

72. Депонирование и мобилизация жиров в жировой ткани, физиологическая роль этих процессов. Роль инсулина, адреналина и глюкагона в регуляции метаболизма жира.
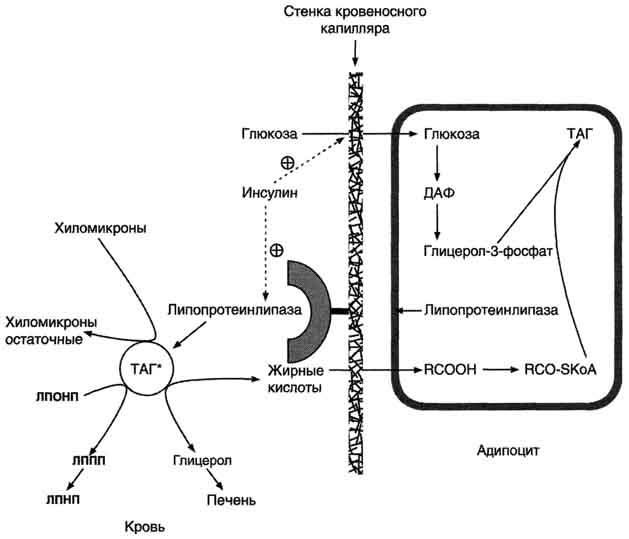 Депонирование
жира в адипоцитах в абсорбтивном
периоде. После
еды при повышении концентрации глюкозы
в крови увеличивается секреция инсулина.
Инсулин активирует транспорт глюкозы
внутрь адипоцитов, действуя на ГЛЮТ-4,
и синтез ЛП-липазы в адипоцитах и её
экспонирование на поверхности стенки
капилляров. ЛП-липаза, связанная с
эндотелием сосудов, гидролизует жиры
в составе ХМ и ЛПОНП. АпоС-II на поверхности
ХМ и ЛПОНП активирует ЛП-липазу. Жирные
кислоты проникают в адипоцит, а глицерол
транспортируется в печень. Так как в
адипоцитах нет фермента глицеролкиназы,
то свободный глицерол не может
использоваться для синтеза ТАГ в этой
ткани. Активированные жирные кислоты
взаимодействуют с глицерол-3-фосфатом,
образующимся из дигидроксиацетонфосфата,
и через фосфатидную кислоту превращаются
в ТАГ, которые депонируются в адипоцитах.
Сокращения: ТАГ* - триацилглицеролы в
составе ХМ и ЛПОНП; ДАФ - дигидроксиацетонфосфат.
Депонирование
жира в адипоцитах в абсорбтивном
периоде. После
еды при повышении концентрации глюкозы
в крови увеличивается секреция инсулина.
Инсулин активирует транспорт глюкозы
внутрь адипоцитов, действуя на ГЛЮТ-4,
и синтез ЛП-липазы в адипоцитах и её
экспонирование на поверхности стенки
капилляров. ЛП-липаза, связанная с
эндотелием сосудов, гидролизует жиры
в составе ХМ и ЛПОНП. АпоС-II на поверхности
ХМ и ЛПОНП активирует ЛП-липазу. Жирные
кислоты проникают в адипоцит, а глицерол
транспортируется в печень. Так как в
адипоцитах нет фермента глицеролкиназы,
то свободный глицерол не может
использоваться для синтеза ТАГ в этой
ткани. Активированные жирные кислоты
взаимодействуют с глицерол-3-фосфатом,
образующимся из дигидроксиацетонфосфата,
и через фосфатидную кислоту превращаются
в ТАГ, которые депонируются в адипоцитах.
Сокращения: ТАГ* - триацилглицеролы в
составе ХМ и ЛПОНП; ДАФ - дигидроксиацетонфосфат.
Б. Мобилизация жиров из жировой ткани
Адипоциты (место депонирования жиров) располагаются в основном под кожей, образуя подкожный жировой слой, и в брюшной полости, образуя большой и малый сальники. Мобилизация жиров, т.е. гидролиз до глицерола и жирных кислот, происходит в постабсорбтивный период, при голодании и активной физической работе. Гидролиз внутриклеточного жира осуществляется под действием фермента гормончувствительной липазы - ТАГ-липазы. Этот фермент отщепляет одну жирную кислоту у первого углеродного атома глицерола с образованием диацилглицерола, а затем другие липазы гидролизуют его до глицерола и жирных кислот, которые поступают в кровь. Глицерол как водорастворимое вещество транспортируется кровью в свободном виде, а жирные кислоты (гидрофобные молекулы) в комплексе с белком плазмы - альбумином.
