

Лекция № 1. Предмет и задачи физиологии, ее связи с другими дисциплинами. Краткая история развития физиологии как науки. Методы физиологии. Общий план строения нервной системы и ее физиологическое значение. Основные физиологические понятия.
"Наша способность к самообмену по поводу работы собственного мозга почти безгранична главным образом потому, что часть, о которой мы можем сообщить, составляет лишь ничтожную долю того, что происходит у нас в голове".
Фрэнсис Крик
Физиология (греч. physis - природа) - это наука изучающая функции организма человека, его органов и систем, а также механизмы регуляции этих функций.
Вместе с анатомией физиология является основным разделом биологии.
Современная физиология представляет собой сложный комплекс общих и специальных научных дисциплин, таких как: общая физиология, физиология человека нормальная и патологическая, возрастная физиология, физиология животных, психофизиология и др.
Физиология изучает процессы жизнедеятельности, протекающие в организме на всех его структурных уровнях: клеточном, тканевом, органном, системном, аппаратном и организменном. Она тесно связана с дисциплинами морфологического профиля: анатомией, цитологией, гистологией, эмбриологией, так как структура и функция взаимно обусловливают друг друга. Физиология широко использует данные биохимии и биофизики для изучения функциональных изменений, происходящих в организме, и механизма их регуляции. Физиология также опирается на общую биологию и эволюционное учение, как основы для понимания общих закономерностей.
Для специалистов-психологов изучение физиологии имеет важное теоретическое и практическое значение. Работа их не может быть полноценной, если они не будут хорошо знать функциональные особенности нервной системы и закономерности высшей нервной деятельности человека.
Физиология прошла длинный и сложный путь развития, включающий в себя 7 периодов (смотри 1 лекцию по анатомии). Как и анатомия она возникла из потребностей медицины, постепенно расширяя свое прикладное значение для других наук: философии, педагогики, психологии.
Первоначальные представления о функциях организма были сформулированы врачами и учеными Древней Греции (Аристотель, Гиппократ), Древнего Рима (Гален), Древнего Китая (Хуанди, Бянь Цяо), Древней Индии и др. стран. Изучение ими строения тела проводилось одновременно с исследованиями функций организма.
В эпоху Средневековья естествознание было подчинено церкви и Святейшей инквизиции, поэтому период от 2 до 15 века называют периодом "сумерек истории".
В эпоху Возрождения в естествознании и медицине большое значение начали придавать опыту и наблюдению. Дальнейшее развитие физиологии связано с успехами анатомии, где работы Леонардо да Винчи и Андреаса Везалия подготовили почву для открытий в области физиологии.
Самостоятельной научной дисциплиной физиология стала к началу 17 века. Здесь важнейшее значение имело открытие Вильямом Гарвеем кругов кровообращения, исследование капилляров Марчелло Мальпиги, формулирование Рене Декартом понятия о рефлексе, учение Джакомо Борелли о механике движений. Большую роль сыграли в развитии физиологии успехи физики и химии. Правда, это нередко приводило к ложным механистическим выводам. Механицизм отождествлял физиологию с физикой и химией, способствовал появлению метафизического направления в науке, отрицавшего всякое развитие в природе. В биологии появилось антинаучное направление - витализм, утвердивший наличие в организме нематериальной "жизненной силы".
Значительного расцвета физиология достигла после великих открытий Ломоносова (закон сохранения вещества и превращения энергии), Шванна и Шлейдена (клеточная теория), Дарвина (эволюционное учение). В 19 и особенно 20 веке физиология обогатилась новыми открытиями. Клод Бернар создал представление о гомеостазе, изучил роль НС в регуляции тонуса сосудов и углеводного обмена. Дюбуа-Реймон явился основоположником электрофизиологии. Шеррингтон изучил физиологию спинного мозга. Капитальные исследования физиологии ВНС выполнил Кэннон. Создание приборов для исследования роли НС в регуляции дыхания, кровообращения и др. систем позволило выяснить, что процесс возбуждения всегда связан с электрическими изменениями в клетках и тканях. В противовес виталистическому направлению в физиологии получает развитие нервизм - прогрессивное направление, которое разработано русскими физиологами Сеченовым, Павловым, Боткиным, Бехтеревым, Введенским, Ухтомским.
Работа Сеченова "Рефлексы головного мозга" и павловское учение о сигнальных системах стали фундаментальной основой современной мировой физиологии.
В настоящее время физиологические исследования проводят в крупных научных институтах и лабораториях, в которых трудятся специалисты разных профилей (морфологи, биофизики, биохимики, психологи, математики, инженеры, биокибернетики).
Если еще недавно в физиологии применяли сравнительно простые приборы - кимографы, индукционные катушки, то сейчас применяется сложная аппаратура - электрокардиографы, спирографы, электроэнцефалографы, тепловизоры, компьютерная техника.
Большая заслуга в развитии современной физиологии принадлежит последователям и ученикам Павлова - Орбели, Быкову, Черниговскому, Анохину и др.
Академик Орбели создал учение об адаптационно-трофической функции симпатической нервной системы и внес большой вклад в развитие эволюционной и возрастной физиологии. Академик Быков исследовал влияние коры мозга на внутренние органы и установил два механизма регуляции их работы: пусковой и коорригирующей (исправляющей), соответственно потребностям организма в данных условиях. Академик Черниговский создал представление о двусторонней (прямой и обратной) связи внутренних органов с корой мозга, возглавлял исследования в области космической физиологии и медицины. Академик Анохин создал учение о функциональных системах организма, в котором изложены современные представления об организации поведения живых организмов.
Процесс физиологической регуляции является основой самоудовлетворения потребностей живого организма. Потребности удовлетворяются благодаря деятельности управляющих систем - нервной и эндокринной.
Для удовлетворения своих потребностей в условиях изменений внешней среды организму необходимо:
1. ставить определенные задачи;
2. достигать намеченного результата.
Согласно учению Анохина, именно полезный результат является фактором определяющим поведение и образующим функциональную систему (ФУС). ФУС формируется как группа взаимосвязанных нейронов, обеспечивающих достижение полезного результата. В задачу ФУС входит выявление и оценка результата действия.
Компонентами ФУС являются:
1 - полезный результат,
2 - рецепторы,
3 - нервный центр,
4 - исполнительные механизмы,
5 - пути обратной связи для сообщения результата действия.
Отчетность исполнительных органов перед центрами обеспечивает оценку результата и внесение поправок в работу ФУС, если результат не достигнут.
Фус регуляции движений

Физиология - это экспериментальная наука. Она использует два основных метода: наблюдение и эксперимент.
Наблюдение - основной метод познания окружающего и используется в любом научном исследовании. Его недостатком является пассивность исследователя, который может выяснить лишь внешнюю сторону явления, например - работу (функцию) органа. Механизм регуляции работы органа можно выяснить только опытным путем.
Эксперимент позволяет исследователю создать определенные условия, в которых выясняются количественные и качественные характеристики того ил иного явления.
Эксперимент может быть острым или хроническим. Острый опыт (вивисекция) позволяет в короткое время изучить какой-либо регуляторный механизм, срабатывающий в экстремальных для подопытного организма ситуациях. Хронический эксперимент позволяет длительное время исследовать механизмы регуляции в условиях нормального взаимодействия организма и среды.
В опытах на животных используют хирургические методы - экстирпацию (удаление) или пересадку органов, вживление электродов, датчиков. Объективным методом является метод телеметрии, позволяющий регистрировать параметры процесса или явления на расстоянии.
Экспериментальные исследования в последние годы проводят с помощью сложной оптической, радиотехнической, электронной аппаратуры, позволяющей, одновременно изучать десятки функций, их изменения во взаимодействии, т.е. комплексно.
Обработка полученного массива данных происходит с применением методов математической статистики и компьютерной техники.
Нервная система состоит из центрального и периферического отделов. ЦНС включает в себя головной и спинной мозг, а ПНС - это все нервы и узлы, лежащие за пределами ЦНС.
Различают также соматическую и вегетативную нервную систему. Первая регулирует работу скелетных мышц и органов чувств. Вторая регулирует работу внутренних органов и желез.
Выделение вышеназванных отделов в нервной системе является условным, удобным для изучения в определенной логической последовательности. В действительности нервная система представляет собой анатомически и функционально единое целое, элементарной основой которого являются нейроны.
Нервная система является ведущей физиологической системой организма, главной системой управления. Это подтверждается тем, что НС плода начинает функционировать задолго до его рождения (Сайенс Ньюс, № 16, 1984).
Функции нервной системы можно поделить на два типа: высшие и низшие.
Низшая нервная деятельность представляет собой процессы регуляции работы органов и систем в организме.
Высшая нервная деятельность включает в себя те функциональные механизмы мозга, которые обеспечивают организму соответствующий контакт с окружающей средой. Высшие функции лежат в основе психической деятельности человека, формировании свойств личности: темперамента, характера, способностей, потребностей и интересов. Высшая нервная деятельность требует оперативного и адекватного изменения в режиме работы внутренних органов. Следовательно, высшая и низшая нервная деятельность накладываются друг на друга и должны рассматриваться в тесном гармоничном единстве.
Активность животных и человека проявляется в виде функций и физиологических актов.
Функция это специфическая деятельность клеток, тканей, органов. Например, функцией мышцы является сокращение, железы - секреция, нейронов - генерирование и проведение импульсов. За счет изменения функций организм приспосабливается к изменениям условий существования.
Все функции можно разделить на:
1) соматические (животные), которые осуществляются за счет деятельности скелетных мышц, иннервируемых СНС;
2) вегетативные (растительные), которые связаны с обменом веществ, ростом и размножением. Они осуществляются за счет работы внутренних органов, иннервируемых ВНС.
Физиологический акт - сложный процесс, который осуществляется с участием различных систем организма (физиологические акты дыхания, пищеварения, выделения, дыхания и т.д.). Например, физиологический акт пищеварения включает в себя возбуждение сенсорных отделов ЦНС (зрительных, обонятельных, вкусовых, тактильных), двигательных центров (добывание, обработка и приготовление пищи), секреторного аппарата ЖКТ (выделение пищеварительных соков), гладких мышц ЖКТ (моторика, перистальтика), кишечного эпителия (всасывание). Таким образом, акт пищеварения обеспечивается проявлением сложных и многочисленных функций на клеточном, тканевом, органном и системном уровнях, которые включаются в функциональную систему (ФУС) и обеспечивают достижение полезного результата.
