

биохимия (Восстановлен)
.pdfЧасть глюкозы (более половины) через капилляры кишечных ворсинок попадает в кровеносную систему и по воротной вене доставляется в печень. Остальное количество глюкозы поступает в клетки других тканей.
Б. Транспорт глюкозы из крови в клетки
Потребление глюкозы клетками из кровотока происходит также путѐм облегчѐнной диффузии. Следовательно, скорость трансмембранного потока глюкозы зависит только от градиента еѐ концентрации. Исключение составляют клетки мышц и жировой ткани, где облегчѐнная диффузия регулируется инсулином (гормон поджелудочной железы). В отсутствие инсулина плазматическая мембрана этих клеток непроницаема для глюкозы, так как она не содержит белки-переносчики (транспортѐры) глюкозы. Транспортѐры глюкозы называют также рецепторами глюкозы. Например, описан транспортѐр глюкозы, выделенный из эритроцитов. Это трансмембранный белок, полипептидная цепь которого построена из 492 аминокислотных остатков и имеет доменную структуру. Полярные домены белка расположены по разные стороны мембраны, гидрофобные располагаются в мембране, пересекая еѐ несколько раз. Транспортѐр имеет участок связывания глюкозы на внешней стороне мембраны. После присоединения глюкозы конформация белка изменяется, в результате чего глюкоза оказывается связанной с белком в участке, обращѐнном внутрь клетки. Затем глюкоза отделяется от транспортѐра, переходя внутрь клетки (см. раздел
5).
Считают, что способ облегчѐнной диффузии по сравнению с активным транспортом предотвращает транспорт ионов вместе с глюкозой, если она транспортируется по градиенту концентрации.
Глюкозные транспортѐры (ГЛЮТ) обнаружены во всех тканях. Существует несколько разновидностей ГЛЮТ (табл. 7-1), они пронумерованы в соответствии с порядком их обнаружения.
Структура белков семейства ГЛЮТ отличается от белков, транспортирующих глюкозу через мембрану в кишечнике и почках против градиента концентрации.
Описанные 5 типов ГЛЮТ имеют сходные первичную структуру и доменную организацию.
ГЛЮТ-1 обеспечивает стабильный поток глюкозы в мозг;
ГЛЮТ-2 обнаружен в клетках органов, выделяющих глюкозу в кровь. Именно при участии ГЛЮТ-2 глюкоза переходит в кровь из энтероцитов и печени. ГЛЮТ-2 участвует в транспорте глюкозы в β- клетки поджелудочной железы;
ГЛЮТ-3 обладает большим, чем ГЛЮТ-1, сродством к глюкозе. Он также обеспечивает постоянный приток глюкозы к клеткам нервной и других тканей;
ГЛЮТ-4 - главный переносчик глюкозы в клетки мышц и жировой ткани;
ГЛЮТ-5 встречается, главным образом, в клетках тонкого кишечника. Его функции известны недостаточно.
Все типы ГЛЮТ могут находиться как в плазматической мембране, так и в цитозольных везикулах. ГЛЮТ-4 (и в меньшей мере ГЛЮТ-1) почти полностью находятся в цитоплазме клеток. Влияние инсулина на такие клетки приводит к перемещению везикул, содержащих ГЛЮТ, к плазматической мембране, слиянию с ней и встраиванию транспортѐров в мембрану. После чего возможен облегчѐнный транспорт глюкозы в эти клетки. После снижения концентрации инсулина в крови транспортѐры глюкозы снова перемещаются в цитоплазму, и поступление глюкозы в клетку прекращается (рис. 7-19).
Перемещение глюкозы из первичной мочи в клетки почечных канальцев происходит вторичноактивным транспортом, подобно тому, как это осуществляется при всасывании глюкозы из просвета кишечника в энтероциты. Благодаря этому глюкоза может поступать в клетки даже в том случае, если еѐ концентрация в первичной моче меньше, чем в клетках. При этом глюкоза реабсорбируется из первичной мочи почти полностью (99%).
