жасушаның лиофильді кептірілуі (вакуумда мұздатылғаннан кейін 60ºС және одан төмен температурада обезвоживание). Бұл әдіс –жақсы, мысалы, белсенділігін бірнеше жылға сақтайтын антибиотиктер үшін. Дегенмен бионысаналардың барлығы лиофилизациядан өте алмайды. Лиофилизация жағдайында өміршеңдігі төмен жасушалар өледі де, популяция өмір сүруге қабілеті мол жасушамен толықтанады.
Бұл байланыста осы немесе өзге штамм-продуценттің белсенділігін сақтау үшін селективті жағдайлар қажет.
Стерильді топырақ ауасында, құмда, белсендендірілген көмірде, кейбір өсімдіктердің ұрықтарында, агар-агар дисктерінде, қағазда, жүнді жіпшелерде және өзге тасымалдаушыларда кептіру. Бұл әдіс салыстырмалы түрде қарапайым. Бірақ ол популяцияда болып жататын гентикалық өзгерістерді жеткіліксіз деңгейде тоқтатады. Соңғы жылдары сұйық жағдайдағы вакуум астында кептіру қолданылады. Микроорганизмнің кей түрлері, әсіресе, лиофилизацияны өте алмайтындар осындай өңдеуден кейін өмірге қабілетті жағдайда сақталады. Ал бионысананың көпшілігі кептіруді көтере алмай, өледі.
Споралар сақталуы (спора түзетін бактерияларға жарамды).
Криоконсервация –жасушалардың терең мұздатылуы және сонан соң сұйық азотта (-19º) немесе оның буында (-150º) кептірілуі. Басқа сақтау әдістерінде сақтала алмайтын - цианобактериялар, пурпурлы және жасыл бактериялар, мицелиальді саңырауқұлақтар, актиномицеттер, қарапайымдылар, балдырдар, өсімдік және жануар жасушаларының культуралары, гибридомдар, гендік-инженерлік мутанттарды осы жолмен ұзақ уақыт бойы сақтауға болады. Терең мұздатылған жағдайдағы организмдер жинақтамасы – крио-банкалар жасалынды. Криоконсервация толықтай жасуша популяциясының гендік фондының бұзылуына кедергі болады.
Комбинирленген сақтау әдістері. Кейбір жағдайларда маңызды қасиеттердің жақсы сақталуы бірнеше сақтау әдістерінің комбинациясы арқылы жүзеге асады. Яғни, жасушалардың біртіндеп кептірілуі, келесі лиофилизацияда немесе төменгі температурада сақталуында, олардың өміршеңдігі мен биохимиялық белсенділігін сақтауға септігін тигізеді.