

ФІЗІОЛОГІЯ ЛЮДИНИ
Лекція № 2 Фізіологія скелетних і гладких м’язів, опорно-рухового апарату. Фізіологія нейрона. Збудження та гальмування в ЦНС. Інтегративна функція нейронних ланцюгів
2.1. Фізіологія м’язів, будова м’язового волокна. Механізми скорочення та розслаблення скелетних м’язів. Будова та функції нервово-м’язового синапсу. Функції й властивості скелетних м’язів. Типи м’язових волокон.
2.2. Типи скорочення м’язів залежно від зміни їхньої довжини та напруження: ізометричні, ізотонічні. Фізіологія та властивості гладких м’язів. Автоматія.
2.3. Нейрон та нейроглія як структурно-функціональні одиниці ЦНС, їхні види, функції. Нейронні ланцюги та нервові центри. ( СПРС)
2.4. Синапси ЦНС, будова, механізм передачі інформації. Нейромедіатори, їхні види. Гальмівні синапси, їх нейромедіатори. Постсинаптичне та пресинаптичне гальмування, механізм розвитку. Види постсинаптичного гальмування.
2.5. Процеси збудження в ЦНС. Збуджувальні синапси, їх нейромередіатори, розвиток збуджувального постсинаптичного потенціалу, параметри, фізіологічна роль.
Самостійна робота: Нейрон та нейроглія як структурно-функціональні одиниці ЦНС, їхні види, функції. Нейронні ланцюги та нервові центри.
2.1. Фізіологія м’язів, будова м’язового волокна. Механізми скорочення та розслаблення скелетних м’язів. Будова та функції нервово-м’язового синапсу. Функції й властивості скелетних м’язів. Типи м’язових волокон.
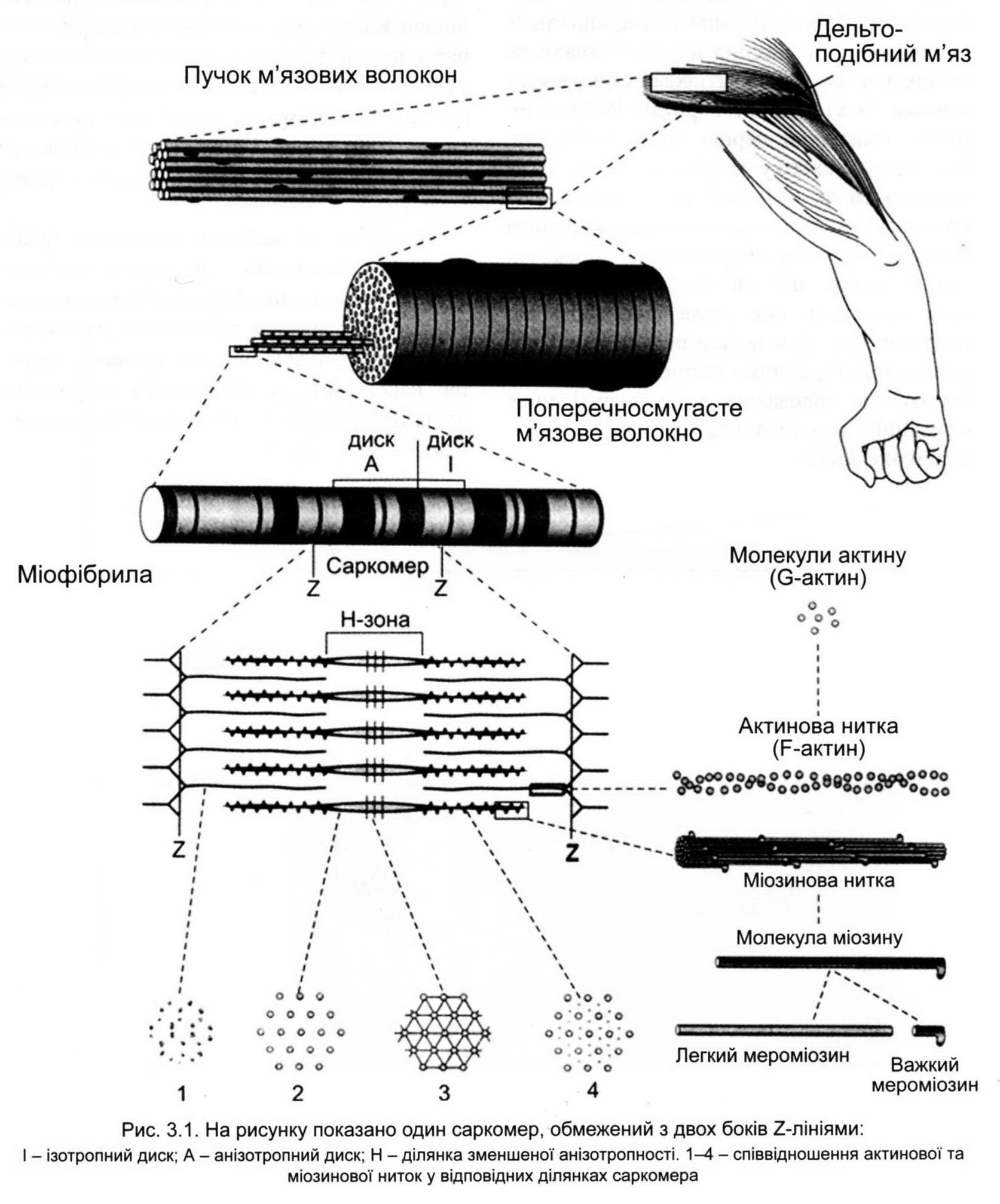
Основний структурний елемент скелетного м'яза - м'язове волокно - виникає внаслідок злиття багатьох клітин. Одне м'язове волокно в діаметрі не перевищує 0,1 мм, а довжина його може складати від декількох міліметрів до 12 см. Кожен м'яз складається з різної кількості таких волокон (рис. 3.1).
Під світловим мікроскопом у м'язовому волокні видно чергування темних і світлих смуг (поперечна посмугованість).Темні диски мають подвійне променезаломлення, а світлі диски цієї властивості не мають, що і створює поперечну покресленість м'яза. Частина м'язового волокна від середини одного диска І до середини іншого називається саркомером. Довжина саркомера у спокійному м'язі близько 2 мкм, а в скороченому з максимальною силою - близько 1 мкм, тобто м'яз максимально може скорочуватися на 50-60 % від своєї початкової довжини.