Известны различные нарушения в работе транспортѐров глюкозы. Наследственный дефект этих белков может лежать в основе инсулинонезависимого сахарного диабета (см. раздел 11). В то же время причиной нарушения работы транспортѐра глюкозы может быть не только дефект самого белка. Нарушения функции ГЛЮТ-4 возможны на следующих этапах:
передача сигнала инсулина о перемещении этого транспортѐра к мембране;
перемещение транспортѐра в цитоплазме;
включение в состав мембраны;
отшнуровывание от мембраны и т.д.
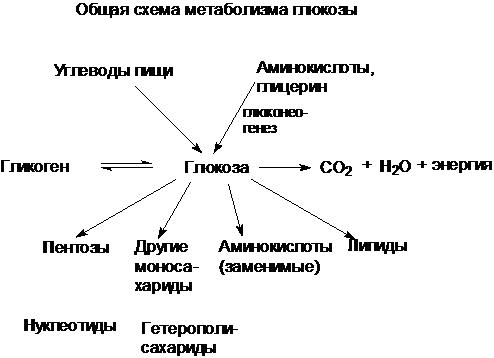
56. Глюкоза как важный метаболит углеводного обмена: общая схема источников и путей расходования глюкозы в организме. Поддерживание постоянного уровня глюкозы крови, количественное определение глюкозы крови.
В организме человека имеется несколько десятков различных моносахаридов и очень много (тысячи) разных олиго- и полисахаридов. Функции углеводов в организме заключаются в следующем.
Углеводы служат источником энергии: за счет их окисления удовлетворяется примерно половина всей потребности человека в энергии. В энергетическом обмене главная роль принадлежит глюкозе и гликогену.
Углеводы входят в состав структурно-функциональных компонентов клеток. К ним относятся пентозы нуклеотидов и нуклеиновых кислот, углеводы гликолипидов и гликопротеинов, гетерополисахариды межклеточного вещества.
Из углеводов в организме могут синтезироваться соединения других классов, в частности липиды и некоторые аминокислоты.
Таким образом, углеводы выполняют разнообразные функции, но главная функция углеводов - энергетическая.
Наиболее распространенный углевод животных - глюкоза. Именно в форме глюкозы поступает в кровь основная масса углеводов пищи. В глюкозу превращаются углеводы в печени и из глюкозы могут образовываться все остальные углеводы. Глюкоза используется как основной вид топлива в тканях млекопитающих. Таким образом, она играет роль связующего между энергетическими и пластическими функциями углеводов. Источником углеводов организма служат углеводы пищи - главным образом крахмал, а также сахароза и лактоза. Кроме того, глюкоза может образовываться в организме из аминокислот, а также из глицерина, входящего в состав жира.
В результате сложного взаимодействия описанных выше процессов создается тонко сбалансированное равновесие, обеспечивающее постоянный уровень глюкозы в циркулирующей крови. Если глюкоза не поступает с пищей, то у здорового человека в крови поддерживается постоянная концентрация глюкозы от 70 до 90 мг/100 мл. После приема содержащей углеводы пищи концентрация глюкозы на короткое время
возрастает примерно до 150 мг/ 100 мл, но через 2 ч она обычно возвращается к норме. Печень действует как первая буферная система; она удаляет поступающие по воротной вене моносахариды, образующиеся при переваривании пищи, и превращает их в глюкозу, а затем в гликоген. Поступление избытка глюкозы приводит к повышению ее концентрации в общем кровотоке, что в свою очередь стимулирует поглощение глюкозы периферическими тканями (мышцы, жировая клетчатка и другие). Достигая поджелудочной железы, избыточная концентрация глюкозы крови стимулирует выделение инсулина клетками островков Лангерганса; инсулин вызывает дальнейшее усиление поглощения глюкозы периферическими тканями. Как было отмечено выше, инсулин также стимулирует использование глюкозы для синтеза гликогена и подавляет образование глюкозы в ходе реакций глюконеогенеза; инсулин представляет собой главный действующий по принципу отрицательной обратной связи агент, участвующий в снижении содержания глюкозы в крови до нормальных величин после гипергликемии. Во время длительного голодания клетки почек также вносят значительный вклад в поддержание постоянного уровня глюкозы в крови. Другие ткани участвуют лишь в удалении глюкозы из внеклеточных жидкостей, поскольку в этих тканях отсутствует глюкозо-6-фосфатаза, необходимая