

1. Физиология возбудимых тканей
В основе приспособительных реакций организма лежит раздражимость - способность реагировать на воздействия изменением структуры и функций. Раздражимостью обладают все клетки животных и растений. В ходе эволюции раздражимость некоторых тканей достигла наивысшего развития и трансформировалась в возбудимость (способность отвечать на раздражение возбуждением). К возбудимым относят нервную, мышечную и секреторную ткани. Возбудимость оценивают по порогу раздражения (минимальной силе раздражителя, которая способна вызвать возбуждение). Раздражители по их природе делят на физические, химические, биологические (вирусы, бактерии и др.), адекватные и неадекватные. Адекватными называют раздражители, к восприятию которых биологическая структура специально приспособлена. Поэтому пороговая сила адекватных раздражителей наименьшая. Например, адекватным для фоторецепторов является свет, для мышц - нервный импульс. Неадекватными называют раздражители, которые действуют на структуру, не приспособленную для их восприятия. Например, скелетная мышца реагирует сокращением и на электрические раздражения.
Биоэлектрические явления в возбудимых тканях. Возбуждение - это совокупность процессов, в результате которых кратковременная деполяризация цитоплазматической мембраны вызывает специализированную реакцию клетки (проведение нервного импульса, сокращение мышцы и т.д.).
Луиджи Гальвани обратил внимание на сокращение мышц препарата задних лапок, подвешенного на медном крючке, при соприкосновении с железными перилами балкона. На основании этого (первый опыт Гальвани) был сделан вывод, что сокращение вызвано «электричеством», которое передается по крючку и перилам от спинного мозга к мышцам. Однако физик А. Вольта предположил, что источником тока является не мозг, а потенциал в месте контакта разнородных металлов. В ответ на это Л. Гальвани стеклянным крючком набрасывал седалищный нерв на мышцы голени, что вызывало сокращение мышц (второй опыт или опыт без металлов) и доказывало существование «животного электричества». Позднее установлено, что клетки в покое внутри заряжены отрицательно по отношению к их поверхности. Этот потенциал покоя (ПП) составляет от 30 до 100 мВ.
В середине 20 века. А.Ходжкин, Э.Хаксли и Б.Катц создали мембранно-ионную теорию, согласно которой МП обусловлен разными концентрациями ионов калия, натрия и хлора по обе стороны клеточной мембраны. По сравнению с внеклеточной жидкостью, цитоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора. Высокая проницаемость мембраны для калия приводит к выходу части внутриклеточного калия в окружающую клетку среду и к появлению положительного заряда на наружной поверхности мембраны. Органические анионы, для которых мембрана непроницаема, создают на внутренней поверхности мембраны отрицательный заряд, а поддерживает высокие концентрации калия в клетке и натрия вне ее натрий-калиевый насос.
Раздражение возбудимой клетки может вызвать локальный ответ или потенциал действия. Локальный ответ возникает при подпороговых раздражениях. Он находится в прямой зависимости от силы стимула, локализуется на поверхности клетки только в месте ее раздражения и увеличивает возбудимость клетки. Потенциал действия (ПД) возникает под влиянием порогового или сверхпорогового раздражений. При этом проницаемость мембраны для натрия увеличивается и в результате проникновения натрия в клетку ее мембрана заряжается положительно по отношению к наружной среде. Затем закрываются натриевые и открываются дополнительные калиевые каналы. В результате выхода калия из клетки начинается восстанавление МП (реполяризация мембраны).
В ПД различают (рис. 1.):
1. Предспайк (локальный ответ) - деполяризация мембраны до критического уровня.
2. Спайк - состоит из восходящей (деполяризация) и нисходящей (реполяризация) частей.
3. Следовой потенциал - состоит из следовой деполяризации и гиперполяризации.
Возбудимость в период предспайка повышена (фаза повышенной возбудимости) и даже слабый дополнительный стимул может вызвать формирование ПД. В период спайка мембрана не возбудима (абсолютная рефрактерность). Затем возбудимость постепенно восстанавливается (относительная рефрактерность). В это время для нового возбуждения нужно сверхпороговое раздражение. При следовой деполяризации возбудимость повышена (экзальтация), а при гиперполяризации - понижена (субнормальная возбудимость).
Законы раздражения отражают зависимость ответной реакции возбудимой ткани от силы раздражителя.
Закон «все или ничего»: подпороговые раздражители не вызывают ответной реакции («ничего»), а пороговые раздражители вызывают максимальный ответ («все»). По этому закону сокращается одиночное мышечное волокно и сердце.