Що собою являють мембрана і цитоплазма м'язового волокна?
Мембрана м'язового волокна - сарколема (від грецьк. sаrх, sаrkоs - м'ясо) утворена типовою плазматичною мембраною, укріпленою сполучнотканинними волокнами. Ці волокна, об'єднуючись біля кінців м'язових волокон, формують сухожилки, за допомогою яких м'яз кріпиться до кісток.
У цитоплазмі (саркоплазмі) м'язового волокна, крім типового набору органоїдів, знаходиться саркоплазматичний ретикулум (СР), що являє собою депо кальцію. Крім того, необхідно згадати про наявність у саркоплазмі важливого білка міоглобіну, який служить депо кисню всередині волокна (він надає м'язу червоного кольору).
Які структури м'язового волокна забезпечують його скорочення?
Усередині м'язового волокна в саркоплазмі впорядковано розташовуються скоротливі протофібрили. Розрізняють протофібрили двох типів: товсті (товщина 15-17 нм) і тонкі (товщина близько 6 нм). Тонкі протофібрили розташовані в І-зоні і являють собою білкові актинові нитки. Товсті нитки, що знаходяться в зоні А, іменуються міозиновими.
Скорочення
В результаті сполучення вказаних процесів відбувається: а) підтягання ниток міозину до актинових; б) зарядка міозину енергією, що використовується для виконання повороту міозинової головки. Після цього утворені Ф- і АДФ- відходять, а на їх місце приєднується нова молекула АТФ, що призводить до розриву зв'язку міозину з активним центром актину.
У даний
час тонкий механізм, що забезпечує
входження актоміозинових ниток одна в
одну, точно ще не відомий. Загальноприйнятою
сьогодні є гіпотеза "шарнірного
механізму" (рис.
3.5). Після з'єднання головки міозину з
активним центром актину відбувається
поворот головки на 45°. В результаті
розриву містка шийка головки міозину
випрямляється, займаючи початкове
положення. За ці рухи дана система і
отримала назву "шарнірного механізму".