для превращения других углеводных соединений в свободную глюкозу. Снижение концентрации глюкозы в крови, достигающей поджелудочной железы, стимулирует выделение островковыми клетками глюкагона. Этот гормон оказывает избирательное действие на печень, ускоряя гликогенолиз и способствуя повышению уровня глюкозы в крови. Таков кратковременный и быстро действующий ответ на лишение организма пищи, но он не обеспечит поддержания уровня глюкозы более чем на 24 ч; по истечении этого времени запасы гликогена в печени оказываются истощенными. Вторую линию обороны держит кора надпочечников, которая включается в эти процессы при длительном голодании, выделяя глюкокортикоиды. Эти гормоны стимулируют более медленный синтез ферментов глюконеогенеза в печени и усиливают распад белков в других тканях, что обеспечивает процесс глюконеогенеза углеродсодержащими субстратами. Поскольку этим эффектам в норме противодействует инсулин, реагирующий на гипергликемию. Очевидно, что именно сочетание и согласованность действия этих взаимосвязанных факторов, определяющих совершенство регуляции уровня глюкозы в крови, обеспечивает приспособление обмена углеводов в целом организме к изменяющимся условиям питания, роста, активности и другим физиологическим функциям.
Методы определение глюкозы в крови
Определение глюкозы в крови – важный этап, чтобы диагностировать сахарный диабет. Для этого необходимо определить состояние углеводного обмена, а в первую очередь содержание глюкозы в крови. В норме ее концентрация лежит в пределах 3, 3-5, 5 ммоль/л. Существует огромное количество методик определения, позволяющих установить глюкозы в крови.
Среди них можно выделить редуктометрические, колориметрические, ферментативные методы определения:
Редуктометрические методы определения. Основаны на способности сахаров, в частности глюкозы, восстанавливать соли тяжелых металлов в щелочной среде. Существуют различные реакции. Одна из них заключается в восстановлении красной кровяной соли в желтую кровяную соль сахарами при условии кипячения и щелочной среде. После этой специфической реакции путем титрования определяется содержание сахаров. Но этот метод не нашел широкого применения в клинике из-за своей трудоемкости и недостаточной точности;
Колорометрические методы определения. Глюкоза способна реагировать с различными соединениями, в результате чего образуются новые вещества определенной окраски (так называемые «цветные реакции»). По степени окраски раствора с помощью специального прибора (фотоколориметра) судят о концентрации глюкозы в крови. Примером такой реакции может послужить метод Самоджи. В основе его лежит реакция восстановление глюкозой гидрата окиси меди, в результате чего образуется молибденовая лазурь.
Ферментативные методы определения концентрации глюкозы в крови наиболее распространены. Выделяют две основные разновидности этих методов: глюкозооксидазный и гексокиназный. На данный момент наиболее распространенными являются глюкозооксидазные методы определения. Они основаны на применении фермента глюкозооксидазы. Этот фермент реагирует с глюкозой, в результате чего образуется перекись водорода. Количество образовавшейся перекиси водорода равно количеству глюкозы в исходной пробе.
Гексокиназный метод определения также является высокоспецифичным и точным, поэтому нашел широкое применение в клинической практике.
57. Аэробный гликолиз. Последовательность реакций до образования пирувата (аэробный гликолиз). Физиологическое значение аэробного гликолиза. Использование глюкозы для синтеза жиров.
Катаболизм глюкозы - основной поставщик энергии для процессов жизнедеятельности организма.
А. Основные пути катаболизма глюкозы
Окисление глюкозы до СО2 и Н2О (аэробный распад). Аэробный распад глюкозы можно выразить суммарным уравнением:
С6Н12О6 + 6 О2 → 6 СО2 + Н2О + 2820 кДж/моль.
Этот процесс включает несколько стадий (рис. 7-33).