Закон силы: чем сильнее раздражение, тем больше ответная реакция. В соответствии с этим законом функционирует скелетная мышца. Она состоит из мышечных волокон с разной возбудимостью. На пороговые раздражители отвечают наиболее возбудимые волокна. Увеличение силы раздражителя дополнительно вовлекает в ответ волокна с меньшей возбудимостью и амплитуда сокращения мышцы растет.
|
|
|
Рис. 1. Соотношение одиночного цикла возбуждения (I) и фаз возбудимости (II) I: а - потенциал покоя; б – предспайк (локальный ответ); в - спайк; г - следовая деполяризация; д - следовая гиперполяризация. II: а - исходный уровень возбудимости; б - фаза повышенной возбудимости; в - фаза абсолютной и относительной рефрактерности; г - фаза экзальтации (супернормальная возбудимость); д - фаза субнормальной возбудимости. |

Закон раздражения Дюбуа-Реймона: действие постоянного тока зависит от его силы и скорости нарастания. При медленном нарастании ткань приспосабливается к раздражителю (аккомодация) и возбуждение может не возникать.
Закон силы-времени отражает зависимость пороговой величины постоянного тока от времени его действия. Чем короче импульсы тока, тем меньшую раздражающую способность они имеют. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение которого ток, равный реобазе, вызывает возбуждение, называется полезным временем. Хронаксия - минимальное время, в течение которого ток, равный двум реобазам, вызывает реакцию.
Закон полярного действия постоянного тока: при замыкании тока возбуждение возникает под катодом, а при размыкании - под анодом.
Закон физиологического электротона: в области катода возбудимость повышается (катэлектротон), а у анода - снижается (анэлектротон). При длительном действии постоянного тока возбудимость под катодом снижается (катодическая депрессия), а под анодом - растет (анодная экзальтация).
Нервное волокно обладает: возбудимостью, проводимостью и лабильностью. Возбуждения распространяется по нервному волокну только при его анатомической и физиологической целостности, не переходит на соседнее нервное волокно (закон изолированного проведения), не изменяется по амплитуде (закон незатухающего или бездекрементного проведения) и проводится в обе стороны от места раздражения (закон двустороннего проведения).
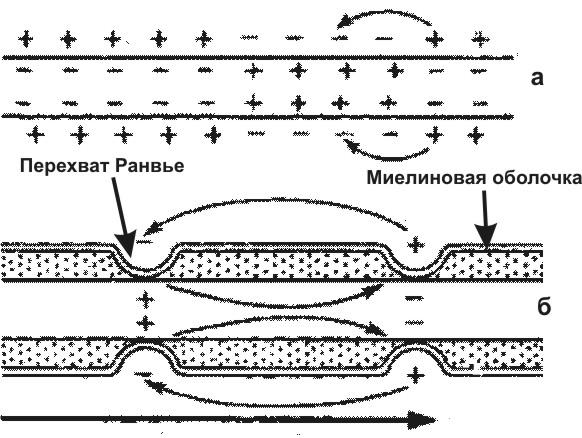
Возбужденный участок поверхности нервного волокна приобретает отрицательный заряд. Так как соседний невозбужденный участок заряжен положительно, то между ними потечет электрический ток. Это вызовет возбуждение покоящегося участка и тоже изменит его заряд. В конечном итоге возбуждение распространится по всей поверхности не покрытого миелиновой оболочкой (безмякотного) нервного волокна (рис. 2а.). В миелиновых (мякотных) отростках нейронов, возбуждение может возникать только в перехватах Ранвье. Поэтому оно распространяется скачками от одного перехвата к другому (рис. 2б.) и движется гораздо быстрее, чем в безмякотных волокнах.
|
|
|
Рис. 2. Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б) нервных волокнах |
По диаметру и скорости проведения возбуждения нервные волокна делят на типы А, В и С. Самые толстые волокна типа А (диаметр 12-22 мкм) с наибольшей скоростью (70-120 м/с) проводят возбуждение от мозга к скелетным мышцам и от рецепторов мышц к мозгу. От многих других рецепторов идут волокна типа А с несколько меньшими диаметром (от 8 до 1 мкм) и скоростью проведения возбуждения (5-70 м/с). К волокнам типа В относятся преганглионарные вегетативные волокна (диаметр - 1-3,5 мкм, скорость проведения возбуждения - 3-18 м/с). Только волокна типа С являются безмякотными (их диаметр 0,5-2 мкм, скорость проведения возбуждения менее 3 м/с). Они являются постганглионарными симпатическими волокнами, а также идут от болевых рецепторов, части терморецепторов и рецепторов давления.
