

биохимия (Восстановлен)
.pdfсродство к гормону, может изменяться также в результате ковалентной модификации, главным образом путѐм фосфорилирования. Концентрация внутриклеточных рецепторов может также регулироваться по механизму индукции и репрессии.
Б. Механизмы передачи гормональных сигналов в клетки
По механизму действия гормоны можно разделить на 2 группы. К первой группе относят гормоны, взаимодействующие с мембранными рецепторами (пептидные гормоны, адреналин, а также гормоны местного действия - цитокины, эйкозаноиды). Вторая группа включает гормоны, взаимодействующие с внутриклеточными рецепторами.
Связывание гормона (первичного посредника) с рецептором приводит к изменению кон-формации рецептора. Это изменение улавливается другими макромолекулами, т.е. связывание гормона с рецептором приводит к сопряжению одних молекул с другими (трансдукция сигнала). Таким образом, генерируется сигнал, который регулирует клеточный ответ путѐм изменения активности или количества ферментов и других белков. В зависимости от способа передачи гормонального сигнала в клетках меняется скорость реакций метаболизма:
в результате изменения активности ферментов;
в результате изменения количества ферментов .
82.Циклические АМФ и ГМФ как вторичные посредники. Активация протеинкиназ и фосфорилирование белков, ответственных за проявление гормонального эффекта.
Передача гормональных сигналов через мембранные рецепторы
Гормоны (первичные посредники), связываясь с рецепторами на поверхности клеточной мембраны, образуют комплекс гормон-рецептор, который трансформирует сигнал первичного посредника в изменение концентрации особых молекул внутри клетки - вторичных посредников. Вторичными посредниками могут быть следующие молекулы: цАМФ, цГМФ, ИФ3, ДАТ, Са2+, NO.
Гормоны, взаимодействие которых с рецептором клетки-мишени приводит к образованию цАМФ, действуют через трѐхкомпонентную систему, которая включает белок-рецептор, G-белок и фермент аденилатциклазу. Образующийся под действием аденилатциклазы цАМФ активирует протеинкиназу А, фосфорилируюшую ферменты и другие белки (см. раздел 5). Известно более 200 различных G-белков, в структуре которых обнаружены 3 субъединицы α, β и γ (см. раздел 5). В отсутствие гормона α-субъединица G-белка связана с ГДФ. Образование комплекса гормонрецептора приводит к конформационным изменениям α-субъединицы, замене ГДФ на ГТФ и отщеплению димера βγ от α-ГТФ. В случае рецепторов, сопряжѐнных с Gs-белком, субъединица αs-ГТФ активирует аденилатциклазу
В случае рецепторов, сопряжѐнных с Giбелком, субъединица αi-ГТФ ингибирует аденилатциклазу. В таблице 11-4 приведены примеры гормонов, взаимодействие которых с соответствующим рецептором активирует или ингибирует аденилатциклазу.
Другая система, генерирующая цГМФ как вторичный посредник, сопряжена с гуанилатциклазой. Цитоплазматический домен такого типа рецепторов обладает активностью гуанилатциклазы, которая катализирует реакцию образования цГМФ из ГТФ> (подобно аденилатциклазе). Молекулы цГМФ могут активировать ионные каналы либо активировать цГМФ-за-висимую протеинкиназу G, участвующую в фосфорилировании других белков в клетке. Например, фосфодиэстерааа, которая гидролизует цАМФ до АМФ, активируется в результате фосфорилирования цГМФ-зависимой протеинкиназой.
83. Фосфатидилинозитольный цикл как механизм внутриклеточной коммуникации. Инозитол 1,4,5-трифосфат и диацилглицерол - вторичные посредники передачи сигнала. Ионы кальция как вторичные посредники, кальмодуллин.
Некоторые гормоны (например, вазопрессин или адреналин), образуя комплекс с соответствующими рецепторами (рецептор V1 для вазопрессина и αi-рецептор для адреналина), через активацию соответствующих G-белков активируют фосфолипазу С, в результате чего в клетке появляются вторичные посредники ИФ3, ДАГ. Молекула ИФ3 стимулирует высвобождение Са2+ из ЭР. Кальций связывается с
белком кальмодулином. Этот комплекс активирует Са2+-кальмодулинзависимую протеинкиназу. Ионы кальция и ДАГ участвуют в активации протеинкиназы С
84. Передача сигналов через внутриклеточные рецепторы. Образование комплекса гормонрецептор и его взаимодействие с ДНК, гормон чувствительные элементы (HRE). Передача сигналов через рецепторы сопряженные с ионными каналами. Строение рецептора ацетилхолина.
