

[Sazanov_A.A.]_Genetika(BookFi.org)
.pdfпахитена. Происходит дальнейшая конденсация хро-
мосом;
диплотена. Распадаются синаптонемальные комплексы. Происходит разделение несестринских хроматид, которые остаются связанными друг с другом в отдельных точках, где образуются Х-образные структуры – хиазмы;
диакинез. Происходит терминализация хиазм – хиазмы сдвигаются к теломерам. Происходит полное разделение гомологичных хромосом.
В конце метафазы I гомологичные хромосомы начинают расхождение к полюсам клетки. Анафаза I и телофаза I проходят аналогично анафазе и телофазе обычного митотического деления.
Эквационное деление является обычным митотическим делением гаплоидной клетки. Оно включает профазу II, метафазу II, анафазу II и телофазу II.
Рис. 38. Обобщенная схема мейотического деления:
1–5 – профаза I, 1 – лептотена, 2 – зиготена, 3 – пахитена, 4 – диплотена, 5 – диакинез, 6 – метафаза I, 7 – анафаза I, 8 – телофаза I, 9 – профаза II, 10 – метафаза II,
11 – анафаза II, 12 – телофаза II
80
У человека формирование мужских половых клеток – сперматозоидов и женских – яйцеклеток имеет существенные биологические особенности (рис. 39, 40).
Начиная с момента полового созревания (в постпубертантный период) сперматогенез у мужчин происходит постоянно. В семенниках сперматоциты – клетки-предшественники сперматозоидов подвергаются мейотическому делению. Из одного сперматоцита получается четыре сперматозоида, каждый из которых содержит плотно упакованный гаплоидный набор хромосом и митохондрию, которая служит для выработки энергии для жгутика. Жгутик позволяет сперматозоиду быстро передвигаться в жидкой среде. Следует отметить, что от зачатия до начала мейоза в семенниках клеткипредшественники сперматозоидов проходят большое число митотических делений, в ходе которых могут накапливаться мутации. В этом проявляется эволюционная роль мужского пола как носителя наследственной изменчивости. Так, при сравнении последовательностей ДНК хромосом шимпанзе и человека выяснилось, что быстрее всего накапливаются замены нуклеотидов именно в Y-хромосоме, что служит прекрасной иллюстрацией этого положения.
Эволюционная роль женского пола как хранителя уже устоявшихся наследственных изменений, как правило, имеющих адаптивную (приспособительную) ценность, выражается и в особенностях созревания яйцеклеток. Мейоз начинается в яичниках девочек на третьем месяце внутриутробного развития, когда с момента оплодотворения прошло не так много митотических делений и еще не успели накопиться мутации. До седьмого месяца делящиеся клетки находятся в стадии лептотены и зиготены. От седьмого месяца до рождения успевают пройти пахитены и диплотены, затем мейоз останавливается, образуются кариолемма и ядрышко. Эта особая стадия – диктиотена – продолжается до первой овуляции. При рождении у девочки имеется около 2,6 х 106 ооцитов, из которых большинство (около 90 %) дегенерирует. За всю жизнь женщины только около 400 ооцитов созревают
81
полностью. В яичнике каждый ооцит окружен слоем эпителиальных клеток и двумя слоями соединительной ткани – эта структура называется фолликул. В постпубертантный период (в среднем с 12 лет) в первой половине цикла под действием лютеинизирующего гормона начинается диакинез. В ходе редукционного деления образуется первое полярное тельце. Первое полярное тельце, как правило, позднее проходит эквационное деление. Далее мейоз проходит до метафазы II и останавливается до встречи яйцеклетки со сперматозоидом в фаллопиевой трубе. Проникновение сперматозоида стимулирует расхождение хроматид, формирование ядерной мембраны, женского пронуклеуса и второго полярного тельца. Из хроматина сперматозоида образуется мужской пронуклеус, который через несколько часов сливается с женским пронуклеусом, образуя зиготу – оплодотворенную яйцеклетку. Путем дробления зиготы образуется эмбрион.