В. Гормональная регуляция синтеза и мобилизации жиров
Какой процесс будет преобладать в организме - синтез жиров (липогенез) или их распад (липолиз), зависит от поступления пищи и физической активности. В абсорбтивном состоянии под действием инсулина происходит липогенез, в постабсорбтивном состоянии - липолиз, активируемый глюкагоном. Адреналин, секреция которого увеличивается при физической активности, также стимулирует липолиз.
Регуляция синтеза жиров. В абсорбтивный период при увеличении соотношения инсулин/глюкагон в печени активируется синтез жиров. В жировой ткани индуцируется синтез ЛП-липазы в адипоцитах и осуществляется её экспонирование на поверхность эндотелия; следовательно, в этот период увеличивается поступление жирных кислот в адипоциты. Одновременно инсулин активирует белки-переносчики глюкозы - ГЛЮТ-4. Поступление глюкозы в адипоциты и гликолиз также активируются. В результате образуются все необходимые компоненты для синтеза жиров: глицерол-3-фосфат и активные формы жирных кислот. В печени инсулин, действуя через различные механизмы, активирует ферменты путём дефосфорилирования и индуцирует их синтез. В результате увеличиваются активность и синтез ферментов, участвующих в превращении части глюкозы, поступающей с пищей, в жиры. Это - регуляторные ферменты гликолиза, пируватдегидрогеназный комплекс и ферменты, участвующие в синтезе жирных кислот из ацетил-КоА. Результат действия инсулина на обмен углеводов и жиров в печени - увеличение синтеза жиров и секреция их в кровь в составе ЛПОНП. ЛПОНП доставляют жиры в капилляры жировой ткани, где действие ЛП-липазы обеспечивает быстрое поступление жирных кислот в адипоциты, где они депонируются в составе триацилглицеринов.
Запасание жиров в жировой ткани - основная форма депонирования источников энергии в организме человека. Запасы жиров в организме человека массой 70 кг составляют 10 кг, но у многих людей количество жиров может быть значительно больше.
Жиры образуют в адипоцитах жировые вакуоли. Жировые вакуоли иногда заполняют значительную часть цитоплазмы. Скорость синтеза и мобилизации подкожного жира происходит неравномерно в разных частях организма, что связано с неодинаковым распределением рецепторов гормонов на адипоцитах.
Регуляция мобилизации жиров. Мобилизация депонированных жиров стимулируется глюкагоном и адреналином и, в меньшей степени, некоторыми другими гормонами (соматотропным, кортизолом). В постабсорбтивный период и при голодании глюкагон, действуя на адипоциты через аденилатциклазную систему, активирует протеинкиназу А, которая фосфо-рилирует и, таким образом, активирует гормончувствительную липазу, что инициирует липо-лиз и выделение жирных кислот и глицерина в кровь. При физической активности увеличивается секреция адреналина, который действует через β-адренергические рецепторы адипоцитов, активирующие аденилатциклазную систему. В настоящее время обнаружено 3 типа β-рецепторов: β1, β2, β3, активация которых приводит к липолитическому действию. К наибольшему липолитическому действию приводит активация β3-рецепторов. Адреналин одновременно действует и на α2-рецепторы адипоцитов, связанные с ингибирующим G-белком, что инактивирует аденилатциклазную систему. Вероятно, действие адреналина двояко: при низких концентрациях в крови преобладает его антилиполитическое действие через α2-рецепторы, а при высокой - преобладает липолитическое действие через β-рецепторы.
Для мышц, сердца, почек, печени при голодании или физической работе жирные кислоты становятся важным источником энергии. Печень перерабатывает часть жирных кислот в кетоновые тела, используемые мозгом, нервной тканью и некоторыми другими тканями как источники энергии.
В результате мобилизации жиров концентрация жирных кислот в крови увеличивается приблизительно в 2 раза, однако абсолютная концентрация жирных кислот в крови невелика даже в этот период. Т1/2 жирных кислот в крови тоже очень мал (менее 5 мин), что означает существование быстрого потока жирных кислот из жировой ткани к другим органам. Когда постабсорбтивный период сменяется аборбтивным, инсулин активирует специфическую фосфатазу, которая дефосфорилирует гормончувствительную липазу, и распад жиров останавливается.