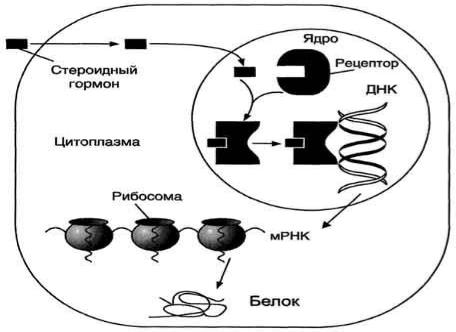
Стероидные и тиреоидные гормоны связываются с рецепторами внутри клетки и регулируют скорость транскрипции специфических генов .
В отсутствие гормона внутриклеточные рецепторы связаны обычно с другими белками в цитозоле или ядре. Например, рецепторы глюкокортикоидов образуют в цитозоле комплекс с шапероном, что препятствует связыванию рецептора с молекулой ДНК .
Взаимодействие гормона с центром связывания на С-концевом участке полипептидной цепи рецептора вызывает конформационные изменения и освобождение рецептора от шаперона. Происходит объединение 2 молекул рецептора с образованием гомодимера. Димер рецептора узнаѐт специфическую последовательность нуклеотидов, которая расположена в промоторной области гена. Взаимодействие со специфическим участком ДНК HRE (от англ, hormone response element, элемент, реагирующий на воздействие гормона) обеспечивает центральный домен рецептора. Этот домен содержит аминокислотную последовательность, образующую 2 "цинковых пальца". В каждом "цинковом пальце" атом цинка связан с 4 остатками цистеина (рис. 11-7).
В структуре одного "цинкового пальца" имеется последовательность аминокислот, отвечающая за связывание с ДНК, а второй "цинковый
Рис. 11-5. Передача гормональных сигналов через внутриклеточные рецепторы (рецепторы стероидных гормонов могут находиться в цитоплазме и ядре).
палец" содержит последовательность аминокислот, участвующую в димеризации рецепторов. Взаимодействие комплекса гормон-рецептор с определѐнной последовательностью нуклеотидрв в промоторной части ДНК приводит к активации транскрипции.
Рецепторы тиреоидных гормонов всегда связаны с ДНК. В отсутствие гормонов соответствующие рецепторы ингибируют экспрессию генов. Напротив, взаимодействие с гормоном превращает их в активаторы транскрипции.
3. Передача сигналов через рецепторы, сопряжѐнные с ионными каналами
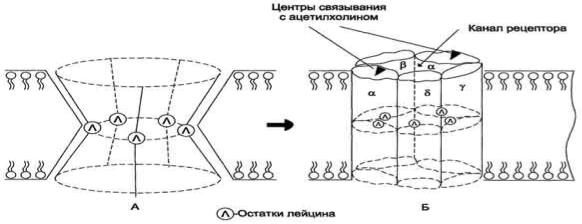
Рецепторы, сопряжѐнные с ионными каналами, являются интегральными мембранными белками, состоящими из нескольких субъединиц. Они действуют одновременно как ионные каналы и как рецепторы, которые способны специфически связывать с внешней стороны эффектор, изменяющий их ионную проводимость. Эффекторами такого типа могут быть гормоны и нейромедиаторы .
Известны рецепторы для ряда гормонов, ассоциированных с ионными каналами, и большинства медиаторов, среди которых наиболее изучен рецептор ацетилхолина. Рецептор ацетилхолина состоит из пяти цилиндрообразных субъединиц, расположенных в мембране параллельно друг другу:α2, β, γ, δ. Между ними вдоль оси цилиндров находится заполненный молекулами воды канал. Каждая субъединица рецептора состоит из большого количества гидрофобных аминокислотных остатков. Кроме этого, все субъединицы содержат один спирализованный трансмембранный фрагмент, аминокислотные радикалы которого (полярные незаряженные аминокислотные остатки, в основном серии и треонин) выстилают центральный канал рецептора изнутри. В средней части субъединиц, обращѐнной к каналу, локализованы остатки лейцина. В присутствии ацетилхолина боковые взаимодействия между субъединицами поддерживают канал в открытом состоянии и создают возможность для транспорта ионов. В отсутствие ацетилхолина в результате изменения ориентации субъединиц относительно друг друга канал закрывается, так как выступающие внутрь канала остатки лейцина образуют плотное гидрофобное кольцо, блокируя движение гидратированных ионов в этой области .
Схема строения рецептора ацетилхолина. А - закрытый канал рецептора в отсутствие ацетилхолина; Б - открытый канал рецептора в присутствии ацетилхолина. Трансмембранные спирализованные участки всех 5 субъединиц содержат полярные незаряженные радикалы аминокислот; гидрофобные остатки лейцина (Л), локализованные в середине каждого спирализованного гидрофильного участка, выступают в центральную часть канала и препятствуют движению ионов.
85. Гормоны гипоталамуса и передней доли гипофиза, химическая природа и биологическая роль.