Аэробный гликолиз - процесс окисления глюкозы с образованием двух молекул пирувата;
Общий путь катаболизма, включающий превращение пирувата в ацетил-КоА и его дальнейшее окисление в цитратом цикле;
ЦПЭ на кислород, сопряжѐнная с реакциями дегидрирования, происходящими в процессе распада глюкозы.
Б. Аэробный гликолиз
Аэробным гликолизом называют процесс окисления глюкозы до пировиноградной кислоты, протекающий в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки.
1. Этапы аэробного гликолиза
Ваэробном гликолизе можно выделить 2 этапа.
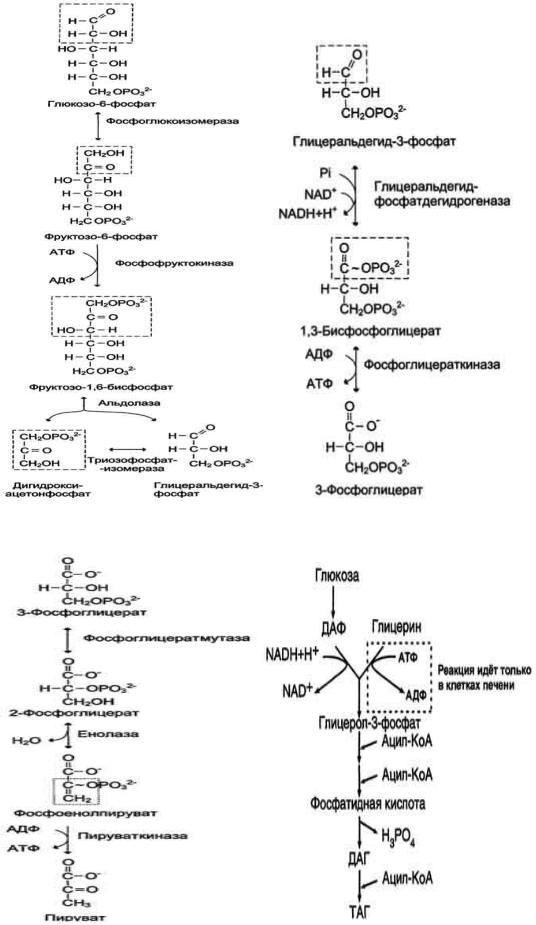
1.Подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта серия реакций протекает с использованием 2 молекул АТФ.
2.Этап, сопряжѐнный с синтезом АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 моль АТФ.
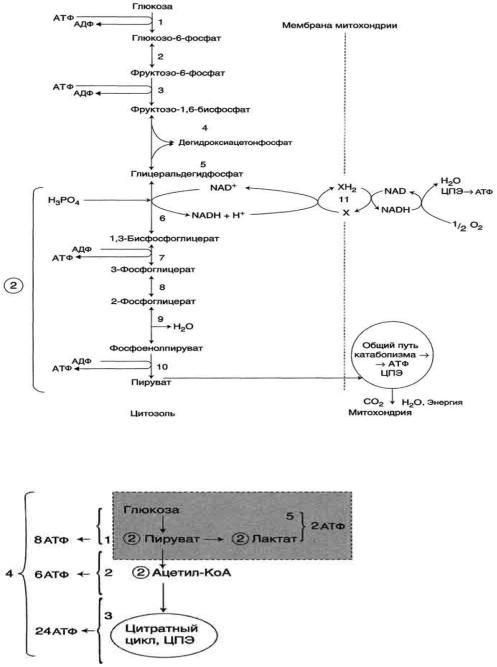
Аэробный распад глюкозы. 1-10- реакции аэробного гликолиза; 11 - малат-аспартатный челночный механизм транспорта водорода в митохондрии; 2 (в кружке) - стехиометрический коэффициент.
Пути катаболизма глюкозы. 1 - аэробный гликолиз; 2, 3 - общий путь катаболизма; 4 - аэробный распад глюкозы; 5 - анаэробный распад глюкозы (в рамке); 2 (в кружке) - стехиометрический коэффициент.