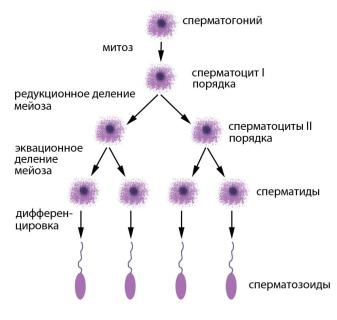
Рис. 39. Сперматогенез человека: сперматогонии – предшественники сперматоцитов, способные к митотическому делению;
сперматоциты I порядка – диплоидные продукты митотического деления сперматогониев; сперматоциты II порядка – гаплоидные продукты редукционного деления сперматоцитов I порядка; сперматиды – гаплоидные продукты эквационного деления сперматоцитов II порядка
82
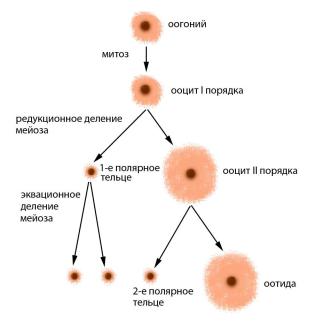
Рис. 40. Оогенез человека: оогонии – предшественники ооцитов, способные к митотическому делению; ооциты I порядка – диплоидные продукты
митотического деления оогониев; ооциты II порядка – гаплоидные продукты редукционного деления ооцитов I порядка; оотида – крупная яйцеклетка, возникшая при эквационном делении ооцита II порядка
4.3. Структурно-функциональнаяорганизацияхромосом
Хроматин – это вещество, из которого состоят хромосомы эукариотических клеток, представляющее собой комплекс ДНК, РНК и белков. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК.
Основную массу хроматина составляют белки гистоны, котрые ответственны не только за укладку ДНК, но выполняют и регуляторную функцию. Гистоны H2A, H2B, H3 и H4 образуют нуклеосомы – особые структуры шайбовидной формы высотой 6 нанометров и 11 нанометров в диаметре, участвующие в самом первом этапе упаковки ДНК в хромосомы (рис. 41). В состав каждой нуклеосомы входит по две молеку-
83
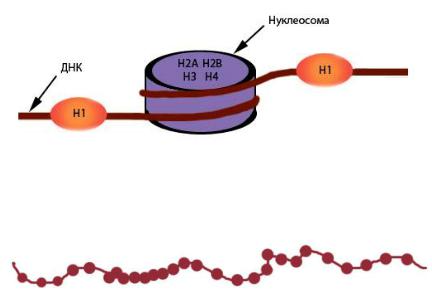
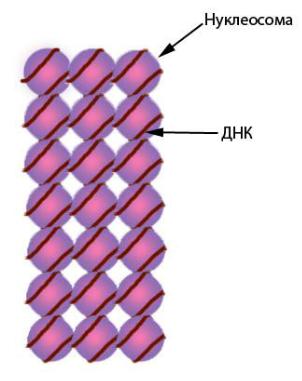
лы каждого из указанных видов гистонов (всего восемь молекул, такие комплексы называют октамерами). Из-за того что нуклеосомы располагаются более или менее регулярно, образующаяся структура напоминает бусы (рис. 42). Наиболее крупный из всех гистонов – гистон H1 – связывается с ДНК на участке между нуклеосомами (рис. 41). Компактизация и декомпактизация хроматина на уровне нуклеонемы (нуклеосомной нити) регулируется гистоном H1. При репликации – удвоении нити ДНК – нуклеосомы не распадаются, а переходят на одну из дочерних нитей случайным образом. Недостающие октамеры синтезируются заново. Самый первый уровень укладки хромосом – нуклеонемный – обеспечивает компактизацию ДНК в шесть–семь раз.
Рис. 41. Схема нуклеосомной организации хроматина.
Взаимное пространственное расположение гистонов H2A, H2B, H3 и H4 не отражено
Рис. 42. Нуклеосомная нить – нуклеонема
Нить ДНК с нуклеосомами образует структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу (рис. 43). Ее можно увидеть на препаратах выделенного хроматина. Удаление даже некоторой части молекул гистона H1 приводит к распадению фибриллы до толщины 10 нанометров, что соответствует примерной толщине нук-
84
леосомы. Это свидетельствует о том, что гистон H1 играет ключевую роль во взаимодействии между нуклеосомами. На уровне фибриллы 30 нанометров достигается примерно сорокакратная компактизация ДНК. На таком уровне спирализации существенно снижается способность ДНК связываться с белками, участвующими в транскрипции, что приводит к уменьшению генетической активности упакованных в фибриллу 30 нанометров участков.
Рис. 43. Фибрилла толщиной 30 нанометров
Около 20 % всех белков хроматина составляют негистоновые белки. Именно они имеют значение для более высоких уровней компактизации хроматина. Выявляемые на препаратах клеточного ядра шарики диаметром 100 нанометров (хромомеры) имеют петлевую розетковидную структуру (рис. 44). Это третий уровень компактизации, обеспечивающий примерно 600–700-кратное укорочение ДНК.