Гормоны гиппоталамуса
Гипоталамус занимает важнейшее место в иерархической системе, объединяя высшие отделы ЦНС и эндокринные железы. В клетках нейронов гипоталамуса синтезируются пептидные гормоны 2 типов. Одни через систему гипоталамо-гипофизарных сосудов поступают в переднюю долю гипофиза, где стимулируют или ингибируют синтез тропных гормонов; другие, как окситоцин и вазопрессин, поступают через аксоны нервных клеток в заднюю долю гипофиза, где они хранятся в везикулах и секрети-руются в кровь в ответ на соответствующие сигналы.
В настоящее время известно несколько гипоталамических гормонов, регулирующих синтез и секрецию гормонов гипофиза .
1. Тиреолиберин - трипептид, состоящий из пироглутаминовой кислоты, гистидина и пролинамида . Синтез тиреолиберина происходит в различных участках гипоталамуса, но в большей степени в паравентрикулярном ядре, а также в других областях ЦНС, где он выполняет функцию нейромедиатора, повышающего двигательную активность и АД. Предшественник тиреолиберина препротиреолиберин человека включает 242 аминокислотных остатка. Образование активного гормона происходит по механизму частичного протеолиза. В передней доле гипофиза тиреолиберин стимулирует синтез и секрецию
тиреотропина, а также оказывает стимулирующее влияние на синтез многих других гормонов. В результате взаимодействия тиреолиберина с рецепторами плазматической мембраны клеток гипофиза происходит повышение концентрации внутриклеточного цАМФ и Са2+. Трансдукция сигнала происходит как через аденилатциклазную, так и через инозитолфосфатную системы.
Тиреолиберин разрушается в клетках-мишенях и в крови под действием специфических протеаз. Т1/2 в крови составляет 3-4 мин.
2. Кортиколиберин
Кортиколиберин - полипептид, содержащий 41 аминокислотный остаток. Как и другие пептидные гормоны, Кортиколиберин синтезируется в виде прогормона. Т1/2 кортиколиберина в плазме крови составляет 60 мин. Основное количество кортиколиберина образуется в гипоталамусе, однако он обнаруживается и в других отделах ЦНС, где выполняет роль медиатора, участвуя в ответной реакция на различные стрессовые ситуации.
В передней доле гипофиза кортиколиберин увеличивает синтез и секрецию проопиомеланокортина и образование кортикотропина. Рецепторы кортиколиберина находятся в плазматической мембране клеток в составе аденилатциклазного комплекса. Стимуляция секреции АКТГ требует присутствия ионов Са2+. Увеличение уровня внутриклеточного кальция, вероятно, является результатом фосфорилирования белков кальциевых каналов.
3. Гонадолиберин
Гонадолиберин - декапептид. Предшественник гонадолиберина человека состоит из 92 аминокислотных остатков и имеет молекулярную массу около 10 кД. Гонадолиберин стимулирует синтез и секрецию 2 гормонов гипофиза - ЛГ и ФСГ. Помимо гипоталамуса, нейроны, содержащие Гонадолиберин, находятся и в других областях ЦНС, контролирующих эмоциональное и половое поведение. Рецептор гонадолиберина в плазматической мембране входит в состав инозитолфосфатного комплекса, активация которого стимулирует фосфорилирование белков и мобилизацию Са2+, что приводит к освобождению гормонов. Т 1/2 гонадолиберина в плазме крови составляет 5-7 мин. Инактивация гонадолиберина происходит при участии специфических протеаз.
4. Соматолиберин
Соматолиберин - полипептид, состоящий из 44 аминокислотных остатков. В передней доле гипофиза Соматолиберин стимулирует синтез и секрецию соматотропина. Трансдукция сигнала сопровождается повышением концентрации как цАМФ, так и ионов кальция. Т1/2 соматолиберина в крови составляет около 7 мин. Соматолиберин применяют в клинической практике для диагностики нарушений функции гипофиза.
5.Соматостатин
Соматостатин первично был выделен из гипоталприсутствуют во многих опухолевых клетках, секретирующих гормоны. Это обстоятельство используется для разработки методов ранней диагностики опухолей поджелудочной железы, феохромоцитомы, рака щитовидной железы, рака почек и молочной железы.
Результат трансдукции сигнала соматостатина - снижение уровня внутриклеточной концентрации цАМФ и Са2+ в цитозоле клеток. Соматостатин тормозит секрецию гормона роста, глюкагона, инсулина, гастрина, секретина, вазоактивного интестинального пептида (ВИП, VIP), холецистокинина, кальцитонина, паратгормона, иммуноглобулинов, ренина; он также ингибирует секрецию бикарбонатов и ферментов поджелудочной железы, уменьшает кровоток на всѐм протяжении ЖКТ, снижает секрецию жѐлчи.
Б. Гормоны гипофиза
Гипофиз секретирует большое количество гормонов, участвующих в регуляции различных биохимических процессов и физиологических функций. В передней доле гипофиза (аденогипофизе) синтезируются так называемые тропные гормоны, стимулирующие синтез и секрецию гормонов других эндокринных желѐз или оказывающие влияние на метаболические реакции в других тканях-мишенях
Задняя доля гипофиза, или нейрогипофиз, секретирует гормоны, регулирующие в основном водный баланс и лактацию.