Лекция № 2. Понятие о возбудимых тканях. Возбуждение. Возбудимость. Проводимость. Рефрактерность и лабильность. Физиологические свойства нервных волокон (безмиелиновых и миелиновых). Утомляемость нервного волокна. Физиологические свойства синапсов.
"Все регулируется, течет по расчищенным руслам, совершает свой кругооборот в соответствии с законом и под его защитой".
И. Ильф и Е. Петров "Золотой теленок"
Все клетки и ткани живого организма под действием раздражителей переходят из состояния относительного физиологического покоя в состояние активности (возбуждения). Наибольшая степень активности наблюдается в нервной и мышечной ткани.
Главными свойствами возбудимых тканей являются: I. возбудимость, II проводимость, III рефрактерность и лабильность, которые связаны с одним из самых общих свойств живого - раздражимостью.
Изменения в окружающей среде или организме называют раздражителями, а их действие - раздражением.
По природе раздражители бывают: механические, химические, электрические, температурные.
По биологическому признаку раздражители делятся на:
1. адекватные, которые воспринимаются соответствующими специализированными рецепторами (глаза - свет, уха - звук, кожа - боль, температура, прикосновение, давление, вибрация);
2. неадекватные, к которым специализированные рецепторы не приспособлены, но воспринимают их при чрезмерной силе и длительности (удар - глаз - свет).
Наиболее общим, адекватным и естественным раздражителем для всех клеток и тканей организма является нервный импульс.
Основные физиологические свойства нервной ткани (возбудимость, проводимость, рефрактерность и лабильность) характеризуют функциональное состояние нервной системы человека, определяют его психические процессы.
I. Возбудимость - способность живой ткани отвечать на действие раздражителя возникновением процесса возбуждения с изменением физиологических свойств.
Количественной мерой возбудимости является порог возбуждения, т.е. минимальная величина раздражителя, способная вызвать ответную реакцию тканей.
Раздражитель меньшей силы называют подпороговым, а большей - надпороговым.
Возбудимость представляет собой, в первую очередь, изменение обмена веществ в клетках тканей. Изменение обмена веществ сопровождается переходом через клеточную мембрану отрицательно и положительно заряженных ионов, которые изменяют электрическую активность клетки. Разность потенциалов в покое между внутренним содержимым клетки и клеточной оболочкой, составляющая 50-70 мВ (миллиВольт) называется мембранным потенциалом покоя.
 Основой
этого состояния клетки являетсяизбирательная
проницаемость мембраны
по отношению к ионам К+
и Na+.
Ионам Na+,
находящимся во внеклеточной среде,
через мембрану в клетки путь закрыт, а
К+
свободно проникает через поры клеточной
мембраны из цитоплазмы клетки в тканевую
жидкость. В результате в цитоплазме
остаются отрицательно заряженные ионы,
а на поверхности мембраны накапливаются
положительно заряженные ионы К+
и Na+.
Основой
этого состояния клетки являетсяизбирательная
проницаемость мембраны
по отношению к ионам К+
и Na+.
Ионам Na+,
находящимся во внеклеточной среде,
через мембрану в клетки путь закрыт, а
К+
свободно проникает через поры клеточной
мембраны из цитоплазмы клетки в тканевую
жидкость. В результате в цитоплазме
остаются отрицательно заряженные ионы,
а на поверхности мембраны накапливаются
положительно заряженные ионы К+
и Na+.
При
возбуждении клетки проницаемость ионов
Na+
резко увеличивается, и они устремляются
в цитоплазму, снижая потенциал покоя
до нуля, а затем увеличивая разность
потенциалов противоположного значения
до 80-110 мВ. Такое
кратковременное (0,004-0,005 сек) изменение
разности потенциалов называется
потенциалом действия
(спайком);
англ. s pike
- острие.
pike
- острие.
Вслед за этим нарушенное равновесие ионов вновь восстанавливается. Для этого существует специальный клеточный механизм - "натрий-калиевый насос", который обеспечивает активное "выкачивание" Na+ из клетки и «нагнетание» в нее К+. Таким образом, существуют 2 типа движения ионов через клеточную мембрану:
1 - пассивный ионный транспорт по градиенту концентрации ионов;
2 - активный ионный транспорт против градиента концентрации, осуществляемый "натрий-калиевым насосом" с затратой энергии АТФ.
Вывод: возбуждение нервной клетки связано с изменением обмена веществ и сопровождается появлением электрических потенциалов (нервных импульсов).
Проводимость - способность живой ткани проводить волны возбуждения - биоэлектрические импульсы.
 Для
обеспечения гомеостатического единства
все структуры организма (клетки, ткани,
органы и т.д.) должны иметь возможностьпространственного
взаимодействия.
Распространение возбуждения от места
его возникновения до исполнительных
органов - один из основных способов
такого взаимодействия. Возникший в
месте нанесения раздражения потенциал
действия является причиной раздражения
соседних, невозбужденных участков
нервного (или мышечного) волокна.
Благодаря этому явлению волна потенциала
действия создает ток
действия,
который распространяется по всей длине
нервного волокна. В безмиелиновых
нервных волокнах возбуждение проводится
с некоторым затуханием - декрементом,
а в миелиновых нервных волокнах - без
затухания. Проведение возбуждения также
сопровождается изменением обмена
веществ и энергии.
Для
обеспечения гомеостатического единства
все структуры организма (клетки, ткани,
органы и т.д.) должны иметь возможностьпространственного
взаимодействия.
Распространение возбуждения от места
его возникновения до исполнительных
органов - один из основных способов
такого взаимодействия. Возникший в
месте нанесения раздражения потенциал
действия является причиной раздражения
соседних, невозбужденных участков
нервного (или мышечного) волокна.
Благодаря этому явлению волна потенциала
действия создает ток
действия,
который распространяется по всей длине
нервного волокна. В безмиелиновых
нервных волокнах возбуждение проводится
с некоторым затуханием - декрементом,
а в миелиновых нервных волокнах - без
затухания. Проведение возбуждения также
сопровождается изменением обмена
веществ и энергии.

III.
Рефрактерность
- временное снижение возбудимости ткани,
возникающее при появлении потенциала
действия. В этот момент повторные
раздражения не вызывают ответной реакции
(абсолютная рефрактерность). Она длится
не более 0,4 миллисекунды, а затем наступает
фаза относительной
рефракторности,
когда раздражение может вызвать слабую
реакцию. Эта фаза сменяется фазой
повышенной возбудимости - супернормальности.
III.
Рефрактерность
- временное снижение возбудимости ткани,
возникающее при появлении потенциала
действия. В этот момент повторные
раздражения не вызывают ответной реакции
(абсолютная рефрактерность). Она длится
не более 0,4 миллисекунды, а затем наступает
фаза относительной
рефракторности,
когда раздражение может вызвать слабую
реакцию. Эта фаза сменяется фазой
повышенной возбудимости - супернормальности.
Такая динамика возбудимости обусловлена процессами изменения и восстановления равновесия ионов на мембране клетки.
Профессор Н.Е. Введенский исследовал особенности этих процессов и установил, что возбудимые ткани могут отвечать разным числом потенциалов действия на определенную частоту раздражений. Он назвал это явление лабильностью (функциональной подвижностью).
Лабильность - свойство возбудимой ткани воспроизводить максимальное число потенциалов действия в единицу времени.
Максимальная лабильность - у нервной ткани. Частота раздражений, вызывающая максимальную реакцию называется оптимальной (лат. optimum - наилучший), а вызывающая угнетение реакции - пессимальной (лат. pessimum - наихудший).
*Нервное волокно - до 1000 имп./сек, мышца - 200-250 имп./сек., синапс - до 100-125 имп./сек.
Пессимум - активная реакция ткани, направленная на защиту ее от чрезмерных раздражений. Это одна из форм проявления торможения. Возбуждение и торможение это противоположные по значению саморегулирующиеся процессы, которые устанавливают "золотую середину" уровня отношений организма со средой.
Нервные волокна (отростки нервных клеток) обладают всеми свойствами возбудимых тканей, а проведение нервных импульсов является их специальной функцией. Скорость проведения возбуждения зависит от:
1 - диаметра волокон (толще быстрее),
2 - строения их оболочки.
 Безмиелиновые
(безмякотные) волокна покрыты
только леммоцитами
(шванновскими клетками). Между ними и
осевым цилиндром (аксоном нейрона)
имеется щель с межклеточной жидкостью,
поэтому, клеточная мембрана остается
неизолированной. Импульс распространяется
по волокну со скоростью всего 1-3 м/сек.
Безмиелиновые
(безмякотные) волокна покрыты
только леммоцитами
(шванновскими клетками). Между ними и
осевым цилиндром (аксоном нейрона)
имеется щель с межклеточной жидкостью,
поэтому, клеточная мембрана остается
неизолированной. Импульс распространяется
по волокну со скоростью всего 1-3 м/сек.
Миелиновые волокна покрыты спиральными слоями шванновских клеток с прослойкой миелина - жироподобного вещества с высоким удельным сопротивлением. Миелиновая оболочка через промежутки равной длины прерывается, оставляя оголенными участки осевого цилиндра длиной 1 мкм.
И з-за
такого строения электрические токи
могут входить в волокна и выходить из
них только в области неизолированных
участков -перехватов
Ранвье. При
нанесении раздражения в ближайшем
перехвате возникает деполяризация, а
соседние перехваты поляризованы. Между
ними возникает разность потенциалов,
которая приводит к появлению круговых
токов действия.
з-за
такого строения электрические токи
могут входить в волокна и выходить из
них только в области неизолированных
участков -перехватов
Ранвье. При
нанесении раздражения в ближайшем
перехвате возникает деполяризация, а
соседние перехваты поляризованы. Между
ними возникает разность потенциалов,
которая приводит к появлению круговых
токов действия.
Т аким
образом, импульс в миелиновом волокне
проходит скачкообразно (сальтаторно)
от перехвата к перехвату. Возбуждение
при этом распространяется без затухания,
а скорость проведения импульса достигает
120-130 м/сек.
аким
образом, импульс в миелиновом волокне
проходит скачкообразно (сальтаторно)
от перехвата к перехвату. Возбуждение
при этом распространяется без затухания,
а скорость проведения импульса достигает
120-130 м/сек.
