

18.2. Хімічні основи виникнення та проведення нервових імпульсів
18.2.1. Роль нейромедіаторів у передачі нервових імпульсів. Зв’язок мільярдів нейронів мозку здійснюється за допомогою численних медіаторів (нейротрансміттерів): ацетилхоліну (Ах), норадреналіну (НА), дофаміну, адреналіну, серотоніну, гістаміну, ГАМК тощо. Одним з основних медіаторів у нервовій системі є ацетилхолін.
Ацетилхолін — це складний ефір оцтової кислоти та холіну. Він синтезується в нервовій клітині з холіну й активної форми ацетату — ацетил-КоА за допомогою спеціального ферменту холінацетилтрансферази (холінацетилази).
Передача імпульсів з нейрона на нейрон або з нейрона на ефекторний орган відбувається за допомогою синапса. Синапс можна уявити собі як вузький простір (щілину), обмежений з одного боку пресинаптичною, а з іншого – постсинаптичною мембраною (рис.18.4). Пресинаптична мембрана складається з внутрішнього шару, що належить цитоплазмі нервового закінчення, і зовнішнього, утвореного нейроглією. Мембрана в деяких місцях потовщена й ущільнена, в інших – тонша і має отвори для сполучення цитоплазми аксона з синаптичним простором. Постсинаптична мембрана менш щільна, не має отворів. Подібним чином побудовані і нервово-м’язові синапси, але вони мають складнішу будову мембранного комплексу.
Передачу нервового імпульсу за участі Ах можна відобразити наступним чином. У синаптичних нервових закінченнях є міхурці (везикули) діаметром 30 – 80 нм, що містять нейромедіатори.
Ц
Рис.
18.4. Схематичне зображення синапса: 1 –
вивільнення нейромедіатора (Ах) з
синаптичних міхурців під впливом
потенціалу дії; 2 – нейромедіатор (Ах)
дифундує в синаптичну щілину; 3 –
внаслідок взаємодії нейромедіатора
(Ах) з рецептором відкриваються натрієві
канали; 4 – синаптичні міхурці; 5 –
пресинаптичне закінчення: 6 – синаптична
щілина; 7 – постсинаптична мембрана; 8
- нейромедіатор (Ах), 9 – зв’язування
нейромедіатора з рецептором; 10 – іони
натрію.

Взаємодія між рецептором і медіатором запускає низку реакцій, які змушують постсинаптичну нервову клітину або ефекторну клітину виконувати свою специфічну функцію.
Рецептор Ах – це трансмембранний олігомерний глікопротеїновий комплекс, який складається з 6 субодиниць: 2 – альфа, 2 – бета, 1 – гама і 1 – дельта. У присутності Ах бокові взаємодії між субодиницями підтримують канал у відкритому стані та створюють умови для транспорту іонів. За відсутності Ах змінюється орієнтація субодиниць і канал закривається. Щільність розташування білків-рецепторів у постсинаптичній мембрані дуже велика – біля 20 000 молекул на 1 мкм2. Просторова структура рецептора строго відповідає конформації медіатора. При взаємодії з Ах білок-рецептор так змінює свою конформацію, що,як зазначалося вище, всередині нього формується натрієвий канал. Його катіонна селективність обумовлена тим, що ворота каналу сформовані негативно зарядженими амінокислотами. У такий спосіб підвищується проникність постсинаптичної мембрани для натрію, виникає новий імпульс (або скорочення м’язового волокна). Деполяризація постсинаптичною мембрани викликає дисоціацію комплексу «Ах-білок-рецептор» і Ах вивільняється в синаптичну щілину, де за 40 мкс гідролізується.
Розщеплення Ах у холінергічних синапсах відбувається двома шляхами: перший –ферментативний, другий – енергозалежний активний транспорт Ах у нейрон, де він нагромаджується для наступного повторного використання.
Гідролітичний розпад Ах на оцтову кислоту і холін каталізується ферментом, що отримав назву ацетилхолінестераза.
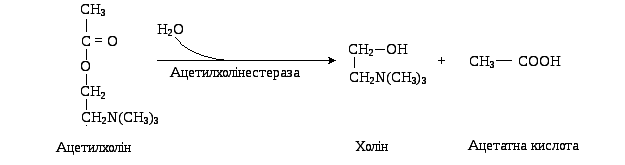
Незворотне інгібування ацетилхолінестерази викликає смерть внаслідок зупинки дихання. Інгібіторами цього фермента є фосфорорганічні сполуки (хлорофос, дихлофос, табун, зарин, зоман, бінарні отрути), які ковалентно зв’язуються з серином у його активному центрі. Зворотні інгібітори ацетилхолінестерази використовують у якості лікарських препаратів для лікування глаукоми та атонії кишок.
У нервовій тканині існують й інші естерази, які можуть гідролізувати Ах, але значно повільніше, ніж, наприклад, бутирилхолін. Ці естерази називаються псевдохолінестеразами.
До числа холінергічних систем належать моторні нейрони, що утворюють нервово-м’язові веретена, усі прегангліонарні нейрони автономної нервової системи і постгангліонарні нейрони парасимпатичної нервової системи. Велика кількість холінергічних рецепторів виявлено також у різних відділах головному мозку. У залежності від чутливості до тої чи іншої групи хімічних сполук, холінергічні нейрони поділяють на мускаринові (активуються мускарином) і нікотинові (активуються нікотином) (табл. 18.4).
Таблиця 19.3.