Секреция гормонов гипофиза обусловлена сочетанием нервных и гуморальных сигналов. При этом один и тот же агонист (например, норадреналин) может вызывать противоположные изменения в секреции гипофизарных гормонов. С другой стороны, секреция каждого гормона может контролироваться многочисленными факторами.
Синтез и секреция гормонов передней доли гипофиза регулируются гормонами гипоталамуса, которые поступают в гипофиз через портальную систему кровеносных сосудов, связывающих гипоталамус и переднюю долю гипофиза. Кроме того, секреция гормонов гипоталамуса и гипофиза регулируется по механизму обратной связи гормонами, продукцию которых они стимулируют в органах-мишенях.
В передней доле гипофиза синтезируются гормоны, которые по химическому строению являются пептидами и гликопротеинами.
По механизму их синтеза и биологическим функциям эти гормоны объединяют в 3 группы.
1. Гормон роста, пролактин
Гормон роста синтезируется в соматотроф-ных клетках, наиболее многочисленных в передней доле гипофиза. Содержание гормона роста составляет 5-16 мг в 1 г ткани железы, в то время как количество других гормонов гипофиза исчисляется в мкг/г. Т1/2 гормона в плазме крови составляет около 50 мин.
Гормон роста у всех видов млекопитающих представляет собой одноцепочечный пептид с молекулярной массой 22 кД, состоящий из 191 аминокислотного остатка и имеющий 2 внутримолекулярные дисульфидные связи .
Гормон роста образуется из прогормона с молекулярной массой 28 кД, не обладающего гормональной активностью. Уровень гормона роста в плазме крови не превышает 3 нг/мл. Секреция гормона роста носит пульсирующий характер с интервалами в 20-30 мин. Один из самых больших пиков отмечается вскоре после засыпания.
Под влиянием различных стимулов (стресс, физические упражнения, гипогликемия, голодание, белковая пища, аминокислота аргинин) даже у нерастущих взрослых людей уровень гормона роста в крови может возрастать до 30-100 нг/мл.
Регуляция синтеза и секреции гормона роста осуществляется множеством факторов. Основной стимулирующий эффект оказывает соматолиберин, основной тормозящий - гипоталамический соматостатин.
Рецепторы гормона роста находятся в плазматической мембране клеток печени, жировой ткани, яичках, жѐлтом теле, скелетных мышцах, хрящевой ткани, мозге, лѐгких, поджелудочной железе, кишечнике, сердце, почках, лимфоцитах. Рецептор гормона роста - белок с одним внутримембранным доменом и молекулярной массой 70 кД. Связывание рецептора с гормоном роста вызывает димеризацию 2 рецепторов, что приводит к активации связанных с рецептором Янус-киназ и фосфорилированию Янускиназ и рецептора по остаткам тирозина. Активация рецептора гормона роста сопровождается повышением активности тирозинкиназ и фосфолипазы С с последующим повышением уровня ДАГ и ИФ3 и активацией протеинкиназы С .
Первичные эффекты гормона роста кратковременны и инсулиноподобны. Они проявляются в основном в отношении обмена жиров и углеводов. В жировой ткани усиливается потребление глюкозы и липогенез, вследствие чего происходит снижение концентрации глюкозы в крови. Однако в дальнейшем проявляются более медленные (в основном, противоположные инсулину) эффекты: усиливается липолиз в жировой ткани, увеличивается концентрация жирных кислот в крови, а в случае недостаточности инсулина увеличивается содержание кетоновых тел в крови. Энергия, образующаяся при повышенном распаде жиров, используется на анаболические процессы. В то же время использование глюкозы жировыми и мышечными клетками снижается, а в печени ускоряется глюконеогенез, следствием чего может быть гипергликемия, особенно при недостатке инсулина . Основное действие гормона роста направлено на регуляцию обмена белков и процессов, связанных с ростом и развитием организма. Под влиянием гормона роста усиливаются
транспорт аминокислот в клетки мышц, синтез белка в костях, хрящах, мышцах, печени и других внутренних органах, увеличивается общее количество РНК, ДНК и общее число клеток.
Влияние гормона роста на рост скелета и мягких тканей требует участия веществ, которые синтезируются в ответ на взаимодействие гормона роста с рецепторами плазматической мембраны клеток различных тканей, в основном печени, и носят название соматомединов. Поскольку эти молекулы отличаются высокой гомологичностью друг к другу, а также к проинсулину и обладают инсулиноподобной активностью и мощным ростстимулирующим действием, они называются инсулиноподобными факторами роста (ИФР-1, или соматомедин С; ИФР-2, или соматомедин А). ИФР-1 - одно-цепочечный полипептид основного характера, содержащий 70 аминокислотных остатков, а полипептид ИФР-2 носит кислотный характер и состоит из 67 аминокислотных остатков. В крови примерно 95% соматомединов циркулирует в комплексе с белками. Синтез ИФР-1 в большей степени зависит от концентрации гормона роста в крови, чем синтез ИФР-2. В то же время ИФР-1, образующийся в печени, ингибирует синтез и секрецию гормона роста по механизму ретроингибирования, действуя на уровне гипофиза и гипоталамуса (рис. 11-12).