При нанесении раздражения на нервное волокно происходит двустороннее распространение возбуждения - в центростремительном и центробежном направлении. Это не противоречит принципу одностороннего проведения импульсов, и объясняется первичностью появления возбуждения в рецепторах или нервных центрах, а также наличием синапсов. Нейротрансмиттер (медиатор) содержится только в пресинаптическом аппарате и переносит потенциал только однонаправленно (см. лекцию по анатомии № 2).
Возбуждение проводится не только в нужном направлении, но и по одному изолированному волокну, не распространяясь на соседние волокна. Это обуславливает строго координированную рефлекторную деятельность. Например, седалищный нерв диаметром до 12 мм несет в себе тысячи нервных волокон (миелиновых и безмиелиновых, чувствительных и двигательных, соматических и вегетативных). В случае неизолированного проведения возбуждения наблюдалась бы хаотическая ответная реакция.
Изолированное проведение возбуждения в миелиновых волокнах обеспечивается миелиновой оболочкой, а в безмиелиновых - высоким удельным сопротивлением окружающей межклеточной жидкости (отсюда и затухание потенциала).
Н.Е. Введенский в 1883 году впервые установил, что нерв малоутомляем. Малая утомляемость нервных волокон объясняется тем, что энергетические затраты в них при возбуждении незначительны, а процессы восстановления протекают быстро. В организме нервные волокна работают также с недогрузкой. Например, двигательное волокно высоколабильно и может проводить до 2500 имп./сек. Из нервных же центров поступает не более 50-40 имп./сек.
Вывод: практическая неутомляемость нервных волокон связана с небольшими энергетическими затратами, с высокой лабильностью нервных волокон, с постоянной недогрузкой волокон.
Синапсы (см. строение в лекции по анатомии № 2) обладают следующими физиологическими свойствами:
 1
-одностороннее
проведение возбуждения,
которое связано с особенностями строения
самого синапса,
1
-одностороннее
проведение возбуждения,
которое связано с особенностями строения
самого синапса,
2 - синаптическая задержка, которая связана с затратой времени на освобождение и диффузию нейротрансмиттера через синаптическую щель, временем взаимодействия нейространсмиттера с соответствующим белком-рецептором.
В 1901 году Н.Е. Введенский ввел в физиологию нервной системы понятие о парабиозе. Эта своеобразная реакция на повреждающее воздействие оказалось универсальной для возбудимых тканей. При контузии нервного волокна, отравлении его фенолом, кокаином, поражением электротоком резко снижается лабильность. Парабиоз протекает в 3 стадии:
1 - провизорная (уравнительная) - мышца отвечает одинаковыми сокращениями как на сильные, так и на слабые импульсы, проводимые по поврежденному нерву,
2 - парадоксальная - частые импульсы вызывают слабые сокращения, а редкие - более или менее сильные сокращения,
3 - тормозная - теряется проводимость по нерву из-за блокирующего возбуждения (деполяризации) поврежденного участка нервного волокна.
Лекция № 3. Эмбиогенез нервной системы. Нервная пластинка, нервный желобок, нервная трубка. Ганглиозная пластинка. Стадия трех мозговых пузырей. Стадия пяти мозговых пузырей. Некоторые аспекты развития мозга в постнатальный период.
"С тех пор, как ваш мозг предается изучению наук, серьезным вычислениям, он увеличился в объеме".
А. Дюма "Виконт де Бражелон"
Для живых организмов, начиная от плоских червей и до высших позвоночных животных характерна трехслойность, т.е. развитие из трех зародышевых листков (экто-, мезо- и эктодермы).

НС человека развивается из наружного
зародышевого листка - эктодермы.
На дорзальной стороне туловища зародыша
клетки эктодермы дифференцируются и
формируют медуллярную
(нервную) пластинку.
НС человека развивается из наружного
зародышевого листка - эктодермы.
На дорзальной стороне туловища зародыша
клетки эктодермы дифференцируются и
формируют медуллярную
(нервную) пластинку.
Первоначально она состоит из одного слоя клеток, которые в дальнейшем разделяются на:
1 - нейробласты (из которых развиваются нейроны);
2 - спонгиобласты (из которых развивается нейроглия).
В следствие
неравномерности деления клеток пластинка
прогибается, постепенно превращаясь внервный
желобок.
Рост его боковых отделов приводит к
сближению, а затем, и к смыканию краев
желобка. Так формируется нервная
трубка.
Сращение в первую очередь происходит
в переднем (краниальном), а потом и в
заднем (каудальном) отделах нервной
трубки. На переднем и заднем концах
трубки остаются открытыми небольшие
отверстия - нейропоры.
После сращения нервная трубка отделяется
от экдотермы и погружается в мезодерму.
Полость трубки называется невроцель.
следствие
неравномерности деления клеток пластинка
прогибается, постепенно превращаясь внервный
желобок.
Рост его боковых отделов приводит к
сближению, а затем, и к смыканию краев
желобка. Так формируется нервная
трубка.
Сращение в первую очередь происходит
в переднем (краниальном), а потом и в
заднем (каудальном) отделах нервной
трубки. На переднем и заднем концах
трубки остаются открытыми небольшие
отверстия - нейропоры.
После сращения нервная трубка отделяется
от экдотермы и погружается в мезодерму.
Полость трубки называется невроцель.
К
 моменту смыкания трубки она состоит изтрех слоев.
Внутренний слой образован эпендимоцитами,
которые выстилают в дальнейшем полости
желудочков мозга и центрального канала
спинного мозга. Средний (плащевой) слой
будет формировать серое вещество мозга.
Наружный слой превращается в белое
вещество, так как содержит отростки
клеток. Боковые (латеральные) отделы
трубки развиваются более интенсивно,
при этом дорзальный и вентральный отделы
остаются в глубине. Так образуются
передняя и задняя срединные борозды, а
нервная трубка становится билатерально
симметричной. Со стороны невроцели
имеются небольшие продольные боковые
бороздки , которые делят боковые отделы
трубки на вентральную
(основную) и дозральную
(крыльную) пластинки.
моменту смыкания трубки она состоит изтрех слоев.
Внутренний слой образован эпендимоцитами,
которые выстилают в дальнейшем полости
желудочков мозга и центрального канала
спинного мозга. Средний (плащевой) слой
будет формировать серое вещество мозга.
Наружный слой превращается в белое
вещество, так как содержит отростки
клеток. Боковые (латеральные) отделы
трубки развиваются более интенсивно,
при этом дорзальный и вентральный отделы
остаются в глубине. Так образуются
передняя и задняя срединные борозды, а
нервная трубка становится билатерально
симметричной. Со стороны невроцели
имеются небольшие продольные боковые
бороздки , которые делят боковые отделы
трубки на вентральную
(основную) и дозральную
(крыльную) пластинки.
 Из
основной пластинки образуютсяпередние
канатики
белого вещества и передние
столбы серого
вещества. Отростки клеток передних
столбов выходят из нервной трубки,
образуя передние
корешки
(двигательные) спинного мозга. Из крыльной
пластинки развиваются здание
канатики
белого вещества и задние
столбы серого
вещества.
Из
основной пластинки образуютсяпередние
канатики
белого вещества и передние
столбы серого
вещества. Отростки клеток передних
столбов выходят из нервной трубки,
образуя передние
корешки
(двигательные) спинного мозга. Из крыльной
пластинки развиваются здание
канатики
белого вещества и задние
столбы серого
вещества.
Еще на стадии нервного желобка в его боковых отделах выделяются боковые клеточные тяжи - медуллярные гребешки. Эти гребешки формируют между экдотермой и нервной трубкой ганглиозную пластинку. В последствии она вторично разделяется на два ганглиозных валика, которые смещаются по бокам нервной трубки и сегментируются. Эти парные сегменты превращаются в спинномозговые узлы, соответственно сегментам туловища, а также в чувствительные узлы черепных нервов. Клетки, выселившиеся из ганглиозных валиков, формируют периферические отделы ВНС.
 В
краниальном отделе нервной трубки
образуется расширение (зачаток головного
мозга). Остальные отделы нервной трубки
формируют спинной мозг. Нейробласты
спинномозговых узлов (биполярные клетки)
имеют центральные отростки, которые
прорастают в спинной мозг и образуютзадние корешки
(чувствительные). Периферические отростки
нейробластов прорастают в ткани и
заканчиваются там рецепторами различных
типов.
В
краниальном отделе нервной трубки
образуется расширение (зачаток головного
мозга). Остальные отделы нервной трубки
формируют спинной мозг. Нейробласты
спинномозговых узлов (биполярные клетки)
имеют центральные отростки, которые
прорастают в спинной мозг и образуютзадние корешки
(чувствительные). Периферические отростки
нейробластов прорастают в ткани и
заканчиваются там рецепторами различных
типов.
В каудальном отделе нервная трубка редуцируется (укорачивается) и постепенно суживается, образуя концевую нить (filum terminale). Позвоночник эмбриона удлиняется более интенсивно, чем спинной мозг, поэтому корешки спинномозговых нервов в нижних отделах меняют горизонтальное направление на косое, а затем и вертикальное, образуя конский хвост (cauda equina). Нижний конец спинного мозга у новорожденного "поднимается" на уровень III поясничного позвонка.
Расширение краниального отдела нервной трубки к 4-м неделям эмбрионального развития превращается в три мозговых пузыря:
 1
-prosencephalon
(передний мозг);
1
-prosencephalon
(передний мозг);
2 - mesencephalon (средний мозг);
3 - rhombencephalon (ромбовидный мозг).
 В
конце 4-й недели начинается стадияпяти
мозговых пузырей:
В
конце 4-й недели начинается стадияпяти
мозговых пузырей:
1 - telencephalon (конечный мозг);
2 - diencephalon (промежуточный мозг;
3 - mesencephalon (средний мозг;
4 - metencephalon (задний мозг);
5 - myelencephalon seu medulla oblongata (продолговатый мозг), которые к 8-й неделе развития приобретают конкретные формы и изгибы. Продолговатый и задний мозг формируются путем деления ромбовидного мозга. Его общая полость превращается в четвертый (IV) желудочек. Ниже его (в спинном мозге) полостью является узкий центральный канал, а выше (в среднем мозге) - водопровод мозга (сильвиев). Полостью промежуточного мозга является третий желудочек (III), который через водопровод соединяется с четвертым (IV) желудочком.