При повороті міозин просувається по
актину на один "крок" або "гребок",
рівний 20 нм (близько 1 % довжини
саркомера).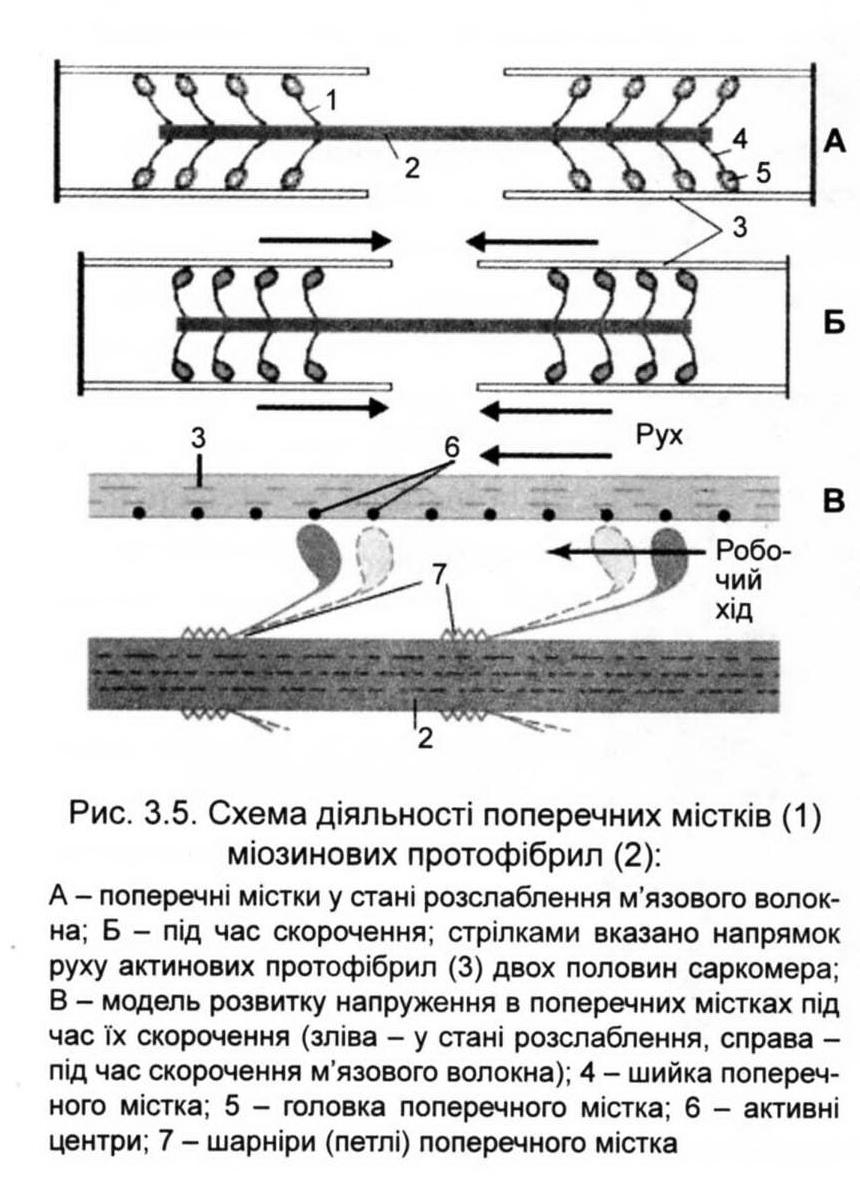
За наявності високої концентрації Са2+ "кроки" повторюються, але тепер уже якоїсь іншої головки, що опинилася навпроти нового активного центру актину, оскільки активні центри розташовані на відстані близько 40 нм один від одного. Оскільки подібні "кроки" відбуваються з обох боків саркомера (від 2-мембрани), то м'яз коротшає до свого центру.
Які процеси забезпечують розслаблення м 'яза?
Доти, поки в саркоплазмі є вільний Са2+ (у концентрації більше 10~5М) і АТФ, повторюватиметься взаємодія актинових і міозино-вих міофіламентів. Якщо немає нової хвилі деполяризації, то кальцій швидко забирається назад у цистерни СР. Він відкачується із саркоплазми проти градієнта концентрації за допомогою Са-насоса, розташованого на мембрані СР. Робота насоса активується самим кальцієм, вірніше, зростанням його концентрації в саркоплазмі, і вимагає витрати великої кількості АТФ: для видалення кожного іона Са2+ використовується 2 молекули АТФ. Результатом відкачування кальцію із саркоплазми є розрив усіх зв'язків актину і міозину і розслаблення м'яза.