58. Анаэробный гликолиз. Реакция гликолитической оксидоредукции; субстратное фосфорилирование. Распространение и физиологическое значение анаэробного распада глюкозы.
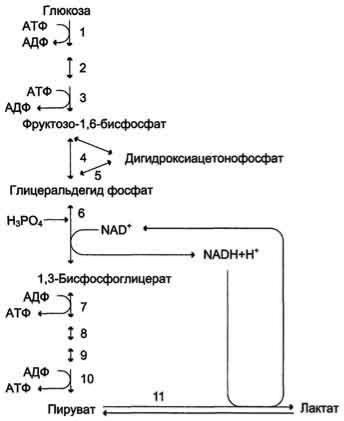
Анаэробным гликолизом называют процесс расщепления глюкозы с образованием в качестве конечного продукта лактата. Этот процесс протекает без использования кислорода и поэтому не зависит от работы митохондриальной дыхательной цепи. АТФ образуется за счѐт реакций субстратного фосфорилирования. Суммарное уравнение процесса:
С6Н1206 + 2 Н3Р04 + 2 АДФ = 2 С3Н6О3 + 2 АТФ + 2 Н2O.
1. Реакции анаэробного гликолиза
При анаэробном гликолизе в цитозоле протекают все 10 реакций, идентичных аэробному гликолизу. Лишь 11-я реакция, где происходит восстановление пирувата цитозольным NADH, является специфической для анаэробного гликолиза. Восстановление пирувата в лактат катализирует лактатдегидро-геназа (реакция обратимая, и фермент назван по обратной реакции). С помощью этой реакции обеспечивается регенерация NAD+ из NADH без участия митохондриальной дыхательной цепи в ситуациях, связанных с недостаточным снабжением клеток кислородом. Роль акцептора водорода от NADH (подобно кислороду в дыхательной цепи) выполняет пируват. Таким образом, значение реакции восстановления пирувата заключается не в образовании лактата, а в том, что данная цитозольная реакция обеспечивает регенерацию NAD+. К тому же лактат не является конечным продуктом метаболизма, удаляемым из организма. Это вещество выводится в кровь и утилизируется, превращаясь в печени в глюкозу, или при доступности кислорода превращается в пируват, который вступает в общий путь катаболизма, окисляясь до СО2 и Н2О. Строение лактатдегидрогеназы, механизм действия и значение определения активности этого фермента для диагностики заболеваний описывались ранее в разделе 2.
Баланс АТФ при анаэробном гликолизе
Анаэробный гликолиз по сравнению с аэробным менее эффективен. В этом процессе катаболизм 1 моль глюкозы без участия митохондриальной дыхательной цепи сопровождается синтезом 2 моль АТФ и 2 моль лактата. АТФ образуется за счѐт 2 реакций субстратного
Рис. 7-40. Анаэробный гликолиз.
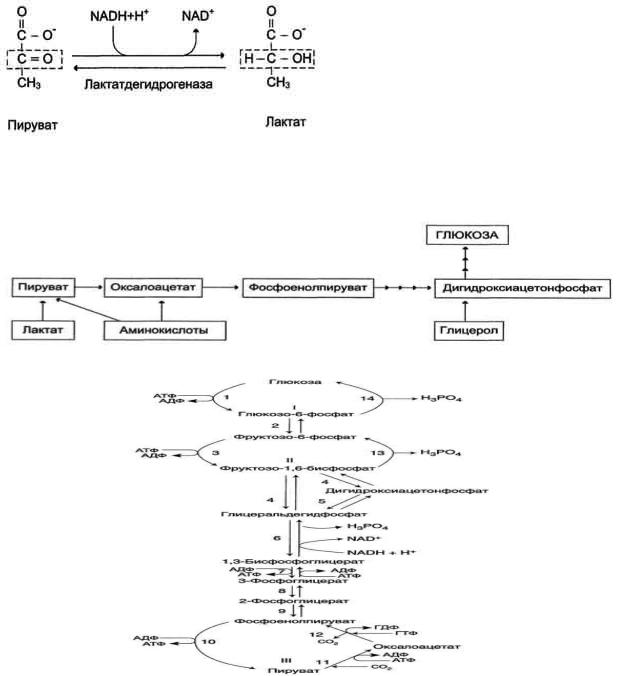
фосфорилирования. Поскольку глюкоза распадается на 2 фосфотриозы, то с учѐтом стехиометри-ческого коэффициента, равного 2, количество моль синтезированного АТФ равно 4. Учитывая 2 моль АТФ, использованных на первом этапе гликолиза, получаем конечный энергетический эффект процесса, равный 2 моль АТФ. Таким образом, 10 цитозольных ферментов, катализирующих превращение глюкозы в пируват, вместе с лактатдегидрогеназой обеспечивают в анаэробном гликолизе синтез 2 моль АТФ (на 1 моль глюкозы) без участия кислорода.