 Конечный
мозг на ранних этапах развития состоит
из непарного мозгового пузыря, а за счет
преимущественного развития боковых
отделов превращается в два пузыря -
будущие полушария мозга. Непарная в
начале полость конечного мозга также
делится на двабоковых
желудочка,
которые сообщаются с третьим желудочком
с помощью межжелудочковых отверстий
Монро.
Конечный
мозг на ранних этапах развития состоит
из непарного мозгового пузыря, а за счет
преимущественного развития боковых
отделов превращается в два пузыря -
будущие полушария мозга. Непарная в
начале полость конечного мозга также
делится на двабоковых
желудочка,
которые сообщаются с третьим желудочком
с помощью межжелудочковых отверстий
Монро.
Боковые желудочки вследствие сложного развития полушарий имеют сложную конфигурацию.

 Интенсивный
рост полушарий приводит к тому, что они
постепенно покрывают сверху и с боков
промежуточный, средний мозг и мозжечок.
На внутренней поверхности полушарий
развиваютсябазальные
ядра.
Неравномерный рост стенок пузырей
конечного мозга приводит к появлению
на их наружной поверхности основных
борозд
полушарий. Ранее других появляются
латеральные (сильвиевы) борозды, а затем
центральные (роландовы) борозды. Эти
борозды разделяют полушария на доли. В
пределах долей мозга более мелкие
борозды разделяют отдельные участки
поверхности полушарий на извилины
полушарий.
Интенсивный
рост полушарий приводит к тому, что они
постепенно покрывают сверху и с боков
промежуточный, средний мозг и мозжечок.
На внутренней поверхности полушарий
развиваютсябазальные
ядра.
Неравномерный рост стенок пузырей
конечного мозга приводит к появлению
на их наружной поверхности основных
борозд
полушарий. Ранее других появляются
латеральные (сильвиевы) борозды, а затем
центральные (роландовы) борозды. Эти
борозды разделяют полушария на доли. В
пределах долей мозга более мелкие
борозды разделяют отдельные участки
поверхности полушарий на извилины
полушарий.
Наружные слои стенок пузырей образованы серым веществом - корой мозга. Складчатость коры значительно увеличивает ее поверхность. Площадь коры у взрослого человека составляет площадь одной страницы полноразмерной газеты - 2400 кв. см. Рельеф полушария, с его бороздами и извилинами, индивидуален для каждого человека. У женщин количество борозд и извилин несколько больше, чем у мужчин вследствие меньших размеров черепа. Окончательное формирование головного мозга завершается к 20 годам. Следует отметить, что абсолютная масса мозга не определяет умственные способности человека. При средней массе 1375 г допускаются индивидуальные колебания в пределах от 1100 до 2000 и более г. Мозг Карла Гаусса весил 2400 г, Оливера Кромвеля - 2300 г, лорда Байрона - 2238г, Тургенева - 1012г, Есенина - 1920 г, Жоржа Кювье - 1872 г, канцлера Бисмарка - 1800 г, Людвига ван Бетховена - 1750 г, академика Сахарова и Гельмгольца - 1440г, И.П. Павлова - 1653 г, Д.И. Менделеева 1571 г, А.Ф. Кони - 1100 г, Либиха - 1362 г, Анатоля Франса - 1017 г, У. Теккерея - 1644 г, Ф. Шиллера - 1871 г. Установлено, что интеллект человека снижается только тогда, когда масса мозга уменьшается до 900 г и менее. Интеллектуальные возможности мозга зависят от качественного состояния нервных клеток, богатства синаптических связей между ними, но не от его абсолютной массы.
Развитие головного мозга идет гетерохронно. Прежде всего созревают те структуры, которые отвечают за нормальную жизнедеятельность организма на конкретном возрастном этапе. Функциональной полноценности достигают прежде всего стволовые, подкорковые и корковые структуры, которые регулируют вегетативные функции организма. Они в целом сформированы уже к 2-4 годам жизни. В первые месяцы после рождения регуляция основных жизненных функций осуществляется преимущественно таламо - стрио - паллидарной системой. Кора мозга, как высший отдел ЦНС, в целом формируется к 6 годам жизни, а окончательно формируется к 20 годам. Процесс обучения способствует прогрессивному развитию сети нейронов с увеличением синаптических связей. Это значительно увеличивает качество интеллекта несмотря на то, что после рождения количество нейронов может только уменьшаться, так как нейробласты прекращают свое деление в самом начале постнатального периода. У людей старшего, пожилого и старческого возраста сохранение интеллекта вероятно зависит от включения в условно-рефлекторную деятельность и процессы памяти помимо нейронов еще и клеток нейроглии. Об этом свидетельствует ряд исследовательских данных последнего времени.
Качественные изменения претерпевают и нервные волокна, составляющие белое вещество ЦНС. В первую очередь начинается миелинизация нервных волокон в филогенетически более старых отделах мозга, чем в более новых. В полушариях мозга миелинизация начинается раньше у волокон, проводящих различные виды чувствительности, а также осуществляющих связи с подкорковыми ядрами. Миелинизация афферентных волокон начинается примерно с 2-х месяцев жизни и заканчивается к 4-5 годам, а эфферентных волокон - несколько позже, в период от 4-5 месяцев до 7-8 лет.
Миелинизация волокон в ассоциативных зонах коры лобных, теменных и височных долей, которые являются наиболее молодыми в эволюционном отношении и обеспечивают сложнейшие формы интеллектуальной деятельности (речь, мышление, принятие решений), может продолжаться до 30-35 летнего возраста, а возможно, и до конца жизни. Это характерно только для человека (А.Л. Рылов).
Лекция № 4. Понятие о нервных центрах. Морфологические и функциональное определение нервного центра. Свойства нервных центров.
Нервный центр - это центральная часть рефлекторной дуги.
Анатомический нервный центр - это совокупность нервных клеток, выполняющих общую для них функцию и лежащих в определенном отделе ЦНС.
В функциональном отношении нервный центр это сложное объединение нескольких анатомических нервных центров, расположенных в разных отделах ЦНС и обусловливающих сложнейшие рефлекторные акты.
А.А. Ухтомский называл такие объединения "созвездиями" нервных центров. Различные анатомические нервные центры объединяются в ФУС для получения определенного полезного результата.
Нервные центры также непосредственно реагируют на БАВ, содержащиеся в протекающей через них крови (гуморальные влияния).
Для выявления функций нервных центров используют ряд методов:
1. метод электродного раздражения;
2. метод экстирпации (удаления, для нарушения исследуемой функции);
3. электрофизиологический метод регистрации электрических явлений в нервном центре и др.
Свойства нервных центров в значительной мере связаны с обилием синапсов и с особенностями проведения импульсов через них. Именно синаптические контакты определяют основные свойства нервных центров:
1 - односторонность проведения возбуждения;
2 - замедление проведения нервных импульсов;
3 - суммацию возбуждений;
4 - усвоению и трансформацию ритма возбуждений;
5 - следовые процессы;
6 - быструю утомляемость.
Одностороннее проведение возбуждения означает распространение импульса только в одном направлении - от чувствительного нейрона к двигательному. Это обусловлено синапсами, где проведение информации с помощью нейротрансмиттеров (медиаторов) идет от пресинаптической мембраны через синаптическую щель к постсинаптической мембране. Обратное проведение невозможно, чем достигается направленность потоков информации в организме.
Замедление проведения импульсов связано с тем, что электрический способ передачи информации в синапсах сменяется химическим (медиаторным) способом, который в тысячу раз медленнее. Время синаптической задержки в мотонейронах соматической НС составляет 0,3 мс. В вегетативной НС такая задержка более длительна, т.е. не менее 10 мс.
Множество синапсов на пути нервного импульса обеспечивают суммарную задержку, когда время задержки - центральное время проведения увеличивается до сотен и более мс.
Например, время реакции водителя на включение красного света светофора составляет не менее 200 мс, а при утомлении может превышать 1000 мс. Время от начала действия раздражителя до начала ответной реакции называется временем реакции или латентным (скрытым) временем рефлекса.
 Суммация
возбуждений
была открыта И.М. Сеченовым в 1863 году. В
нервном центре различают два вида
суммации:
Суммация
возбуждений
была открыта И.М. Сеченовым в 1863 году. В
нервном центре различают два вида
суммации:
временная (последовательная);
2. пространственная.
Временная суммация возникает при последовательном поступлении к постсинаптической мембране нейрона серии импульсов, в отдельности не вызывающих возбуждение нейрона. Сумма этих импульсов достигает пороговой величины раздражения и только после этого вызывает появление потенциала действия.
Пространственная суммация наблюдается при одновременном поступлении к нейрону нескольких слабых импульсов, которые в сумме достигают пороговой величины и вызывают появление потенциала действия.
Усвоение и трансформация ритма возбуждений в нервных центрах были изучены А.А. Ухтомским и его учениками (Голиковым, Жуковым и др.). Нейроны способны настраиваться на ритм раздражений как на более высокий, так и на более низкий. В результате такой способности нервные клетки сонастраиваются, работают сообща в едином ритме. Это имеет большое значение для взаимодействия между различными нервными центрами и создания временных ФУС для достижения определенного полезного результата. С другой стороны, нейроны способны трансформировать (изменять) ритм поступающих к ним импульсов в собственный ритм.
Следовые процессы или последействие означает, что после окончания действия раздражителя активное состояние нервного центра продолжается еще некоторое время. Длительность следовых процессов различна. В спинном мозге - несколько секунд или минут. В подкорковых центрах мозга - десятки минут, часы и даже дни. В коре больших полушарий - до нескольких десятков лет.
 Следовые
процессы имеют важное значение в
понимании механизмов памяти.
Непродолжительное последействие до 1
часа связано с циркуляцией импульсов
в нервных цепях (Р. Лоренте де Но, 1934) и
обеспечиваеткратковременную
память.
Механизмы долговременной
памяти
основаны на изменении структуры белков.
В процессе запоминания, согласно
биохимической теории памяти (Х. Хиден,
1969) происходят структурные изменения
в молекулах РНК, на основе которых
строятся измененные белки с отпечатками
прежних раздражителей. Эти белки
длительно содержатся в нейронах, а также
в глиальных клетках головного мозга.
Следовые
процессы имеют важное значение в
понимании механизмов памяти.