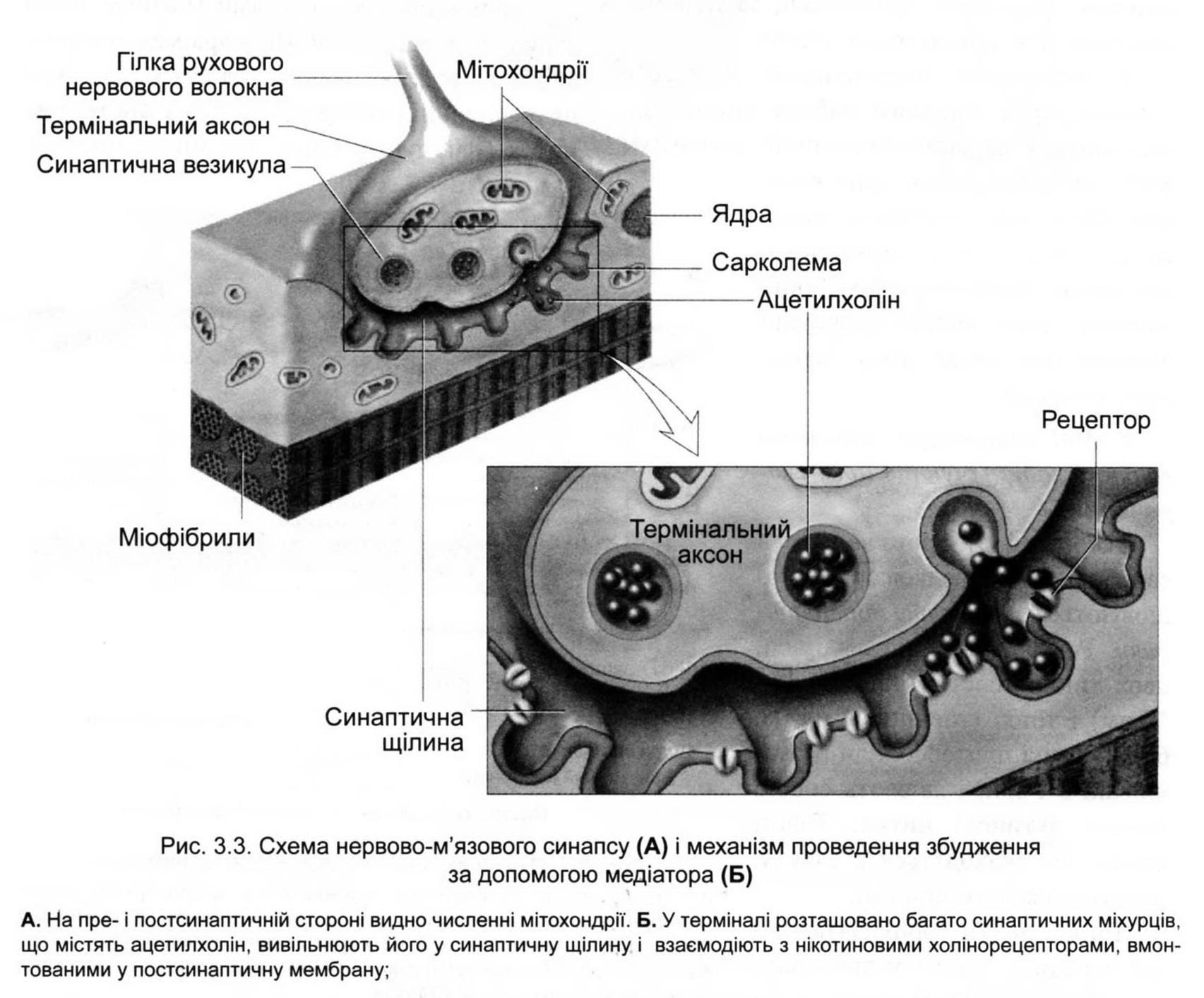
Нервово-м'язовий синапс має наступну будову (рис. 3.3). Простір шириною 20-30 нм між мембранами нервового закінчення та іннервованим м'язом іменується синоптичною щілиною. Мембрана нервового закінчення є пресинаптичною, а відповідна їй частина мембрани м'язового волокна -постсинаптичною (ця ділянка мембрани ще називається кінцевою пластинкою). Нервове волокно закінчується пресинаптичним розширенням. Усередині нього є велика кількість (до 300 000) синаптичних бульбашок (діаметром близько 50 нм), що містять певну кількість хімічної сполуки ацетилхоліну (АХ). Це хімічний передавач збудження -медіатор. Кожна бульбашка містить "квант" медіатора - близько 104 молекул АХ. Синаптична щілина заповнена рідиною, яка за своїм складом нагадує плазму крові.