Инсулиноподобные факторы роста оказывают своѐ действие различными путями: эндокринным, паракринным и аутокринным .
Подобно рецептору инсулина, рецептор ИФР-1 обладает тирозинкиназной активностью и инициирует каскад реакций фосфорилирования других белков, участвующих в различных внутриклеточных процессах, включая активацию транскрипции генов. В большинстве случаев ИФР-1, как и инсулин, инициирует клеточное развитие, однако при значительно меньших, почти физиологических концентрациях. Это указывает на то, что инсулиноподобные факторы роста более активны в отношении их действия на рост и развитие клеток.
Под влиянием гормона роста увеличивается ширина и толщина костей, и одновременно с этим ускоряется рост других тканей, включая соединительную ткань, мыпщы и внутренние органы.
Пролактин синтезируется лактотрофными клетками передней доли гипофиза в виде прогормона с молекулярной массой 40 кД. Число этих клеток резко возрастает при беременности под влиянием эстрогенов. Пролактин близок по химическому строению гормону роста. Он состоит из 199 аминокислотных остатков, образующих одну полипептидную цепь с тремя дисульфидными связями. 35% аминокислотной последовательности пролактина идентично последовательностям гормона роста. Оба гормона имеют общие антигенные детерминанты, сходное строение рецепторов и пути трансдукции сигналов в клетки.
Рецепторы пролактина присутствуют в клетках многих тканей: в печени, почках, надпочечниках, яичках, яичниках, матке и других тканях.
Основная физиологическая функция пролактина - стимуляция лактации. Пролактин индуцирует синтез α-лактальбумина и казеина, активирует синтез фосфолипидов и ТАГ.
На процессы роста Пролактин влияет в значительно меньшей степени, чем гормон роста.
У мужчин Пролактин повышает чувствительность клеток Лейдига к лютеинизирующему гормону, поддерживая таким образом необходимый уровень синтеза тестостерона; в почках пролактин снижает экскрецию воды, влияет на реабсорбцию ионов Na+ и К+; Пролактин также повышает гуморальный и клеточный иммунитет.
Синтез и секрецию пролактина стимулируют тиреолиберин, серотонин, окситоцин, ацетил-холин, ингибирующий эффект оказывает дофамин.
Подобно большинству гормонов, Пролактин секретируется в кровь эпизодически с интервалами 30-90 мин. Максимум секреции отмечается через 6-8 ч после начала сна. Концентрация пролактина в плазме крови женщин составляет 8-10 нг/мл, а мужчин - 5-8 нг/мл. Т1/2 пролактина составляет 15-20 мин.
Плацента продуцирует гормон (плацентарный лактоген) , гомологичный по аминокислотному составу гормону роста и пролактину. Все 3 гормона имеют общие антигенные детерминанты и обладают ростстимулирующей и лакто-генной активностью. Существует гипотеза, согласно которой гены этих гормонов возникли в результате дупликации одного гена-предшественника.
2. Тиреотропин, лютеинизирующий гормон и фолликулостимулирующий гормон
Тиреотропин, ЛГ и ФСГ - гликопротеины. Тиреотропин (TIT) с молекулярной массой около 30 кД синтезируется в тиреотрофных клетках передней доли гипофиза.
Стимуляция секреции тиреотропина происходит под влиянием тиреолиберина, а основное ингибирующее действие оказывает повышение уровня тиреоидных гормонов. Пик секреции ТТГ отмечается в часы, непосредственно предшествующие сну, с последующим снижением в течение ночи.
Основная биологическая функция тиреотропина - стимуляция синтеза и секреции йод-тиронинов (Т3 и Т4) в щитовидной железе. Трансдукция сигнала тиреотропина в клетки щитовидной железы происходит через рецепторы плазматической мембраны и активацию аденилатциклазы.
Рецептор тиреотропина состоит из 2 доменов, один из которых представляет собой гликопротеин, а второй - ганглиозид (гликолипид, содержащий сиаловую кислоту). Для проявления биологического действия необходимо связывание тиреотропина с обоими доменами рецептора.
Тиреотропин оказывает на щитовидную железу 2 типа эффектов. Одни проявляются быстро (в течение нескольких минут) и включают стимуляцию всех стадий синтеза и секреции йодтиронинов (см. ниже подраздел III, В). Проявление других требует нескольких дней. К ним относят стимуляцию синтеза белков, фосфолипидов, нуклеиновых кислот, увеличение размеров и количества тиреоидных клеток.