Непродолжительное последействие до 1
часа связано с циркуляцией импульсов
в нервных цепях (Р. Лоренте де Но, 1934) и
обеспечиваеткратковременную
память.
Механизмы долговременной
памяти
основаны на изменении структуры белков.
В процессе запоминания, согласно
биохимической теории памяти (Х. Хиден,
1969) происходят структурные изменения
в молекулах РНК, на основе которых
строятся измененные белки с отпечатками
прежних раздражителей. Эти белки
длительно содержатся в нейронах, а также
в глиальных клетках головного мозга.
Утомление нервных центров возникает достаточно быстро при длительно повторных раздражениях. Быстрая утомляемость нервных центров объясняется постепенным истощением в синапсах запасов медиаторов, снижением чувствительности к ним постсинаптической мембраны, ее белков-рецепторов, снижением энергоресурсов клеток. В результате рефлекторные реакции начинают ослабевать, а затем полностью прекращаются.
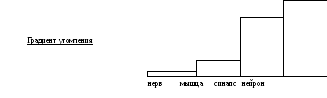
Разные нервные центры имеют различную скорость утомления. Менее утомляемы центры ВНС, координирующие работу внутренних органов. Значительно более утомляемы центры СНС, управляющие произвольной скелетной мускулатурой.
Тонус нервных центров определяется тем, что в состоянии покоя часть его нервных клеток находятся в возбуждении. Импульсы обратной афферентации от рецепторов исполнительных органов постоянно идут к нервным центрам, поддерживая в них тонус. В ответ на информацию с периферии центры посылают редкие импульсы к органам, поддерживая в них соответствующий тонус. Даже во время сна мышцы не расслабляются полностью и контролируются соответствующими центрами.
Влияние химических веществ на работу нервных центров определяется химическим составом крови и тканевой жидкости. Нервные центры очень чувствительны к дефициту кислорода и глюкозы. Клетки коры мозга погибают уже через 5-6 минут, клети ствола мозга выдерживают 15-20 минут, а клетки спинного мозга восстанавливают свои функции даже через 30 минут после полного прекращения кровоснабжения.
Существуют химические вещества избирательного действия. Стрихнин возбуждает нервные центры, блокируя работу тормозных синапсов. Хлороформ и эфир сначала возбуждают, а затем подавляют работу нервных центров. Апоморфин возбуждает рвотный центр, цититон и лобелин - дыхательный центр, а морфин угнетает его работу. Коразол возбуждает клетки двигательной зоны коры, вызывая эпилептические судороги.
Вывод. Функциональные возможности и свойства нервных центров зависят от состояния внутренних механизмов и влияния внешних факторов, действующих на организм.
Лекция № 5. Торможение в ЦНС. Принципы координации в деятельности ЦНС. Специфические и неспецифические структуры ЦНС, их значение.
"Нервная деятельность вообще состоит из явлений раздражения и торможения. Это и есть как бы две половины одной нервной деятельности".
И.П. Павлов
Явление торможения в нервных центрах было впервые открыто И.М. Сеченовым в 1862 г. Значение этого процесса было рассмотрено в его книге "Рефлексы головного мозга".
Торможение - это активный процесс в нервной системе, который вызывается возбуждением и проявляется как подавление другого возбуждения.
Торможение играет важную роль в координации движений, регуляции вегетативных функций, в реализации процессов высшей нервной деятельности. Тормозные процессы:
1 - ограничивают иррадиацию возбуждения и концентрируют его в определенных отделах НС;
2 - выключают деятельность ненужных в данный момент органов, согласовывает их работу;
3 - предохраняют нервные центры от перенапряжения в работе.
По месту возникновения торможение бывает:
1 - пресинаптическое;
2 - постсинаптическое.
По форме торможение может быть:
1 - первичным;
2 - вторичным.
Для возникновения первичного торможения в НС существуют специальные тормозные структуры (тормозные нейроны и тормозные синапсы). В этом случае торможение возникает первично, т.е. без предшествующего возбуждения.
 Пресинаптическое
торможение
возникает перед синапсом в аксональных
контактах. В основе такого торможения
лежит развитие длительной деполяризации
терминали аксона и блокирование
проведения возбуждения к следующему
нейрону.
Пресинаптическое
торможение
возникает перед синапсом в аксональных
контактах. В основе такого торможения
лежит развитие длительной деполяризации
терминали аксона и блокирование
проведения возбуждения к следующему
нейрону.
 Постсинаптическое
торможение
связано с гиперполяризацией
постсинаптической мембраны под влиянием
тормозных медиаторов типа гамма-аминомасляной
кислоты (ГАМК). Тормозные медиаторы
выделяются специальными тормозными
нейронами - клетками
Реншоу (в
спинном мозге) и корзинчатыми
клетками(в
промежуточном мозге).
Постсинаптическое
торможение
связано с гиперполяризацией
постсинаптической мембраны под влиянием
тормозных медиаторов типа гамма-аминомасляной
кислоты (ГАМК). Тормозные медиаторы
выделяются специальными тормозными
нейронами - клетками
Реншоу (в
спинном мозге) и корзинчатыми
клетками(в
промежуточном мозге).
Клетки Реншоу обеспечивают развитие торможения в мотонейронах мышц - антагонистов. Они также обеспечивают возвратное (антидромное) торможение, предохраняя мотонейроны от перевозбуждения.
 Корзинчатые
клетки
регулируют потоки импульсов возбуждения,
идущие к центрам промежуточного мозга
и коре полушарий. Они вызывают синхронное
торможение целой группы нейронов
диэнцефальных центров, регулируя таким
образом ритм активности коры.
Корзинчатые
клетки
регулируют потоки импульсов возбуждения,
идущие к центрам промежуточного мозга
и коре полушарий. Они вызывают синхронное
торможение целой группы нейронов
диэнцефальных центров, регулируя таким
образом ритм активности коры.
Для возникновения вторичного торможения не требуется специальных тормозных структур. Оно возникает в результате изменения функциональной активности обычных возбудимых нейронов. Вторичное торможение иначе называется пессимальным. При высокой частоте импульсов постсинаптическая мембрана сильно деполяризуется и становится неспособной отвечать на импульсы, идущие к клетке.
В живом организме работа всех органов является согласованной.
Согласование отдельных рефлексов для выполнения целостных физиологических актов называется координацией.
За счет координированной работы нервных центров осуществляется управление двигательными актами (бег, ходьба, сложные целенаправленные движения практической деятельности), а также изменение режима работы органов дыхания, пищеварения, кровообращения, т.е. вегетативных функций. Этими процессами достигается приспособление организма к изменениям условий существования.
Координация основывается на ряде общих закономерностей (принципов):
1. Принцип конвергенции (установлен Шеррингтоном) - к одному нейрону поступают импульсы из разных отделов нервной системы. Например, к одному и тому же нейрону могут конвергировать импульсы от слуховых, зрительных, кожных рецепторов.
2. Принцип иррадиации. Возбуждение или торможение, возникнув в одном нервном центре может распространяться на соседние центры.
 3.Принцип
реципрокности
(сопряженности; согласованного
антогонизма) был изучен Сеченовым,
Введенским, Шеррингтоном. При возбуждении
одних нервных центров деятельность
других центров может тормозиться. У
спинальных животных раздражение одной
конечности одновременно вызывает ее
сгибание, а на другой стороне одновременно
наблюдается разгибательный рефлекс.
3.Принцип
реципрокности
(сопряженности; согласованного
антогонизма) был изучен Сеченовым,
Введенским, Шеррингтоном. При возбуждении
одних нервных центров деятельность
других центров может тормозиться. У
спинальных животных раздражение одной
конечности одновременно вызывает ее
сгибание, а на другой стороне одновременно
наблюдается разгибательный рефлекс.
Реципрокность иннервации обеспечивает согласованную работу групп мышц при ходьбе, беге. При необходимости взаимосочетанные движения могут изменяться под контролем головного мозга. Например, при прыжках происходит сокращение одноименных групп мышц обеих конечностей.
 4.Принцип
общего конечного пути
связан с особенностью строения ЦНС.
Дело в том, что афферентных нейронов в
несколько раз больше, чем эфферентных,
поэтому множество афферентных импульсов
стекаются к общим для них эфферентным
путям. Система реагирующих нейронов
образует как бы воронку ("воронка
Шеррингтона"), поэтому множество
разных раздражений может вызвать одну
и ту же двигательную реакцию. Шеррингтон
предложил различать:
4.Принцип
общего конечного пути
связан с особенностью строения ЦНС.
Дело в том, что афферентных нейронов в
несколько раз больше, чем эфферентных,
поэтому множество афферентных импульсов
стекаются к общим для них эфферентным
путям. Система реагирующих нейронов
образует как бы воронку ("воронка
Шеррингтона"), поэтому множество
разных раздражений может вызвать одну
и ту же двигательную реакцию. Шеррингтон
предложил различать:
а) союзные рефлексы (которые усиливают друг друга, встречаясь на общих конечных путях);
б) антагонистические рефлексы (которые тормозят друг друга).
Преобладание на конечных путях той или иной рефлекторной реакции обусловлено ее значением для организма в данный момент. В таком отборе важную роль играет доминанта, обеспечивающая протекание главной реакции.
5. Приницп доминанты (установлен Ухтомским).
Доминанта (лат. dominans - господстввующий) - это господствующий очаг возбуждения в ЦНС, определяющий характер ответной реакции организма на раздражение.
Для доминанты характерно устойчивое перевозбуждение нервных центров, способность к суммации посторонних раздражителей и инертность (сохранность после действия раздражения). Доминантный очаг притягивает к себе импульсы из других нервных центров и за счет них усиливается. Как фактор поведения доминанта связана с высшей нервной деятельностью, с психологией человека. Доминанта является физиологической основой акта внимания. Формирование и торможение условных рефлексов так же связано с доминантным очагом возбуждения.
В нервной системе по современным представлениям, имеются специфические и неспецифические структуры.
Специфические структуры ЦНС лежат в наружных и боковых ее отделах, а неспецифические структуры - в срединных отделах. Они отличаются по строению и функции.
К специфическим структурам относятся все нервные центры и пути, проводящие афферентные нервные импульсы от рецепторов тела (восходящие пути) и эфферентные импульсы к рабочим органам (нисходящие пути).