Анаэробный распад глюкозы происходит в мышцах, в первые минуты мышечной работы, в эритроцитах (в которых отсутствуют митохондрии), а также в разных органах в условиях ограниченного снабжении их кислородом, в том числе в клетках опухолей. Для метаболизма клеток опухолей характерно ускорение как аэробного, так и анаэробного гликолиза. Но преимущественный анаэробный гликолиз и увеличение синтеза лактата служит показателем повышенной скорости деления клеток при недостаточной обеспеченности их системой кровеносных сосудов.
59. Биосинтез глюкозы (глюконеогенез) из аминокислот, глицерина и молочной кислоты; регуляция глюконеогенеза. Биотин, роль в образовании оксалоацетата. Взаимосвязь гликолиза в мышцах и глюконеогенеза в печени (цикл Кори).
Рис. 7-44. Включение субстратов в глюконеогенез.
Рис. 7-45. Гликолиз и глюконеогенез. Ферменты обратимых реакций гликолиза и глюконеогенеза: 2 - фосфоглюкоизоме-раза; 4 - альдолаза; 5 - триозофосфатизомераза; 6 - глицеральдегидфосфатдегидрогеназа; 7 -фосфоглицераткиназа; 8 - фосфоглицератмутаза; 9 - енолаза. Ферменты необратимых реакций глюконеогенеза: 11 - пируваткарбоксилаза; 12 - фосфоенолпируваткарбоксикиназа; 13 - фруктозо-1,6-бисфосфатаза; 14 -глюкозо-6-фосфатаза. I-III -субстратные циклы.
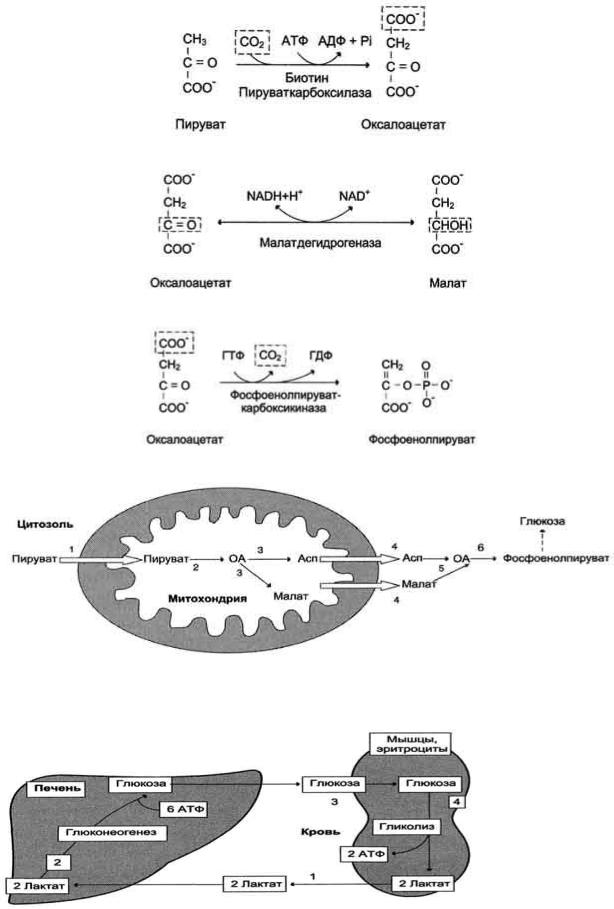
Рис. 7-46. Образование оксалоацетата из пирувата.
