

Скорость реакций цикла Кребса определяется энергетическими потребностями клетки.
Скорость реакций цикла Кребса коррелирует с интенсивностью процесса тканевого дыхания и связанного с ним окислительного фосфорилирования. В свою очередь, функцио(дыхательного контроля): чем меньше это соотношение, тем интенсивнее идёт тканевое дыхание и эффективнее генерируется электро-химический потенциал, т. е. увеличивается выработка АТФ. Ускорение тканевого дыхания приводит к более быстрому потреблению субстратов цикла Кребса, которые одновременно являются также субстратами тканевого дыхания.
Непосредственно регулируется активность дегидрогеназ цикла Кребса, которая зависит от величины отношения НАДН∙Н+/ НАД+: при недостатке НАД+ их активность снижается, поскольку именно окисленная форма кофермента является акцептором водородов для субстратов лимоннокислого цикла. Помимо этого, существует аллостерический механизм регуляции: для изоцитрадегидрогеназы аллостерическими ингибиторами являются АТФ и НАДН∙Н+, активатором – АДФ. Сукцинатдегидрогеназа угнетается избытком субстрата.

Рис.5. Цикл трикарбоновых кислот (цикл Кребса)
Эффект Пастера – подавление гликолиза дыханием впервые было обнаружено у микроорганизмов.
Заключение. Т.о. в отсутствие кислорода клетка может синтезировать АТФ только за счет гликолитического разрушения глюкозы, и дает незначительное количество энергии. При аэробном окислении глюкозы АТФ образуется исключительно за счет окислительного фосфорилирования.
доц.кафедры
Дата
Министерство здравоохранения Республики Беларусь
Учреждение образования
«Гомельский государственный медицинский университет»
Кафедра биологической химии
Обсуждено на заседании кафедры (МК или ЦУНМС)
Протокол № _______
ЛЕКЦИЯ
По биологической химии
для студентов 2-го курса лечебного факультета
Тема: Углеводы 3. Тканевой обмен __углеводов. Регуляция уровня глюкозы в крови
Время 90 мин
Учебная цель:
Сформировать представления о путях метаболизма глюкозы, молекулярных и физиологических механизмах регуляции уровня глюкозы в крови.
ЛИТЕРАТУРА
1.Биохимия человека:, Р.Марри, Д.Греннер, П.Мейес, В.Родуэлл.- М.книга ,2004.- т.1..с.- 196-204., 210-211., 212-224., 311-318.
2.Основы биохимии:А.Уайт, Ф.Хендлер,Э.Смит, Р.Хилл, И.Леман.-М. книга,
1981,т. 2, 541-608,.с.641-646
3.Наглядная биохимия: Кольман., Рем К.-Г-М. книга 2004г. –с. 50.,154 -157., 160-162., 330.
4.. Биохимические основы ...под. ред. член- корр. РАН Е.С. Северина
М.Медицина,2000.-с.156-158.
МАТЕРИАЛЬНОЕ ОБЕСПЕЧЕНИЕ
1.Мультимедийная презентация
РАСЧЕТ УЧЕБНОГО ВРЕМЕНИ
|
№п/п |
Перечень учебных вопросов |
Количество выделяемого времени в минутах |
|
1. |
Пути обмена глюкозо-6 фосфата. Пентозный цикл |
30 мин |
|
2. |
Глюконеогенез. . Регуляция уровня глюкозы в крови |
30 мин |
|
3. |
Биосинтез ГАГ. |
30 мин
|
Всего 90 минут
Введение. Глюкозо-6 фосфат используется клеткой для биосинтеза восстановленной формы NADH*H и биосинтеза сложных соединений, включая углеводы.
Обмен углеводов подвержен сложным механизмам регуляции, в которых участвуют гормоны, метаболиты и коферменты.
1.Пентозофосфатный путь окисления глюкозы обслуживает восстановительные синтезы в клетке.
Как видно из названия, в этом пути образуются столь необходимые клетке пентозофосфаты. Поскольку образование пентоз сопровождается окислением и отщеплением первого углеродного атома глюкозы, то этот путь называется также апотомическим (apex – вершина).
Пентозофосфатный путь можно разделить 2 части: окислительную и неокислительную. В окислительной части, включающей 3 реакции, образуются НАДФН∙Н+ и рибулозо-5-фосфат. В неокислительной части рибулозо-5-фосфат превращается в различные моносахариды с 3, 4, 5, 6, 7 и 8-ью атомами углерода; конечными продуктами являются фруктозо-6-фосфат и 3-ФГА.
Окислительная часть.
Первая реакция – дегидрирование глюкозо-6-фосфата глюкозо-6-фосфатдегидрогеназой с образованием δ-лактона 6-фосфоглюконовой кислоты и НАДФН∙Н+ (НАДФ+ – кофермент глюкозо-6-фосфатдегидрогеназы).