85
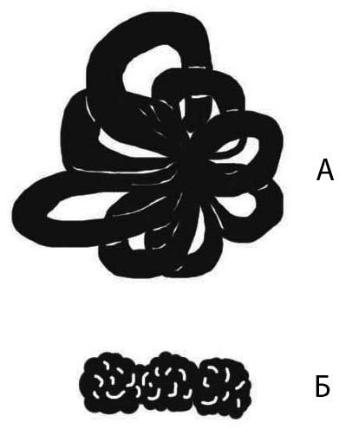
Рис. 44. Петлевой домен: А – структура в виде розетки; Б – хромонема
На препаратах митотических хромосом иногда можно увидеть спираль, образованную нитчатой хроматиновой структурой толщиной около 200 нанометров – хромонемой (рис. 44, 45). Это четвертый уровень укладки хроматина. Витки хромонемы образуют хроматиду толщиной примерно 500 нанометров, что обеспечивает укорочение ДНК примерно в 104 раз.
86

Рис. 45. Хромонемная организация митотической хромосомы
4.4. Дифференциальное окрашивание и блочная организация хромосом
При окрашивании митотических хромосом ацидофильными (т. е. связывающимися с кислотами) красителями, например эозином, или неспецифическими к нуклеотидам флуоресцентными красителями (флуорохромами – веществами, способными излучать световые волны большей длины при возбуждении светом с меньшей длиной волны) выявляется практически равномерный рисунок без исчерченности. Такая окраска называется рутинной (рис. 46).
87
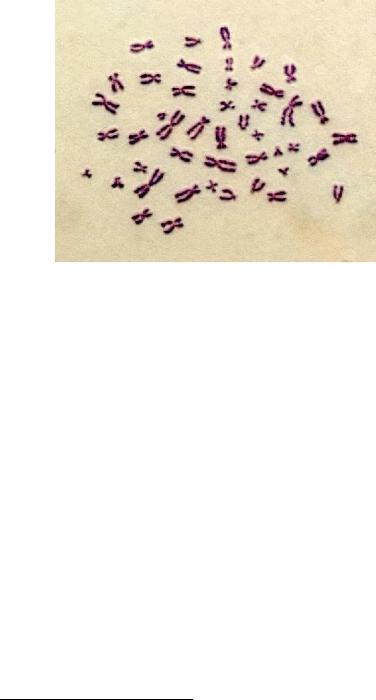
Рис. 46. Митотические хромосомы человека, окрашенные рутинно, краситель Романовского-Гимзы – раствор эозина и метиленового синего
Если перед окрашиванием митотические хромосомы обработать щелочными растворами, вымывающими ДНК в первую очередь из районов с низкой плотностью хроматина, можно наблюдать интенсивное окрашивание прицентромерных районов всех хромосом и практически полное интенсивное окрашивание Y-хромосомы (рис. 47). Такой подход называется C-методом дифференциального окрашивания хромосом или C-бендингом. C-положительные (интенсивно окрашенные при использовании этого метода) районы хромосом соответствуют участкам генетически инертного хроматина – или гетерохроматина. Различная степень окрашивания районов хромосом при C-окраске свидетельствует о неоднородности распределения гетерохроматина по длине хромосом.
URL: http://131.229.114.77/microscopy/galllm.html
88
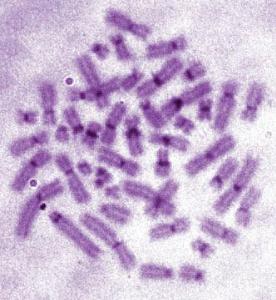
Рис. 47. Митотические хромосомы человека, окрашенные по C-методу (фото Е.Г. Нероновой)
В зависимости от степени плотности упаковки фибриллы 30 нанометров различают более конденсированный хроматин – гетерохроматин – и менее конденсированный эухроматин. Гетерохроматиновые участки очень интенсивно окрашиваются при дифференциальной C-окраске хромосом из-за большей плотности и хорошо видны под микроскопом. ДНК, находящаяся в гетерохроматине, не транскрибируется и содержит большое количество повторяющихся последовательностей. На стадии интерфазы гетерохроматин часто располагается по периферии ядра (пристеночный гетерохроматин). В эухроматине ДНК упакована сравнительно неплотно. Гены, находящиеся в эухроматиновых районах, активно транскрибируются. Плотность упаковки хроматина зависит от модификаций гистонов (метилирования, ацетилирования, фосфорилирования), в первую очередь это относится к гистону H1. Гетерохроматин у млекопитающих состоит преимущественно из АТ (аденин-тимин)-обогащенной высокоповторяющейся ДНК. Таким образом, C-окраска позволяет выявлять наиболее различающиеся по своему составу и плотности районы хромосом.
89