

2.5. Процеси збудження в цнс. Збуджувальні синапси, їх нейромередіатори, розвиток збуджувального постсинаптичного потенціалу, параметри, фізіологічна роль.
Збуджувальними медіаторами можуть бути різні речовини, які спричинюють деполяризацію постсинаптичної мембрани. Найважливіше значення мають похідні глутамінової кислоти (глутамати), субстанція Р. Деякі центральні нейрони мають холінорецептори, тобто містять на постсинаптичній мембрані рецептори, які реагують з холіновими сполуками, наприклад, ацетилхолін у клітинах Реншоу..Збуджувальними медіаторами можуть бути також моноаміни (норадреналін, дофамін, серотонін). Є підстави вважати, що тип медіатора, який утворюється в синапсі, обумовлений не тільки властивостями закінчення, але й загальним спрямуванням біохімічних процесів у всьому нейроні.
Збудження пов'язане з розвитком у нейроні тимчасової деполяризації. Деполяризаційна відповідь нейрона на збудження отримала назву збуджувального постсииаптичного потенціалу (ЗПСП). Внутрішньоклітинно ЗПСП у нейронах ссавців був уперше зареєстрований Д. Екклсом зі співробітниками (1966) за допомогою мїкроелектронної техніки.
На відміну від збудження, що поширюється (ПД), ЗПСП є місцевим. Воно локальне, градуальне (залежить від сили подразнення, тобто не підпорядковується закону «все або нічого»), здатне до сумації з попередньою деполяризацією, не залишає після себе рефрактерності. За своїми характеристиками ЗПСП є ідентичним потенціалу кінцевої пластинки (ПКП) у нервово-м'язовому сполученні . Проте, якщо ПКП є наслідком активації одного синапса, то ЗПСП переважно виникає при одночасній активації кількох синапсів.
ЗПСП досягає максимуму протягом 1,5—2 мс, після чого повільно затухає за експонентою. Загальна тривалість ЗПСП багатьох нейронів ЦНС складає близько 15 мс, а амплітуда може досягати —20—30мВ при величині мембранного потенціалу —70 мВ (мал. 31).
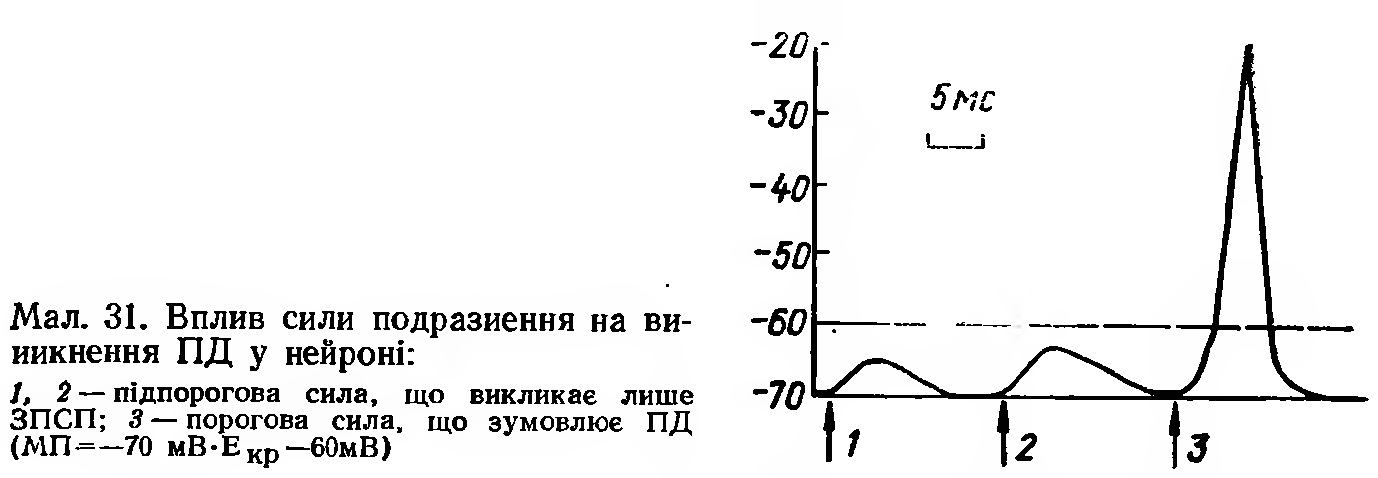
Місцем первинної генерації ПД у нейроні в більшості випадків є початкова частина аксона (аксонний горбик). Висока збудливість цього горбика пояснюється тим, що поріг його деполяризації удвічі менший (15 мВ), ніж у сомі нейрона (30мВ). Це пояснюється тим, що у мембрані аксонного горбика значно більше іонних каналів. То найбільш збудлива ділянка нейрона (так звана тригерна зона), і лише після її збудження імпульс поширюється, з одного боку, на аксон, а з другого — на сому і дендрити. ПД нейрона, що реєструється внутрішньоклітинно, має два компоненти: спочатку реєструється потенціал початкового сегмента, або ПС-потенціал, а потім потенціал соми і дендритів — СД-потенціал.