Восходящие пути проводят сигналы мышечно-суставной, тактильной, слуховой, зрительной, болевой и температурной чувствительности к нервным центрам.
Нисходящие пути проводят импульсы, обеспечивающие рефлекторные реакции мышц и желез (исполнительных органов).
Деятельность специфических структур ЦНС связана с анализом раздражителей и определенным характером ответных реакций организма. В этой деятельности принимают участие и неспецифические структуры, изменяющие восприятие специфических раздражений и эфферентную деятельность органов и систем.
Неспецифические структуры не связаны с анализом какой-либо специфической чувствительности или с выполнением конкретных рефлекторных актов, но играют важную роль в процессах интеграции функций организма. По расположению нейронов и обилию их связей неспецифические структуры называются ретикулярной формацией (РФ). Она оказывает активизирующее или тормозящее влияние на работу других нервных центров. Это влияние на вышележащие центры называется восходящим, а на нижележащие центры - нисходящим.
Лекция № 6. Физиология спинного мозга. Рецепторные поля спинного мозга. Виды передаваемой информации. Основные центры спинного мозга. Рефлексы спинного мозга. Рефлекторные дуги простых и сложных соматических рефлексов спинного мозга.
"Все бесконечное разнообразие внешних проявлений мозговой деятельности сводится к одному лишь явлению - мышечному движению".
И.М. Сеченов
Спинной мозг человека является самым древним и примитивным отделом ЦНС, сохраняющим у самых высокоорганизованных животных свою морфологическую и функциональную сегментарность. В филогенезе отмечается снижение удельного веса спинного мозга по отношению к общей массе ЦНС. Если у примитивных позвоночных удельный вес спинного мозга составляет почти 50%, то у человека удельный вес его равен 2%. Это объясняется прогрессивным развитием полушарий мозга, цефализацией и кортикализацией функций. В филогенезе также наблюдается стабилизация количества сегментов спинного мозга.
Надежность сегментарных функций спинного мозга обеспечивается многочисленностью его связей с периферией. Первая особенность сегментарной иннервации состоит в том, что каждый сегмент спинного мозга иннервирует 3 метамера (сегмента тела) - собственный, половину вышележащего и половину нижележащего сегмента. Получается, что каждый метамер получает иннервацию от трех сегментов спинного мозга. Это гарантирует выполнение спинным мозгом его функций при повреждении мозга и его корешков. Вторая особенность сегментарной иннервации заключается в избытке чувствительных волокон в составе задних корешков спинного мозга по сравнению с количеством двигательных волокон передних корешков ("воронка Шеррингтона") у человека в соотношении 5:1. При большом разнообразии поступающей информации с периферии организм использует небольшое количество исполнительных структур для ответной реакции.
Общее число афферентных волокон у человека достигает 1 млн. Они несут импульсы от рецепторных полей:
1 - кожи шеи, туловища конечностей;
2 - мышц шеи, туловища и конечностей;
3 - внутренних органов.
Самые толстые миелиновые волокна идут от рецепторов мышц и сухожилий. Средней толщины волокна идут от тактильных рецепторов кожи, части мышечных рецепторов и рецепторов внутренних органов. Тонкие миелиновые и безмиелиновые волокна отходят от болевых и температурных рецепторов.
Общее число эфферентных волокон у человека около 200 тыс. Они несут импульсы от ЦНС к исполнительным органам (мышцам и железам). мышцы шеи, туловища, конечностей получают двигательную информацию, а внутренние органы получают вегетативную моторную и секреторную информацию.
Связь спинного мозга с периферией обеспечивается посредством корешков (задних и передних), в которых содержатся рассмотренные выше волокна. Задние корешки, по функции чувствительные, обеспечивают ввод информации в ЦНС. Передние корешки - двигательные и обеспечивают информационный выход из ЦНС.
Функции спинномозговых корешков были выяснены при помощи методов перерезки и раздражения. Белл и Мажанди установили, что при односторонней перерезке задних корешков отмечается утрата чувствительности, а двигательная функция при этом сохраняется. Перерезка передних корешков приводит к параличу конечностей соответствующей стороны, а чувствительность сохраняется полностью.
Мотонейроны спинного мозга возбуждаются за счет афферентных импульсов, идущих от рецепторных полей. Активность мотонейронов зависит не только от потоков афферентной информации, но и от сложных внутрицентральных взаимоотношений. Важную роль здесь играют нисходящие влияния коры полушарий, подкорковых ядер и ретикулярной формации, которые корректируют спинальные рефлекторные реакции. Также большое значение имеют многочисленные контакты вставочных нейронов, среди которых особая роль принадлежит тормозным клеткам Реншоу. Образуя тормозные синапсы, они контролируют работу мотонейронов и предупреждают их перевозбуждение. В работу нейронов вмешиваются также потоки импульсов обратной афферентации, идущие от проприорецепторов мышц.
В сером веществе спинного мозга содержится около 13,5 млн. нейронов. Из них мотонейроны составляют только 3%, а остальные 97% - это вставочные нейроны. Среди спинномозговых нейронов различают:
1 - крупные -мотонейроны;
2 - мелкие -мотонейроны.
От первых идут толстые быстропроводящие волокна к скелетным мышцам и вызывают двигательные акты. От вторых отходят тонкие нескоростные волокна к проприорецепторам мышц (веретенам Гольджи) и повышают чувствительность мышечных рецепторов, информирующих мозг о выполнении этих движений.
Группа -мотонейронов, которая иннервирует отдельную скелетную мышцу называется моторным ядром.
Вставочные нейроны спинного мозга за счет богатства синаптических связей обеспечивает собственную интегративную деятельность спинного мозга, включая управление сложными двигательными актами.
Ядра спинного мозга в функциональном отношении являются рефлекторными центрами спинальных рефлексов.
В шейном отделе спинного мозга находится центр диафрагмального нерва, центр сужения зрачка. В шейном и грудном отделах имеются моторные центры мышц верхних конечностей, груди, живота и спины. В поясничном отделе есть центры мышц нижних конечностей. В крестцовом отделе располагаются центры мочеиспускания, дефекации и половой деятельности. В боковых рогах грудного и поясничного отделов лежат центры потоотделения и сосудодвигательные центры.
Рефлекторные дуги отдельных рефлексов замкнуты через определенные сегменты спинного мозга. Наблюдая нарушение деятельности тех или иных групп мышц, тех или других функций, можно установить, какой отдел или сегмент спинного мозга поражен или поврежден.
Спинальные рефлексы можно изучить в чистом виде после разделения спинного и головного мозга. Спинальные лабораторные животные сразу после перерезки впадают в состояние спинального шока, который длится несколько минут (у лягушки), несколько часов (у собаки), несколько недель (у обезьяны), а у человека продолжается месяцами. У низших позвоночных (лягушка) спинальные рефлексы обеспечивают сохранение позы, движений, защитных, половых и других реакций. У высших позвоночных без участия центров головного мозга и РФ спинной мозг не способен полноценно выполнять эти функции. Спинальная кошка или собака не может сама стоять и ходить. У них наблюдается резкое падение возбудимости и угнетение функций центров, лежащих ниже места перерезки. Такова цена цефализации функций, подчиненности спинальных рефлексов центрам головного мозга. После выхода из спинального шока постепенно восстанавливаются рефлексы скелетных мышц, регуляция АД, мочеиспускания, дефекации, ряд половых рефлексов. Не восстанавливаются произвольные движения, чувствительность, температура тела и дыхание - их центры лежат выше спинного мозга и при перерезке изолируются. Спинальные животные могут жить только в условиях ИВЛ (искусственной вентиляции легких).
Изучая свойства рефлексов у спинальных животных Шеррингтон в 1906 году установил закономерности рефлекторной деятельности и выделил основные виды спинальных рефлексов:
1 - защитные (оборонительные) рефлексы;
2 - рефлексы на растяжение мышц (миотатические);
3 - межсегментарные рефлексы координации движений;
4 - вегетативные рефлексы.
Несмотря на функциональную зависимость спинномозговых центров от головного мозга, многие спинальные рефлексы протекают автономно, мало подчиняясь управлению сознанием. Например, сухожильные рефлексы, которые используются в медицинской диагностике:
|
Название рефлекса |
Раздражение |
Реакция |
Участвующие сегменты сп. мозга |
|
Локтевой
Брюшной кожный Кремастерный
Коленный
Ахиллов
Подошвенный
Анальный |
Удар молоточком по сухожилию бицепса Штриховое раздражение кожи живота Штриховое раздражение внутренней поверхности бедра Удар по сухожилию четырехглавой м. бедра Удар по ахиллову сухожилию Штриховое раздражение подошвы Штрих или укол вблизи заднего прохода |
Сгибание руки
Сокращение мышц живота Поднятие яичка
Разгибание голени
Сгибание стопы
Сгибание пальцев стопы Сокращение сфинктера пр. кишки |
С5 - С6
Т7 - Т12
L1 - L2
L2 - L4
S1 - S2
S1 - S2
S4 - S5 |
Все эти рефлексы имеют простую двухнейронную (гомонимную) рефлекторную дугу.
 Кожно-мышечные
рефлексы
имеют трехнейронную (гетеронимную)
рефлекторную дугу.
Кожно-мышечные
рефлексы
имеют трехнейронную (гетеронимную)
рефлекторную дугу.
Вывод: спинной мозг имеет важное функциональное значение. Выполняя проводниковую и рефлекторную функции, он является необходимым звеном нервной системы в осуществлении координации сложных движений (передвижение человека, его трудовая деятельность) и вегетативных функций.
Лекция № 7. Физиология ромбовидного мозга. Основные рефлекторные центры продолговатого мозга и моста. Бульбарные рефлексы.
Непосредственным продолжением вверх спинного мозга у всех позвоночных животных и человека является продолговатый мозг. Вместе с варолиевым мостом продолговатый мозг является производным ромбовидного мозга (rhombencephalon). Одновременно он является частью ствола мозга (truncus encephalicus). Ствол - это осевые структуры головного мозга, включающие продолговатый мозг, мост, средний и промежуточный мозг.
 В
продолговатом мозге и мосту по сравнению
со спинным мозгомнет
четкой сегментации
серого и белого вещества. Скопления
нейронов здесь формируют ядра,
обеспечивающие сложную рефлекторную
деятельность. У низших позвоночных
(рептилий, птиц) варолиев мост не отделен
от продолговатого мозга, так как у них
эти отделы состоят только из покрышки
ствола.
