

2 Глутатион-sh МетНb Hb витамин с
Рис.2 Значение ПФП
Недостаточность глюкозо-6-фосфатдегидрогеназы является наиболее частым дефектом ферментативных систем эритроцита. При низкой активности этого фермента возникает дефицит НАДФН∙Н+, что ограничивает функцию глутатионредуктазы, т.е. нарушается восстановление глутатиона. Падение концентрации глутатиона-SH влечёт за собой снижение активности глутатионпероксидазы. Это приводит к накоплению продуктов ПОЛ, следствием чего являются метгемоглобинемия и, поскольку избыток перекисных продуктов способен разрушить эритроцитарную мембрану, – гемолитическая анемия
Надо заметить, что НАДФН∙Н+, в отличие от НАДН∙Н+, не участвует в окислительном фосфорилировании, протекающем в митохондриях, и не служит, таким образом, для получения энергии.
Велико значение пентозофосфатного пути как поставщика рибозы-5-фосфата, необходимого для построения мононуклеотидов (АМФ, АДФ, АТФ, ГМФ и т.д.), олигонуклеотидов, коферментов (ФМН, ФАД, НАД, НАДФ), нуклеиновых кислот.
Апотомический путь тесно связан с гликолизом. В зависимости от тех или иных условий оба пути могут переключаться друг на друга, поскольку у них имеются общие метаболиты (например, фруктозо-6-фосфат, 3-ФГА). Однако в отличие от гликолиза в пентозофосфатном пути используется другой кофермент (НАДФ+ вместо НАД+), в нём образуется СО2 (чего нет в гликолизе) и он не обладает энергетической функцией.
Регуляция.
Главными регулируемыми (ключевыми) ферментами пентозофосфатного пути являются 2 дегидрогеназы его окислительной части: глюкозо-6-фрсфатдегидрогеназа и дегидрогеназа 6-фосфоглюконата. Индукторами биосинтеза этих ферментов является инсулин. Активность дегидрогеназ увеличивается при поступлении углеводов в организм и снижается при голодании и диабете. Именно поэтому они считаются адаптивными ферментами
2.Глюконеогенез – механизм синтеза глюкозы.
Глюконеогенез – синтез глюкозы из неуглеводных соединений: ПВК, молочной кислоты; аминокислот, распадающихся до пирувата (в первую очередь, аланина, а также цистеина, глицина, серина, треонина, триптофана); глицерина, субстратов цикла Кребса (оксалацетата и других -).

Глюконеогенез является главным метаболическим путём, в котором синтезируется глюкоза. На образование 1 молекулы глюкозы затрачивается 2 молекулы ПВК.
Глюконеогенез протекает в печени и корковом веществе почек. В сущности, реакции глюконеогенеза протекают в обратном направлении реакциям гликолиза – за одним важным исключением: требующие затрат энергии стадии гиколиза (гексокиназная, фосфофруктокиназная и пируваткиназная) не могут обратиться вспять. Вместо указанных киназ «работают» другие ферменты:
|
Ферменты гликолиза |
Ферменты глюконеогенеза |
|
Гексокиназа |
Глюкозо-6-фосфатаза |
|
Фосфофруктокиназа-1 (ФФК-1) |
Фруктозо-1,6-бифосфатаза |
|
Пируваткиназа |
|
В глюконеогенезе – 11 реакций, однако только 4 из них (пируваткарбоксилаза, фосфоенолпируваткарбоксикиназа, бифосфатаза и глюкозо-6-фосфатаза) считеются истинными фермнтами этого пути.
Пируват является конечным продуктом гликолиза и исходной точкой глюконеогенеза.
Рассмотрим начальный этап глюконеогенеза, идущий в обход пируваткиназной реакции гликолиза. Этот этап включает: 1/ две ферментативные реакции, катализируемые ферментами глюконеогенеза пируваткарбоксилазойифосфоенолПВК-карбоксикиназой, 2/ реакции взаимопревращений оксалацетата в яблочную кислоту.
Пируваткарбоксилазная реакция протекает в митохондриях, мембрана которых непроницаема для образующегося оксалацетата (ЩУК). Однако, превращаясь в яблочную кислоту при участии митохондриальной НАД+-зависимой малатдегидрогеназы, малат легко покидает митохондрию и в цитозоле клетки окисляется в ЩУК при участиицитоплазматической НАД+-зависимой малатдегидрогеназы. Дальнейшее превращение ЩУК в фосфоенолПВК происходит в цитозоле клетки.
Малатдегидрогеназа
НАД+
Рис.3 Последовательность реакций глюконеогенеза
С момента образования фосфоенолПВК все реакции, вплоть до образования фруктозо-1,6-бифосфата, идут в направлении, обратном гликолизу. Превращение фруктозо-1,6-бифосфата во фруктозо-6-фосфат катализируется фруктозобифосфатазой.Затем следует реакция, обратная гликолизу. Наконец, последняя реакция глюконеогенеза – фосфатазная. Глюкозо-6-фосфат гидролизуется до глюкозы ферментомглюкозо-6-фосфатазой. Глюкозо-6-фосфатаза – важнейший фермент, ответственный за образование глюкозы из глюкозо-6-фосфата в печени и почки. Именно эти органы являются основными поставщиками глюкозы для тканей организма. Глюкозо-6-фосфатаза практически отсутствует в мышцах – миоциты получают глюкозу из крови. Кроме того, в мышцах синтезируется глюкозо-6-фосфат из глюкозо-1-фосфата в процессе распада гликогена. Подчеркнём, что между гликолизом, протекающим в мышцах при их интенсивной работе, и глюконеогенезом, осуществляемым печенью, существует тесная взаимосвязь (цикл Кори): образующая в мышцах молочная кислота поступает в общий кровоток, захватывается печенью и используется ею в качестве субстрата глюконеогенеза; синтезируемая при этом глюкоза отдаётся в кровототок и метаболизируется мышцами для получения энергии (рис. 6.10 ).
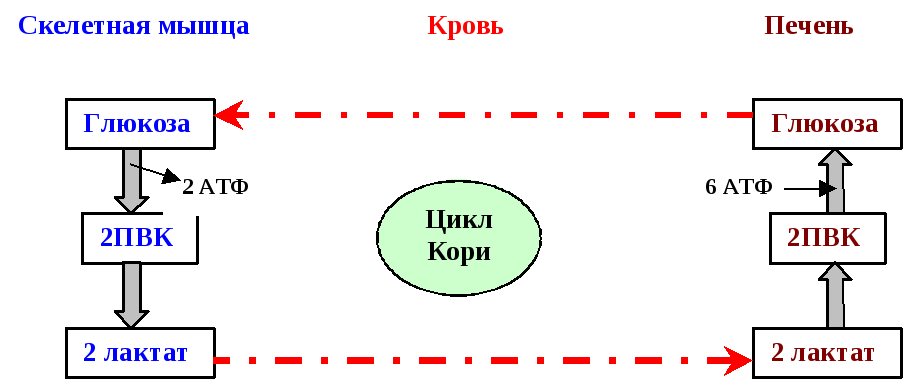
Рис.4. Взаимосвязь между процессами гликолиза и глюконеогенеза.