Некоторые иммуноглобулины класса G, взаимодействуя с рецепторами тиреотропина, имитируют эффекты гормона. Подобные иммуноглобулины обнаруживаются у большинства больных гипертиреозом . Помимо стимулирующих, обнаруживаются и антитела, вызывающие разрушение клеток щитовидной железы. Образование антител, имитирующих эффекты тиреотропина, - одна из частых причин нарушений функций щитовидной железы.
В группу гормонов, относящихся к гликопротеинам, входят также гонадотропные гормоны гипофиза ЛГ и ФСГ и хорионический гонадотропин (ХГ) .
3. Группа гормонов, образующихся из проопиомеланокортина
Проопиомеланокортин (ПОМК) с молекулярной массой 28,5 кД синтезируется в передней и промежуточной долях гипофиза и в некоторых других тканях (кишечнике, плаценте). Полипептидная цепь ПОМК состоит из 265 аминокислотных остатков .
После отщепления сигнального пептида происходит частичный протеолиз оставшейся полипептидной цепи с образованием АКТГ и β-липотропина (β-ЛП). В разных клетках в результате избирательного протеолиза образуется разный набор пептидов: α- и β-меланоцитстимулирующих гормонов (α- и β-МСГ) и эндорфинов. β-МСГ и кортикотропиноподобный гормон промежуточной доли у человека практически не образуются, так как у взрослых людей промежуточная доля не развита. В гипофизе человека найдены β- липотропин, γ-липотропин и β-эндорфин. Функции всех продуктов разрушения ПОМК недостаточно изучены.
Кортикотропин (АКТГ) - пептидный гормон; состоит из 39 аминокислотных остатков; синтезируется в клетках передней доли гипофиза под влиянием кортиколиберина.
Кортикотропин секретируется в импульсивном режиме. Скорость секреции составляет 5-25 мкг/сут. При стрессе (травма, ожог, хирургическое вмешательство, интоксикация химическими веществами, кровотечение, боль, психическая травма) концентрация АКТГ в крови возрастает во много раз. У здоровых людей наименьший уровень АКТГ в крови отмечается в конце дня и непосредственно перед сном, наибольший - в 6-8 ч утра, в момент пробуждения. Т1/2 в крови составляет 15-25 мин.
Механизм действия АКТГ включает взаимодействие с рецептором плазматической мембраны клеток, активацию аденилатциклазы и фосфорилирование белков, участвующих в синтезе кортикостероидов (см. ниже подраздел III, Д). Эти эффекты усиливаются в присутствии ионов Са2+. В клетках коры надпочечников АКТГ стимулирует гидролиз эфиров холестерола, увеличивает поступление в клетки холестерола в составе
ЛПНП; стимулирует превращение холестерола в прегненолон; индуцирует синтез митохондриаль-ных и микросомальных ферментов, участвующих в синтезе кортикостероидов. Подробнее этапы синтеза кортикостероидов рассматриваются в подразделе III, Д.
4. Гормоны задней доли гипофиза
Задняя доля гипофиза, или нейрогипофиз, сек-ретирует 2 активных гормона - вазопрессин, или антидиуретический гормон (АДГ), и окситоцин.
Окситоцин и вазопрессин - ноншепгады со сходной первичной структурой (рис. 11-16).
Оба гормона образуются в гипоталамусе в нейронах разных гипоталамических ядер в форме прогормонов, из которых в результате посттрансляционной модификации образуются гормон и транспортный пептид нейрофизин (окситоцин+нейрофизин I и вазопрессин+нейрофизин II). В процессе транспорта в клетки задней доли гипофиза гормоны остаются нековалентно связанными со своими транспортными пептидами. В крови гормоны не связаны с нейрофизином. Т1/2 составляет 2-4 мин.
Основные биологические эффекты вазопрессина проявляются через взаимодействие с 2 типами рецепторов. V1-рецепторы расположены в клетках гладкой мускулатуры сосудов в комплексе с фосфолипазой С. Результат трансдукции сигнала в эти клетки - сокращение сосудов. V2-рецепторы расположены в клетках почечных канальцев. Взаимодействие вазопрессина с V2-рецепторами активирует аденилатциклазную систему, увеличивая в клетках концентрацию цАМФ и активность протеинкиназы А. В результате этой активации происходит фосфорилирование белков, стимулирующих экспрессию генов белков, которые образуют каналы, обеспечивающие реабсорбцию воды (см. ниже подраздел VI, А).
Окситоцин стимулирует сокращение гладкой мускулатуры матки, а также играет важную роль в стимуляции лактации. Он вызывает сокращение миоэпителиальных клеток молочных желѐз, в результате чего происходит перераспределение молока из альвеолярных протоков в область соска.
Акт сосания материнской груди стимулирует секрецию пролактина, обеспечивая образование и секрецию молока.