Які структури сарколеми забезпечують сприймання регулюючого сигналу?
Медіатор на постсинаптичній мембрані взаємодіє з рецептором. Рецептор - це білковий комплекс, який сприймає сигнал молекули-передавача (біологічно активної сполуки - ліганда) - ацетилхоліну. Такий рецептор називається холінорецептором (ХР).
Але необхідно враховувати, що, крім медіатора, на рецептор можуть впливати і інші сполуки, що мають до нього спорідненість. При цьому можуть бути два взаємно протилежних ефекти: відбудеться блокада рецептора (така сполука іменується літиком), або, навпаки, - імітуватиметься ефект взаємодії природної молекули-передавача з рецептором (така сполука іменується міметиком).
Яким чином відбувається передача збудження у нервово-м'язовому синапсі?
Потенціал дії, що прийшов до синапсу, проходячи по пресинаптичній мембрані, відкриває електрозбудливі кальцієві канали. Іони Са2+ по градієнту концентрації надходять всередину синаптичної бляшки, що призводить до просування синаптичних бульбашок до пресинаптичної мембрани. Потім мембрана везикул зливається з мембраною нервового закінчення, і медіатор шляхом секреції викидається в синаптичну щілину. Медіатор дифундує по синаптич-ній рідині, і велика частина його молекул досягає постсинаптичної мембрани, де і взаємодіє з холінорецептором. Зазвичай для передачі одного ПД вивільняється до мільйона молекул АХ (200-300 везикул). Результатом взаємодії АХ з ХР є відкриття хемозбудливих іонних каналів. Селективна ділянка його має діаметр 0,65 нм. Через нього можуть проходити лише позитивні іони (стінка каналу електронегативна) натрію або кальцію. Але в нормі, як правило, переважає потік іонів натрію. Вони по концентраційному градієнту із синаптичної щілини надходять всередину м'язового волокна і де поляризують постсинаптичну мембрану.
2.2. Типи скорочення м’язів залежно від зміни їхньої довжини та напруження: ізометричні, ізотонічні. Фізіологія та властивості гладких м’язів. Автоматія.
Які є типи і режими м 'язових скорочень?
Під час скорочення м'яз тягне обидва своїх кінці до центру. У природних умовах обидва кінці м'яза прикріплюються за допомогою сухожиль до кісток і при скороченні притягують їх одна до одної. І якщо один кінець м'яза (суглоб) закріплений, то до нього підтягується інший.
Коли м'яз не в змозі підняти вантаж, то тоді він лише напружується без зміни довжини. Зустрічаються і такі стани, коли м'яз поступово збільшується у довжину (вага вантажу перевищує підйомну силу м'яза, або необхідно повільно опустити вантаж). Через це розрізняють наступні типи м'язових скорочень: ізотонічне (концентричне) - скорочення м'язів з укороченням при збереженні постійного напруження, ізометричне - довжина м'яза залишається без змін; ексцентричне - коли м'яз подовжується. Як правило, більшість природних скорочень м'язів є змішаними - ауксотонічного типу, коли м'яз коротшає при підвищенні напруження.
Гладенькі м'язи знаходяться в стінках внутрішніх органів, судин, шкірі. Структурною одиницею їх є витягнутої форми клітина: довжиною 20-400 мкм, завтовшки 2-Ю мкм. На мембрані гладком'язових клітин, на відміну від скелетних, є не тільки натрієві і калієві канали, але й велика кількість кальцієвих каналів.