В основі виникнення збудження лежить трансмембранний рух іонів, серед яких домінуюча роль належить Са2+. Збільшення вмісту кальцію в середовищі і цитоплазмі, вплив антагоністів кальцієвої провідності (ВАYK 8644) призводять до збудження, у той час як блокатори Са2+-каналів (верапаміл, Д-600) гальмують його розвиток. Є вказівки і про можливу участь у генерації ЗПСП Nа+та Cl-.
Збудження нейрона супроводжується змінами метаболізму, зокрема синтезу РНК, та іншими зрушеннями в процесі білкового синтезу, посиленням теплопродукції, поглинанням кисню, які відображають інтенсифікацію життєдіяльності клітини.
Однобічне проведення.На відміну від нервового волокна, здатного проводити збудження в обох напрямках (див. с. 21), в центральному синапсі воно поширюється тільки в одному: від рецептора через вставні нейрони до ефектора.
Це зумовлено тим, що медіатор, звільнений пресинаптичним закінченням,. надходить у синаптичну щілину, діє на рецептори постсинаптичної мембрани, викликаючи там ЗПСП, а потім ПД у постсинаптичній структурі, тобто синапс функціонує як клапан.
Сповільнене проведеннязбудження. У синапсах ЦНС збудження проводиться повільніше, ніж у нервових волокнах. Це пояснюється особливостями синаптичної передачі (так звана синаптична затримка), а саме дифузією медіатора до постсинаптичної мембрани, виникненням ЗПСП, наростанням його до критичної величини, генерацією ПД. Точні вимірювання дозволили встановити, що для проведення збудження через один синапс потрібно 2—3 мс. Виходячи з цього, можна визначити кількість синапсів у нейронній сітці, через які проходить імпульс у ЦНС.
Трансформація ритму збудження. Здатність нервових центрів змінювати (переробляти) ритм імпульсів, які надходять до них, називається трансформацією ритму. Найчастіше трансформація ритму проявляється в тому, що у відповідь на поодинокий стимул-подразнення нервові центри надсилають до виконавчого органа (ефектора) цілий ряд імпульсів, які надходять один за одним з певною частотою. Цим пояснюється й виникнення тетанічного скорочення скелетного м'яза у відповідь на поодиноку стимуляцію відповідного аферентного нерва. Образно кажучи, у відповідь на поодинокий постріл центр відповідає кулеметною чергою. В основі цього явища лежать головним чином слідові коливання мембранного потенціалу, особливо тривалість слідового негативного потенціалу (див. с. 13). Якщо слідовий негативний потенціал, який супроводжує ПД, великий, то він може досягнути критичного рівня деполяризації мембрани і зумовити появу другого імпульсу. У ряді випадків трансформація спричинюється збільшенням тривалості ЗПСП, і після закінчення першого ПД розвиваються наступні.
Сумація збуджень. Під сумацією збуджень на тілі нейрона розуміють сумацію ЗПСП як місцевого потенціалу. Додавання — процес складання двох або більше однакових реакцій, унаслідок якого результуюча реакція буде більшою за своєю величиною, ніж кожна із її складових.