В
продолговатом мозге и мосту по сравнению
со спинным мозгомнет
четкой сегментации
серого и белого вещества. Скопления
нейронов здесь формируют ядра,
обеспечивающие сложную рефлекторную
деятельность. У низших позвоночных
(рептилий, птиц) варолиев мост не отделен
от продолговатого мозга, так как у них
эти отделы состоят только из покрышки
ствола.
У высших позвоночных полость ромбовидного мозга (IV желудочек) также не разделяется и является общей, а дно желудочка - ромбовидная ямка (fossa rhomboidea) не имеет четких границ, которые бы разделяли продолговатый мозг и мост. При формировании структур ромбовидного мозга двигательные и чувствительные ядра образуются из основной и крыльной пластинок нервной трубки. В пределах разделяющих их боковые бороздок формируются вегетативные ядра. Середина продолговатого мозга и моста заняты задним отделом ретикулярной формации (РФ), которая оказывает неспецифические возбуждающие и тормозящие влияния на головной и спинной мозг.
Только с вентральной стороны у высших позвоночных наблюдается обособление моста с развитием мощных нисходящих путей от коры к мосту и спинному мозгу. Здесь же, в вентральной части, идет увеличение собственных ядер моста, где заканчиваются мощные корково-мостовые пути и коллатерали (боковые ответвления) от двигательных пирамидных путей. Ядра моста контролируют быстрый сон.
 В
продолговатом мозге и варолиевом мосту
(в дорзальной их части) находится большая
группа ядер черепных нервов (сV
по XII
пары), которые образуют массу серого
вещества ромбовидной ямки. Эти ядра
являются жизненно важными функциональными
центрами. На дне IV
желудочка в продолговатом мозге находится
дыхательный
центр,
состоящий из центра вдоха и центра
выдоха. Нейроны этих центров посылают
импульсы к дыхательным мышцам через
мотонейроны спинного мозга, контролируя
правильное чередование фаз дыхательного
цикла. Рядом лежит сосудодвигательный
центр
(контролирует тонус стенок сосудов и
уровень АД) и сердечно-сосудистый
центр
(согласует состояние сосудов с работой
сердца). Функции этих центров тесно
связаны. Импульсы из дыхательного центра
изменяют частоту сердечных сокращений,
вызывая физиологическую
дыхательную аритмию -
учащение сердцебиение на вдохе и
замедление ударов сердца на выдохе.
В
продолговатом мозге и варолиевом мосту
(в дорзальной их части) находится большая
группа ядер черепных нервов (сV
по XII
пары), которые образуют массу серого
вещества ромбовидной ямки. Эти ядра
являются жизненно важными функциональными
центрами. На дне IV
желудочка в продолговатом мозге находится
дыхательный
центр,
состоящий из центра вдоха и центра
выдоха. Нейроны этих центров посылают
импульсы к дыхательным мышцам через
мотонейроны спинного мозга, контролируя
правильное чередование фаз дыхательного
цикла. Рядом лежит сосудодвигательный
центр
(контролирует тонус стенок сосудов и
уровень АД) и сердечно-сосудистый
центр
(согласует состояние сосудов с работой
сердца). Функции этих центров тесно
связаны. Импульсы из дыхательного центра
изменяют частоту сердечных сокращений,
вызывая физиологическую
дыхательную аритмию -
учащение сердцебиение на вдохе и
замедление ударов сердца на выдохе.
В продолговатом мозге также лежат пищеварительные центры:
1 - моторные (жевания, глотания, моторики желудка и части кишечника),
2 - секреторные (слюноотделения, желудочной секреции, выделения соков тонкой кишки, поджелудочной железы и др.).
Здесь же находятся центры защитных рефлексов (чихания, кашля, рвоты, мигания, слезоотделения). Таким образом, биологическая роль продолговатого мозга заключается в регуляции постоянства состава внутренней среды организма (гомеостаза). Он также осуществляет более тонкие приспособительные реакции организма к внешней среде, чем спинной мозг.
Кроме ядер черепных нервов в продолговатом мозге имеются переключательные чувствительные ядра. В толще олив лежат крупные нижние оливные ядра, а также медиальные и верхние добавочные оливные ядра. На задней поверхности в одноименных бугорках залегают тонкое ядро Голля и клиновидное ядро Бурдаха.
Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса мышц. Импульсы от вестибулярных ядер (Дейтерса и Бехтерева) усиливают тонус мышц разгибателей, что необходимо для организации позы. Неспецифические отделы продолговатого мозга (ядра РФ) наоборот снижают тонус мышц, в том числе и разгибателей. Продолговатый мозг участвует в осуществлении рефлексов поддержания и восстановления позы тела (установочных рефлексов).
Продолговатый мозг выполняет две функции (рефлекторную и проводниковую). Рефлекторная функция осуществляется за счет:
1 - простых сегментарных рефлексов (защитный мигательный, слезоотделения, движений ушной раковины, кашля, чихания, рвоты),
2 - лабиринтных рефлексов (распределение тонуса между отдельными группами мышц и установки определенной позы),
3 - установочных рефлексов (поддержания позы и рабочих движений),
4 - вегетативных рефлексов (дыхания, кровообращения, пищеварения).
Проводниковая функция осуществляется путем проведения:
1 - восходящих волокон от спинного мозга к коре полушарий,
2 - нисходящих волокон от коры полушарий к спинному мозгу,
3 - собственных проводящих пучков продолговатого мозга и моста, соединяющих ядро и оливу вестибулярного нерва с мотонейронами спинного мозга.
Продолговатый мозг принимает чувствительные волокна от рецепторов мимических и жевательных мышц, мышц шеи, конечностей и туловища, от кожи лица, слизистых оболочек глаз, полости носа и рта, от рецепторов органов слуха и равновесия, от рецепторов гортани, трахеи, легких, интерорецепторов ЖКТ и сердечно-сосудистой системы. Здесь волокна переключаются на другие нейроны, образуя путь в таламус и кору полушарий. Восходящие пути кожно-мышечной чувствительности перекрещиваются на уровне продолговатого мозга так же, как и большая часть пирамидных (двигательных) путей.
Функции продолговатого мозга были изучены на бульбарных животных, у которых поперечным разрезом продолговатый мозг отделен от среднего мозга. Следовательно, жизнь бульбарных животных осуществляется за счет деятельности спинного и продолговатого мозга. У таких животных отсутствуют произвольные движения, отмечается потеря всех видов чувствительности, нарушается регуляция температуры тела (теплокровное животное превращается в холоднокровное). У бульбарных животных сохраняются рефлекторные реакции организма и осуществляется регуляция функций внутренних органов.
У человека бульбарные нарушения проявляются нарушением глотания и фонации. В тяжелых случаях больные совершенно не могут глотать и говорить (признаки поражения ядер IX, X, XII пар черепных нервов). При поражении ядер V, VII и IX пар черепных нервов появляются, расстройства жевания, мимики, движения головой. Разрушение центров ромбовидного мозга приводит к мгновенной смерти из-за остановки сердца и остановки дыхания, паралича сосудов и падения АД (сосудистый коллапс).
Гипоксия - состояние, возникающее при дефиците снабжения тканей кислородом или при нарушении его использования клетками тканей.
Для нормальной жизнедеятельности клеткам необходим постоянный приток кислорода. Он расходуется главным образом в процессе синтеза АТФ - источника клеточной энергии. При нарушении биологического окисления возникает дефицит АТФ, т.е. энергетическое голодание, составляющее основу гипоксии.
Разные органы и ткани имеют неодинаковую чувствительность к гипоксии. Наиболее чувствительной является ткань мозга. При массе мозга, составляющей 2% от массы тела, он поглощает 20% всего потребляемого кислорода. Кислородное голодание организма есть кислородное голодание мозга.
Наиболее ранние функциональные расстройства при гипоксии возникают в сфере высшей нервной деятельности. Первоначально наблюдается общее возбуждение и ослабляется внимание. Изменяется почерк и ослабевает память, возрастает число ошибок в решении сложных задач. Затем, возникают сонливость и безразличие к внешнему миру, теряется ориентация во времени и пространстве. Нарушаются движения и снижается болевая чувствительность. При дальнейшем нарастании гипоксии возможна потеря сознания, судороги, паралич и смерть.
Асфиксия (удушение) существенно отличается от острой гипоксии. Быстро развивается расстройство дыхания и кровообращения. Наступает состояние комы - глубокого угнетения функций ЦНС: полная потеря сознания, утрата всех рефлекторных реакций на внешние раздражители, глубокое расстройство регуляции жизненно важных функций организма. Гибель корковых нейронов наступает уже через 3-4 минуты.
Лекция № 8. Средний мозг и мозжечок, их функциональное значение. Виды передаваемой информации. Основные центры. Рефлексы среднего мозга и мозжечка.
Мозжечок (cerebellum - дословно, малый мозг) - структура ромбовидного мозга. В онтогенезе он образуется из дорзальной стенки ромбовидного мозгового пузыря. У круглоротых из ядер нервов боковой линии и вестибулярных ядер продолговатого мозга, которые координируют движения и регулируют тонус мускулатуры, формируется примитивный мозжечок. Они имеет вид пластинки с наружным волокнистым слоем и внутренним клеточным. У рыб с усложнением моторных функций развиваются боковые ушки мозжечка (auriculae cerebelli lateralis) и особенно его средняя часть - заслонка (valvula)). У амфибий, выходящих на сушу, с начальным упрощением движений мозжечок редуцируется, заслонка полностью исчезает. У рептилий, с освоением ходьбы и бега, формируются структуры древнего мозжечка (paleocerebellum) и появляются ядра мозжечка, развиваются связи мозжечка с другими локомоторными структурами. У птиц тонкая координация движений в полете явилась причиной развития связей центральных структур мозжечка. Задние его отделы (клочок) стали основой старого мозжечка (archicerebellum), средние - устанавливают связи со зрительными центрами среднего мозга, а передние - со спинным мозгом. Так, в мозжечковые рефлексы координации у птиц включается зрение. У млекопитающих появляется филогенетически молодая часть плащ мозжечка (pallium cerebelli). Мозжечок связан со всеми отделами нервной системы, которые контролируют движения. Развитие идет по пути совершенствования связей мозжечка со спинным мозгом и ослабления связей с вестибулярной системой. У высших млекопитающих мозжечок состоит из трех частей:
1 - древний мозжечок (его каудальные отделы) контролирует вестибулярную функцию,
2 - старый мозжечок (его передняя доля) обеспечивает связь со спинным мозгом,
3 - новый мозжечок (его средняя доля и полушария) связаны с корой большого мозга.