Вторая реакция – гидролиз 6-фосфоглюконолактона глюконолактонгидролазой. Продукт реакции – 6-фосфоглюконат.
глюконолактонгидролаза

 6-фосфоглюконолактон
6-фосфоглюконат
6-фосфоглюконолактон
6-фосфоглюконат
Н2О
Третья реакция – дегидрирование и декарбоксилирование 6-фосфоглюко-нолактона ферментом 6-фосфоглюконатдегидрогеназой, коферментом которого является НАДФ+ . В ходе реакции восстанавливается кофермент и отщепляется 1С глюкозы с образованием рибулозо-5-фосфата.
СООН
│
 Н–С–ОН
СН2ОН
Н–С–ОН
СН2ОН
│ СО2 │
Н–С–ОН С=О
│ │
Н–С–ОН 6-фосфоглюконатдегидрогеназа Н–С–ОН
│ (декарбоксилирующая) │
Н–С–ОН Н–С–ОН
│ │
СН2ОРО3Н2 СН2ОРО3Н2
6-фосфоглюконат Рибулозо-5-фосфат
Неокислительная часть.
В отличие от первой, окислительной, все реакции этой части пентозофосфатного пути обратимы.
Рибулозо-5-фосфат может изомеризоваться (фермент – кетоизомераза) в рибозу-5-фосфат и эпимеризоваться (фермент – епимераза) в ксилулозо-5-фосфат. Далее следуют 2 типа реакций: транскетолазная и трансальдолазная.
Транскетолаза (кофермент – тиаминпирофосфат) отщепляет 2С-фрагмент и переносит его на другие сахара (см. схему). Трансальдолаза способна переносить 3С-фрагменты.
В реакцию вначале вступают рибозо-5-фосфат и ксилулозо-5-фосфат. Это – транскетолазная реакция: переносится 2С-фрагмент от ксилулозо-5-фос-фата на рибозо-5-фосфат.
 Рибозо-5-фосфат
Ксилулозо-5-фосфат
Рибозо-5-фосфат
Ксилулозо-5-фосфат
Транскетолаза (ТПФ)
Седогептулозо-7-фосфат 3-ФГА
Затем два образовавшиеся соединения реагируют друг с другом в трансальдолазной реакции; при этом в результате переноса 3С-фрагмента от седогептулозо-7-фосфата на 3-ФГА (3-фосфоглицериновый альдегид) образуются эритрозо-4-фосфат и фруктозо-6-фосфат.
С едогептулозо-7-фосфат
3-ФГА
едогептулозо-7-фосфат
3-ФГА
Трансальдолаза
Эритрозо-4-фосфат Фруктозо-6-фосфат
Однако реакция может идти и по другому пути (.6.17). В этом случае в трансальдолазной реакции образуется октулозо-1,8-дифосфат.
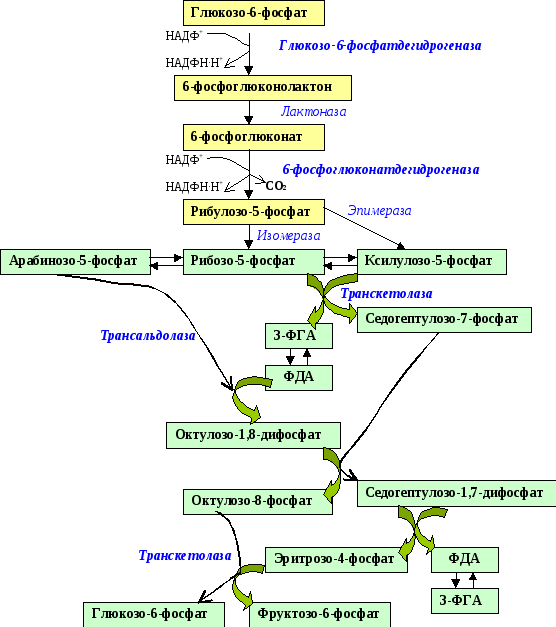
Рис.1. Пентозофосфатный (апотомический) путь обмена глюкозы
Эритрозо-4-фосфат и фруктозо-6-фосфат могут вступать в транскетолазную реакцию, в результате которой образуются фруктозо-6-фосфат и 3-ФГА:
Э ритрозо-4-фосфат
Фруктозо-6-фосфат
ритрозо-4-фосфат
Фруктозо-6-фосфат
Транскетолаза (ТПФ)
Фруктозо-6-фосфат 3-ФГА
Общее уравнение окислительной и неокислительной частей пентозофосфатного пути можно представить в следующем виде:
6 Глюкозо-6-ф-т + 7Н2О
+ 12
НАДФ+
5 Глюкозо-5-ф-т + 6СО2
+
12 НАДФН∙Н+
+
Рн
Глюкозо-6-ф-т + 7Н2О
+ 12
НАДФ+
5 Глюкозо-5-ф-т + 6СО2
+
12 НАДФН∙Н+
+
Рн
Значение пентозофосфатного пути окисления глюкозы
Ферменты пентозофосфатного пути локализуются в цитоплазме. В тканях, синтезирующих стероиды или жирные кислоты (для чего необходим НАДФН∙Н+), окислительная часть пути протекает весьма интенсивно. К таким тканям относятся: надпочечники, печень, жировая ткань, лактирующая молочная железа. Эритроциты нуждаются в коферменте НАДФН∙Н+ для восстановления глутатиона (трипептид). Совместно с витамином С восстановленный глутатион играет основную роль в предупреждении образования метгемоглобина: глутатион-SH является активной частью глутатионпероксидазы, устраняющей токсическое влияние пероксида водорода и других перекисей, окисляющих железо гемоглобина и нарушающих его кислородтранспортную функцию.

глутатион –S-S-глутатион
НАДФН∙ Н+ Н2О
Глутатионредуктаза Глутатионпероксидаза
НАДФ+ Н2О2