86. Регуляция водно-солевого обмена. Строение, механизмдействия и функции альдостерона и вазопрессина. Роль системы ренин-ангиотензин-альдостерон. Предсердный натриуретический фактор.
Важнейшие параметры вводно-солевого гомеостаза - осмотическое давление, рН и объѐм внутриклеточной и внеклеточной жидкости. Изменение этих параметров может привести к изменению
АД, ацидозу или алкалозу, дегидратации и отѐкам тканей. Основные гормоны, участвующие в тонкой регуляции водно-солевого баланса и действующие на дистальные извитые канальцы и собирательные трубочки почек: антидиуретический гормон (АДГ), альдостерон и предсердный натриуретический фактор
(ПНФ).
А. Антидиуретический гормон
Антидиуретический гормон (АДГ), или вазопрессин - пептид с молекулярной массой около 1100 Д, содержащий 9 аминокислот, соединѐнных одним дисульфидным мостиком.
1. Синтез и секреция антидиуретического гормона
АДГ синтезируется в нейронах гипоталамуса в виде предшественника препрогормона, который поступает в аппарат Гольджи и превращается в прогормон. В составе нейросекреторных гранул прогормон переносится в нервные окончания задней доли гипофиза (нейрогипофиз). Во время транспорта гранул происходит процессинг прогормона, в результате чего он расщепляется на зрелый гормон и транспортный белок - нейрофизин. Гранулы, содержащие зрелый антидиуретический гормон и нейрофизин, хранятся в терминальных расширениях аксонов в задней доле гипофиза, из которых секретируются в кровоток при соответствующей стимуляции.
Стимулом, вызывающим секрецию АДГ, служит повышение концентрации ионов натрия и увеличение осмотического давления внеклеточной жидкости. При недостаточном потреблении воды, сильном потоотделении или после приѐма большого количества соли осморецепторы гипоталамуса, чувствительные к колебаниям осмолярности, регистрируют повышение осмотического давления крови. Возникают нервные импульсы, которые передаются в заднюю долю гипофиза и вызывают высвобождение АДГ. Секреция АДГ происходит также в ответ на сигналы от барорецепторов предсердий. Изменение осмолярности всего на 1% приводит к заметным изменениям секреции АДГ.
2. Механизм действия
Для АДГ существуют 2 типа рецепторов: V1 и V2. Рецепторы V2, опосредующие главный физиологический эффект гормона, обнаружены на базолатеральной мембране клеток собирательных трубочек и дистальных канальцев - наиболее важных клеток-мишеней для АДГ, которые относительно непроницаемы для молекул воды. В отсутствие АДГ моча не концентрируется и может выделяться в количествах, превышающих 20 л в сутки (норма 1,0-1,5 л в сутки). Связывание АДГ с V2(рис. 11-32) стимулирует аденилатциклазную систему и активацию протеинкиназы А. В свою очередь, протеинкиназа А фосфорилирует белки, стимулирующие экспрессию гена мембранного белка - аквапорина-2. Аквапорин-2 перемещается к апикальной мембране собирательных канальцев и встраивается в неѐ, образуя водные каналы. Это обеспечивает избирательную проницаемость мембраны клеток для воды, которые свободно диффундируют в клетки почечных канальцев и затем поступают в интерстициальное пространство. Поскольку в результате происходит реабсорбция воды из почечных канальцев и экскреция малого объѐма высококонцентрированной мочи (антидиурез), гормон называют антидиуретическим гормоном.
Рецепторы типа V1 локализованы в мембранах ГМК сосудов. Взаимодействие АДГ с рецептором V1 приводит к активации фосфолипазы С, которая гидролизует фосфатидилинозитол-4,5-бисфосфат с образованием инозитолтрифосфата и диацилглицерола. Инозитолтрифосфат вызывает высвобождение Са2+ из ЭР. Результатом действия гормона через рецепторы V1 является сокращение гладкомышечного слоя сосудов. Сосудосуживающий эффект АДГ проявляется при высоких концентрациях гормона. Поскольку сродство АДГ к рецептору V2 выше, чем к рецептору V1, при физиологической концентрации гормона в основном проявляется его антидиуретическое действие.
3. Несахарный диабет
Дефицит АДГ, вызванный дисфункцией задней доли гипофиза, а также нарушениями в системе передачи гормонального сигнала, приводит к развитию несахарного диабета. При этом происходит нерегулируемая экскреция воды, а наиболее опасным последствием является дегидратация организма.
Под названием "несахарный диабет" объединяют заболевания с разной этиологией. Так, основными причинами центрального несахарного диабета могут быть генетические дефекты синтеза препро-АДГ в гипоталамусе, дефекты процессинга и транспорта проАДГ (наследственная форма), а также повреждения гипоталамуса или нейрогипофиза (например, в результате черепно-мозговой травмы, опухоли, ишемии). Нефрогенный несахарный диабет возникает вследствие мутации гена рецептора АДГ типа V2 (наследственная форма), следствием которого является неспособность почек реагировать на гормон. Основное проявление несахарного диабета - гипотоническая полиурия, т.е. выделение большого количества мочи низкой плотности. Снижение секреции АДГ приводит также к усиленному потреблению воды. Диагностические критерии несахарного диабета: выраженная полиурия (до 20 л в сутки, плотность мочи
<1,010, в норме - 1,020).