На схемі (рис. 3.10) показана компоновка скоротливих міофіламентів усередині клітини. Актинові філаменти згруповані в пучки, які час від часу утворюють ущільнення ("вузли"). Деякі з них безпосередньо прилягають до мембрани, інші знаходяться усередині клітини, виконуючи як би функцію 2-мембран. Між актиновими філаментами вкраплені товстіші - міозинові. Прилеглі до мембрани ділянки двох сусідніх клітин часто співпадають і утворюють тут білкову перемичку, що об'єднує структури цих клітин у синцитій.
З фізіологічної точки зору доцільне виділення двох типів гладком'язових клітин: а) розташовані окремо; б) такі, що створюють функціональний синцитій .
Чим відрізняються два типи гладком 'язових клітин?
Окремо розташовані гладком'язові клітини знаходяться в структурах ока (рай-дужка, війкові), артеріол, сім'яних проток, навколо волосяних цибулин, іх мембрана містить колагенові і глікопротеїнові волокна, які додатково відокремлюють клітини одна від одної. Ці клітини іннервуються вегетативними нервами подібно до скелетних м'язових волокон. Медіатори, що вивільняються з нервових закінчень, викликають препотенціали, що при сумації переходять у потенціали дії. Це призводить до скорочення м'язових клітин. У стінці більшості внутрішніх органів, судин є другий тип гладком'язових клітин, їх мембрани тісно стикаються одна з одною, деколи утворюючи загальні іонні канали (нексуси). Завдяки цьому окремі клітини об'єднуються, утворюючи функціональний синцитій.
Як відбувається скорочення гладком'язових клітин?
У цитоплазмі гладком'язових клітин знаходяться актинові і міозинові філаменти, які самі по собі структурно і функціонально мало відрізняються від відповідних утворень скелетного м'яза. Проте розташовані вони не так впорядковано, як у скелетному м'язі: тут немає типових 2-мембран. Крім того, в гладенькому м'язі менше міозинових волокон. Відрізняються ці два типи м'язів і тим, що в гладком'язовій клітині слабо виражений саркоплазматичний ретикулум.
Механізм скорочення гладеньких м'язів приблизно такий, як і у скелетних, але відрізняється деякими деталями. Ініціатором взаємодії актину і міозину також є кальцій, критична концентрація розслаблення якого - близько 10 8 моль/л. Але після підвищення концентрації кальцію всередині саркоплазми він разом з кальмодуліном активує особливий фермент - кіназу легких ланцюгів міозину. Кіназа переносить фосфатну групу АТФ на міозин (відбувається гідроліз АТФ), що і забезпечує взаємодію міозину з актином, тобто м'язове скорочення. Певну роль при цьому відіграє і цАМФ.
Для розслаблення м'яза необхідно "відкачати" вільний кальцій із цитоплазми. Проте в гладком'язовій клітині це відбувається повільніше, ніж у скелетній. Тому як скорочується, так і розслабляється вона повільніше.
У типових клітинах скорочення починається через 50-100 мс після збудження. Повна хвиля скорочення розвивається близько 0,5 с і триває протягом 1-3...30 с.
Ініціатори скорочення (іони кальцію) надходять всередину волокна двома шляхами: з міжклітинної рідини, коли відкриваються відповідні канали при проходженні ПД, і з саркоплазматичного ретикулуму.
Чим механічні властивості гладеньких м'язів відрізняються від властивостей поперечносмугастих?
• Гладенький м'яз скорочується повільніше.
• При однаковій площі поперечного перетину гладенькі м'язи не поступаються, а можуть навіть перевищувати скелетні м'язи за силою скорочення.
• Гладенькі м'язи можуть скорочуватися і розслаблятися в більшому діапазоні (тобто більше ніж удвічі), ніж скелетні.
• Гладенькі м'язи можуть працювати в режимі сфінктера.
Автоматія. ПД гладких мышечных клеток имеют авторитмический (пейсмекерный) характер, подобно потенциалам проводящей системы сердца. Пейсмекерные потенциалы регистрируются в различных участках гладкой мышцы. Это свидетельствует о том, что любые клетки висцеральных гладких мышц способны к самопроизвольной автоматической активности. Автоматія гладких мышц, т.е. способность к автоматической (спонтанной) деятельности, присуща многим внутренним органам и сосудам.