 У
позвоночных в мозжечке появляются ядра
(зубчатое, пробковидное, шаровидное и
ядро шатра). Через зубчатое ядро мозжечок
посылает восходящие импульсы к красному
ядру, таламусу и лобным долям коры
больших полушарий. Через ядро шатра
мозжечок проводит импульсы к ретикулярной
формации и вестибулярным ядрам, а по
вестибуло-спинальному пути - к мотонейронам
спинного мозга.
У
позвоночных в мозжечке появляются ядра
(зубчатое, пробковидное, шаровидное и
ядро шатра). Через зубчатое ядро мозжечок
посылает восходящие импульсы к красному
ядру, таламусу и лобным долям коры
больших полушарий. Через ядро шатра
мозжечок проводит импульсы к ретикулярной
формации и вестибулярным ядрам, а по
вестибуло-спинальному пути - к мотонейронам
спинного мозга.
Связи мозжечка с различными отделами ЦНС обеспечиваются за счет трех пар мозжечковых ножек. Нижние ножки соединяют мозжечок со спинным и продолговатым мозгом, средние - с варолиевым мостом и двигательной зоной коры полушарий мозга, верхние со средним мозгом и таламусом.
В кору мозжечка импульсы поступают по двум видам афферентных волокон:
1 - лазающие волокна заканчиваются синапсами на клетках Пуркинье и несут к ним возбуждающие импульсы,
2 - моховидные волокна через зернистые клетки обеспечивают возбуждение, а через корзинчатые клетки - торможение клеток Пуркинье. Клетки Гольджи способны тормозить возбуждение зернистых клеток, освобождая таким образом клетки Пуркинье от возбуждающих влияний.
Функции мозжечка были изучены итальянским физиком Луиджи Лючиани в 1893 году на животных с частично или полностью удаленным мозжечком (мозжечковые животные), а также путем регистрации его биоэлектрической активности в покое и при раздражении.
 При
удалении половины мозжечка
повышается тонус мышц разгибателей,
конечности животного вытягиваются,
изгибается туловище и отклоняется
голова в оперированную сторону.
Наблюдается движение по кругу в
оперированную сторону - "манежные
движения". Постепенно описанные
нарушения сглаживаются, но сохраняется
некоторая дискоординация движений.
При
удалении половины мозжечка
повышается тонус мышц разгибателей,
конечности животного вытягиваются,
изгибается туловище и отклоняется
голова в оперированную сторону.
Наблюдается движение по кругу в
оперированную сторону - "манежные
движения". Постепенно описанные
нарушения сглаживаются, но сохраняется
некоторая дискоординация движений.
При удалении всего мозжечка появляются выраженные двигательные расстройства. Эти расстройства постепенно сглаживаются за счет компенсаторной активизации двигательной зоны коры мозга, но животное все же остается двигательным инвалидом с нарушением координации движений. Двигательные расстройства у мозжечкового животного называются триадой Лючиани:
1 - амония (потеря тонуса мышц и его неправильное распределение),
2 - астения (ослабление мышечных сокращений и выраженное мышечное утомление),
3 - астазия (неустойчивость при стоянии, качательные движения головы и тела, их дрожание).
В результате у животного происходит дискоординация движений (шаткая походка, размашистые, неловкие, неточные движения). Такой комплекс двигательных расстройств поучил общее название мозжечковой атаксии.
Академик Орбели в 1938 году установил, что мозжечок влияет на состояние рецепторного аппарата, а также на вегетативные процессы и состояние гладкой мускулатуры внутренних органов. Наступающие под влиянием мозжечка изменения кровообращения, состава крови, пищеварения, дыхания направлены на трофическое обеспечение деятельности скелетных мышц. Орбели рассматривал мозжечок не только как помощника коры мозга в регуляции тонуса и мышечных движений, но и как высший адаптационно-трофический центр (греч. trophe - питание).
Мозжечок оказывает адаптационно-трофическое влияние на все отделы мозга через симпатический отдел нервной системы, регулируя в мозге обмен веществ и способствуя приспособлению ЦНС к изменяющимся условиям среды. Деятельность мозжечка непосредственно связана с корой больших полушарий и осуществляется под ее контролем.
 В
коре мозжечка представительство
различных периферических рецепторов
имеетсоматотопическую
организацию,
т.е. голова, туловище и конечности
проецируются на определенные участки
червя и полушарий мозжечка (см. схему).
Эти участки коры мозжечка связаны с
соответствующими чувствительными
зонами коры больших полушарий мозга.
Например, зрительная зона мозжечка
связана со зрительной зоной коры мозга,
а представительство каждой группы мышц
в мозжечке - с представительством этих
же мышц в коре мозга.
В
коре мозжечка представительство
различных периферических рецепторов
имеетсоматотопическую
организацию,
т.е. голова, туловище и конечности
проецируются на определенные участки
червя и полушарий мозжечка (см. схему).
Эти участки коры мозжечка связаны с
соответствующими чувствительными
зонами коры больших полушарий мозга.
Например, зрительная зона мозжечка
связана со зрительной зоной коры мозга,
а представительство каждой группы мышц
в мозжечке - с представительством этих
же мышц в коре мозга.
Такое соответствие облегчает совместную деятельность мозжечка и коры большого мозга в управлении различными функциями организма. В мозжечке происходит интеграция самых различных сенсорных влияний (проприоцептивных, вестибулярных и др.)
Вывод: Мозжечок выполняет три важнейших функции:
1 - координация движений,
2 - распределение мышечного тонуса и контроль равновесия,
3 - регуляция вегетативных процессов.
Средний мозг (mesencephalon) - верхний отдел ствола мозга, состоящий из ножек мозга и четверохолмия. В онтогенезе он образуется из среднего мозгового пузыря. Эволюция среднего мозга связана с возникновением и развитием зрения. У круглоротых впервые в крыше среднего мозга появляется зрительный центр (tectum) и формируются пути к центрам продолговатого мозга. У рыб развивается вентральная часть продолговатого мозга - покрышка (tegmentum), в которой формируются ядра черепных нервов (III, IV, VI), управляющих мышцами глазного яблока. Расширяются связи среднего мозга также и с продолговатым мозгом, его вестибулярными ядрами, ядрами боковой линии. Появляются пути к мозжечку. У рептилий образуется примитивное красное ядро (nucleus ruber), от которого нисходящие пути ведут в спинной мозг. У млекопитающих средний мозг устанавливает связи с таламусом, базальными ядрами и корой больших полушарий. Кроме красного ядра появляется черное вещество (substantia nigra), которое учавствует в регуляции движения. Тектум, который у птиц представляет собой двухолмие, превращается в четрверохолмие. Верхнее двухолмие остается зрительными центрами, а нижнее двухолмие формируется как слуховые центры. В центральной части среднего мозга располагается сетчатая формация (formatio reticularis) - неспецифическая структура ЦНС, изменяющая функциональное состояние вышележащих и нижележащих отделов мозга. В ножках мозга проходят восходящие и нисходящие проводящие пути.
В строении среднего мозга полностью утрачиваются сегментарные признаки. Его клеточные элементы образуют ядра, относящиеся непосредственно к среднему мозгу, а также ядра ретикулярной формации, контролирующие состояние бодрствования.
Верхние холмики четверохолмия получают импульсы от сетчатки глаз и регулируют величину зрачков, а также аккомодацию изменением кривизны хрусталика. Аккомодация - способность ясно видеть предметы на разных расстояниях от глаз. Являясь первичными зрительными центрами, верхние холмики обеспечивают сторожевые реакции с поворотом головы и глазных яблок к источнику внезапных зрительных раздражений.
Нижние холмики четверохолмия получают импульсы от ядер слуховых нервов, лежащих в продолговатом мозге. Являясь первичными слуховыми центрами, нижние холмики обеспечивают сторожевые реакции с поворотом головы, движением ушных раковин к источнику внезапных слуховых раздражений, натяжением барабанной перепонки и слуховых косточек.
 Красное
ядро является
переключательным центром связей мозжечка
с периферией, особенно с мышцами -
сгибателями (усиление тонуса). При
нарушении таких связей (перерезка между
верхними и нижними холмиками) наступает
резкое усиление тонуса мышц-разгибателей,
конечности вытягиваются, голова
откидывается назад, хвост поднимается.
Эффект такой перерезки называется
децеребрационной
ригидностью,
которая возникает как результат нарушения
тормозящего влияния красного ядра на
тонические рефлексы разгибания,
осуществляемые ядром
Дейтерса
(латеральное вестибулярное ядро), лежащим
в продолговатом мозге.
Красное
ядро является
переключательным центром связей мозжечка
с периферией, особенно с мышцами -
сгибателями (усиление тонуса). При
нарушении таких связей (перерезка между
верхними и нижними холмиками) наступает
резкое усиление тонуса мышц-разгибателей,
конечности вытягиваются, голова
откидывается назад, хвост поднимается.
Эффект такой перерезки называется
децеребрационной
ригидностью,
которая возникает как результат нарушения
тормозящего влияния красного ядра на
тонические рефлексы разгибания,
осуществляемые ядром
Дейтерса
(латеральное вестибулярное ядро), лежащим
в продолговатом мозге.
Черное вещество регулирует акты жевания, глотания, кровяное давление, дыхание, участвует в регуляции тонуса мышц (особенно при выполнении мелких движений пальцами рук) и поддержании позы. Гуморальная регуляция тонуса мышц обеспечивается черным веществом путем выделения его пигментными клетками вещества дофамина. Дефицит дофамина проявляется как болезнь Паркинсона (дрожательный паралич).
Вывод. Средний мозг регулирует тонус мышц, участвует в его распределении, что является необходимым условием для координированных движений. Средний мозг обеспечивает регуляцию ряда вегетативных функций организма (жевание, глотание, давление крови, дыхание). Средний мозг за счет сторожевых зрительных и слуховых рефлексов, усиления тонуса мышц-сгибателей подготавливает организм к ответу на внезапное раздражение. На уровне среднего мозга реализуются статические и статокинетические рефлексы.