Рис. 7-47. Превращение оксалоацетата в малат.
Рис. 7-48. Превращение оксалоацетата в фосфоенолпируват.
Рис. 7-49. Образование оксалоацетата, транспорт в цитозоль и превращение в фосфоенолпируват. 1 - транспорт пирувата из цитозоля в митохондрию; 2 - превращение пирувата в оксалоацетат (ОА); 3 - превращение ОА в малат или аспартат; 4 - транспорт аспартата и малата из митохондрии в цитозоль; 5 - превращение аспартата и малата в ОА; 6 - превращение ОА в фосфоенолпируват.
Рис. 7-50. Цикл Кори (глюкозолактатный цикл). 1 - поступление лактата из сокращающейся мышцы с током крови в печень; 2 - синтез глюкозы из лактата в печени; 3 - поступление глюкозы из печени с током крови в работающую мышцу; 4 - использование глюкозы как энергетического субстрата сокращающейся мышцей и образование лактата.
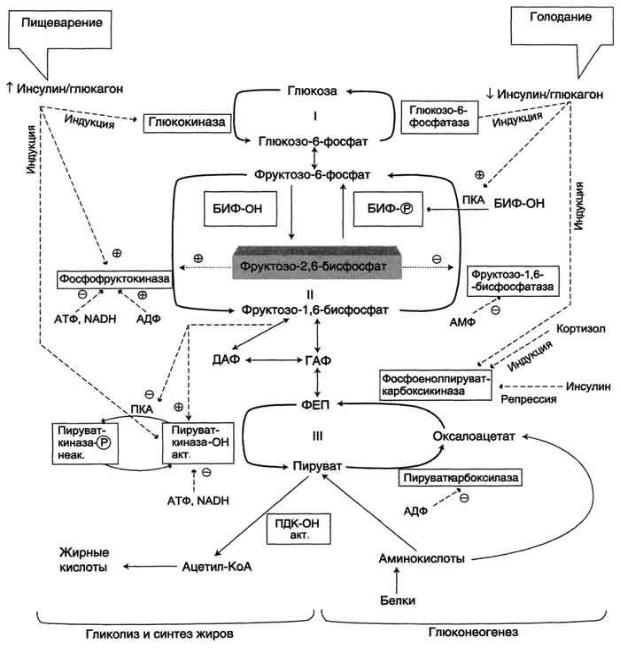
Рис. 7-54. Регуляция метаболизма глюкозы в печени. БИФ - бифункциональный фермент (фруктозо-2,6-
бисфосфатаза/фос-фофруктокиназа-2); БИФ-ОН - дефосфорилированный фермент; БИФ-Р - фосфорилированный фермент, ПДК-ОН - дефосфорилированный пируватдегидрогеназный комплекс; ПКОН - дефосфорилированная пируваткиназа; ГАФ - глицеральдегидфосфат; ДАФ - дигидроксиацетонфосфат, ФЕП - фосфоенолпируват. I-III - субстратные циклы: в рамках - регупяторные ферменты глиполиза и глюконеогенеза.
59. Гликоген, биологическое значение. Биосинтез и мобилизация гликогена. Регуляция синтеза и распада гликогена.
Многие ткани синтезируют в качестве резервной формы глюкозы гликоген. Синтез и распад гликогена обеспечивают постоянство концентрации глюкозы в крови и создают депо для еѐ использования тканями по мере необходимости.
Гликоген - разветвлѐнный гомополимер глюкозы, в котором остатки глюкозы соединены в линейных участках α-1,4-гликозидной связью. В точках ветвления мономеры соединены α-1,6-гликозидными связями. Эти связи образуются примерно с каждым десятым остатком глюкозы. Следовательно, точки ветвления в гликогене встречаются примерно через каждые десять остатков глюкозы. Так возникает древообразная