Нервные волокна обладают лабильностью (функциональной подвижностью). Ее измеряют по максимальному количеству возбуждений, которое способно воспроизвести нервное волокно. У нервных волокон лабильность выше (до 1000 Гц) чем в других возбудимых структурах. Если нерв повредить (химическим веществом, нагреванием, охлаждением или током) не нарушая анатомическую целостность, то в нем развивается состояние парабиоза. При этом последовательно сменяются уравнительная, парадоксальная и тормозная фазы. В уравнительную фазу - при редких раздражениях все импульсы проводятся через поврежденный участок, а при высоком ритме только часть. В парадоксальную - ответная реакция на частые раздражения меньше, чем на редкие. В тормозную - нерв не проводит любые возбуждения. При выходе из парабиоза наблюдаются те же фазы, но в обратной последовательности.
Мышцы обладают возбудимостью (возбуждаются при действии раздражителей), проводимостью (проводить возбуждение) и сократимостью (способны изменять свою длину или напряжение при возбуждении). Для сердечных и части гладких мышечных волокон, дополнительно к перечисленным свойствам, характерна автоматия (способность к самопроизвольному возбуждению), а уникальным свойством гладких мышц является пластичность (долго сохраняют приданную им длину).
Сила мышцы определяется максимальным грузом, который она может поднять, а работа - произведением величины поднятого груза на высоту подъема. Максимальная работа производится при средних величинах нагрузок. При изотоническом сокращении мышцы изменяется ее длина, а напряжение постоянно (так сокращаются мышцы при отсутствии сопротивления изменению длины). При изометрическом сокращении длина мышцы постоянна, а ее напряжение растет (например, при попытке поднять чрезмерный груз). В естественных условиях наблюдаются смешанные сокращения (изменяются и длина и напряжение мышцы).
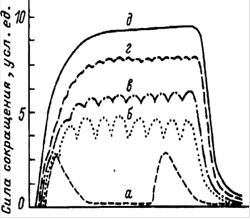
При однократном раздражении возникает одиночное мышечное сокращение. В нем выделяют: латентный период (время от начала раздражения до начала ответной реакции), фазу укорочения и фазу расслабления. Если каждый следующий стимул поступает к скелетной мышце в период ее укорочения (рис. 3) - возникает гладкий тетанус, а в фазу расслабления - зубчатый тетанус. В естественных условиях к скелетной мышце поступает такая серия импульсов, на которую мышца отвечает гладким тетанусом. Его амплитуда, как правило, выше амплитуды одиночного сокращения. Н.Е. Введенский объяснил это оптимумом и пессимумом. Оптимум - частота, при которой раздражения наносятся в фазу повышенной возбудимости (тетанус максимальный). Пессимум - частота, при которой новое раздражение наносится в фазу пониженной возбудимости (тетанус – минимален).
|
| |
|
Рис. 3. Виды суммации сокращений скелетной мышцы а - одиночные сокращения; б-г - зубчатые тетанусы; д - гладкий тетанус |
|
При изучении скелетных и сердечных мышц в поляризованном свете, видны чередующиеся зоны с разной оптической плотностью (рис. 4). Это позволило разделить сократительные элементы поперечнополосатых мышечных волокон (миофибрилл) на функциональные единицы - саркомеры (участки между соседними Z-мембранами).
Рис. 4. Микрофотография участка поперечнополосатой мышцы
Характерная оптическая плотность участков саркомера обусловлена особенностями расположения в них (рис. 5) сократительных белков (актина и миозина).
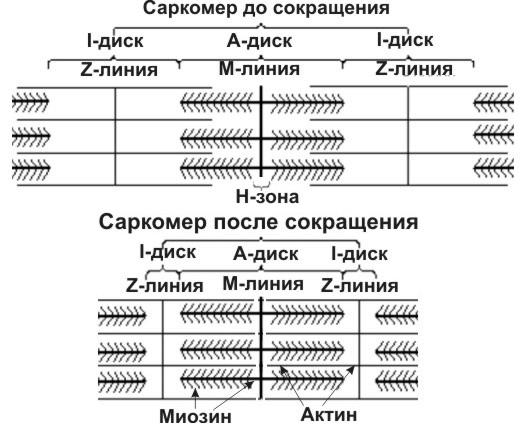
Рис. 5. Схема расположения сократительных белков в саркомере