Б. Альдостерон
Альдостерон - наиболее активный минералокортикостероид, синтезирующийся в коре надпочечников из холестерола.
Синтез и секреция альдостерона клетками клубочковой зоны непосредственно стимулируются низкой концентрацией Na+ и высокой концентрацией К+ в плазме крови. На секрецию альдостерона влияют также простагландины, АКТГ. Однако наиболее важное влияние на секрецию альдостерона оказывает ренинангиотензиновая система.
Альдостерон не имеет специфических транспортных белков, но за счѐт слабых взаимодействий может образовывать комплексы с альбумином. Гормон очень быстро захватывается печенью, где превращается в тетрагидроальдостерон-3-глюкуронид и экскретируется с мочой.
1. Механизм действия альдостерона
В клетках-мишенях гормон взаимодействует с рецепторами, которые могут быть локализованы как в ядре, так и в цитозоле клетки. Образовавшийся комплекс гормон-рецептор взаимодействует с определѐнным участком ДНК и изменяет скорость транскрипции специфических генов. Результат действия альдостерона - индукция синтеза: а) белков-транспортѐров Na+ из просвета канальца в эпителиальную клетку почечного канальца; б) Nа+,К+,-АТФ-азы, обеспечивающей удаление ионов натрия из клетки почечного канальца в межклеточное пространство и переносящей ионы калия из межклеточного пространства в клетку почечного канальца; в) белков-транспортѐров ионов калия из клеток почечного канальца в первичную мочу; г) митохондриальных ферментов ЦТК, в частности цитратсинтазы, стимулирующих образование молекул АТФ, необходимых для активного транспорта ионов .
Суммарным биологическим эффектом индуцируемых альдостероном белков является увеличение реабсорбции ионов натрия в канальцах нефронов, что вызывает задержку NaCl в организме, и возрастание экскреции калия.
2. Роль системы ренин-ангиотензин- альдостерон в регуляции водно-солевого обмена
Главным механизмом регуляции синтеза и секреции альдостерона служит система ренинангиотензин.
Ренин - протеолитический фермент, продуцируемый юкстагломерулярными клетками, расположенными вдоль конечной части афферентных (приносящих) артериол, входящих в почечные клубочки .
Юкстагломерулярные клетки особенно чувствительны к снижению перфузионного давления в почках. Уменьшение АД (кровотечение, потеря жидкости, снижение концентрации NaCl) сопровождается падением перфузионного давления в приносящих артериолах клубочка и соответствующей стимуляцией высвобождения ренина.
Субстратом для ренина служит ангиотензиноген. Ангиотензиноген - α2-глобулин, содержащий более чем 400 аминокислотных остатков. Образование ангиотензиногена происходит в печени и стимулируется глюкокортикоидами и эстрогенами. Ренин гидролизует пептидную связь в молекуле ангиотензиногена и отщепляет N-концевой декапептид (ангиотензин I), не имеющий биологической активности.
Под действием карбоксидипептидилпептидазы, или антиотензин-превращающего фермента (АПФ), выявленного в эндотелиальных клетках, лѐгких и плазме крови, с С-конца ангиотензина I удаляются 2 аминокислоты и образуется октапептид - ангиотензин II.
Ангиотензин II, связываясь со специфическими рецепторами, локализованными на поверхности клеток клубочковой зоны коры надпочечников и ГМК, вызывает изменение внутриклеточной концентрации диацилглицерола и инозитолтрифосфата. Инозитолтрифосфат стимулирует высвобождение из ЭР ионов кальция, совместно с которым активирует протеинкиназу С, опосредуя тем самым специфический биологический ответ клетки на действие ангиотензина П.
При участии аминопептидаз ангиотензин II превращается в ангиотензин III - гептапептид, проявляющий активность ангиотензина II. Однако концентрация гептапептида в плазме крови в 4 раза меньше концентрации октапептида, и поэтому большинство эффектов являются результатом действия ангиотензина П. Дальнейшее расщепление ангиотензина II и ангиотензина III протекает при участии специфических протеаз (ангиотензиназ).
Ангиотензин II оказывает стимулирующее действие на продукцию и секрецию альдостерона клетками клубочковой зоны коры надпочечников, который, в свою очередь, вызывает задержку ионов натрия и воды,
врезультате чего объѐм жидкости в организме восстанавливается. Кроме этого, ангиотензин II, присутствуя
вкрови в высоких концентрациях, оказывает мощное сосудосуживающее действие и тем самым повышает АД.
3. Восстановление объѐма крови при обезвоживании организма