

[Sazanov_A.A.]_Genetika(BookFi.org)
.pdfРис. 60. Пуф
В некоторых клетках растений и животных (например, в трофобластах млекопитающих) наблюдается политения с неполной конденсацией (скрытая политения) – только отдельные районы хромосом (как правило, гетерохроматиновые) представлены политенными участками, в остальных районах многократно редуплицированные хроматиды не связаны друг с другом. Поэтому скрытая политения не нашла настолько широкого применения в общей генетике, как истинная политения хромосом двукрылых насекомых.
URL: http://www.sbs.utexas.edu/genetics/genweb/images/puff.gif
110
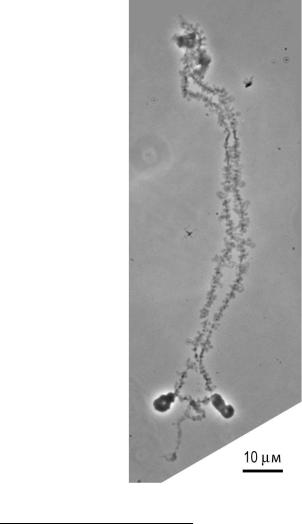
4.9. Хромосомы типа ламповых щеток
В оогенезе некоторых животных (земноводных, рептилий, птиц) встречается особая форма хромосом, отличающаяся крупными размерами и характерной петлевой структурой (рис. 61), которые из-за сходства с ершиками для чистки плафонов керосиновых ламп назвали хромосомами типа ламповых щеток. Они представляют собой биваленты на стадии диплотеныпрофазыI мейоза(разд. 4.2).
Рис. 61. Хромосома 2 домашней курицы (GGA2)
URL: http://www.biomedcentral.com/1471-2164/9/168/figure/F3?highres=y
111
Каждый бивалент состоит из двух гомологичных хромо-
сом, состоящих в свою очередь из двух сестринских хроматид
(рис. 62). Попарно симметричные петли образованы активно транскрибируемым хроматином. При помощи электронного микроскопа можно увидеть несколько транскриптов на каж-
дой петле, причем, как правило, длина транскриптов возрас-
тает от одного участка прикрепления петли до другого.
Настолько интенсивная транскрипция требуется в ходе созре-
вания женских половых клеток для превителлогенеза – нако-
пления мРНК перед формированием желтка и, в меньшей степени, вителлогенеза – собственно формирования желтка.
Определенный рисунок петель характерен для отдельных хромосом и отдельных хромосомных районов, т. е. петли мо-
гут служить морфологическими маркерами. Интересно, что в хромосомах типа ламповых щеток активно транскрибируется обычно «молчащая» высокоповторяющаяся ДНК. В местах прикрепления петель на хромосомах видны узелки – хромо-
меры, в которых находится более плотно упакованный инерт-
ный в транскрипционном отношении хроматин (рис. 62).
Наиболее детально хромосомы типа ламповых щеток изучены у гребенчатого тритона и домашней курицы. Такая форма хромосом нашла применение в качестве модельного объекта для изучения особенностей транскрипции и анализа распределения событий генетической рекомбинации по дли-
не хромосом. В последние годы активно проводятся исследо-
вания хромосом типа ламповых щеток на нескольких видах птиц.
112
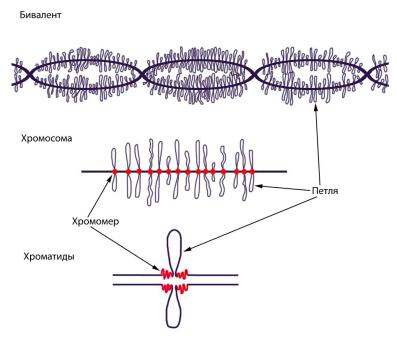
Рис. 62. Схема строения хромосом типа «ламповых щеток»
113
Глава 5. Основы генетики пола
Несмотря на то что различия между полами сложны и многообразны, в общем виде пол можно рассматривать как простой дискретный признак, имеющий, как правило, два альтернативных проявления – женский и мужской.
Существует большое число типов определения пола.
Принципиально их можно разделить на генетический тип – связанный с различиями генов и хромосом у разных полов – и
ненаследственный. К последнему относятся системы опреде-
ления пола под влиянием условий окружающей среды. На-
пример, у морского червя Bonellia viridis личинка,
прикрепившаяся к хоботу самки, развивается в самца, кото-
рый проводит всю жизнь в ее матке, занимаясь исключитель-
но оплодотворением яиц. Если личинка прикрепляется ко дну, она развивается в самку – самостоятельную особь. У кро-
кодилов пол детёнышей определяется температурой инкуба-
ции яиц в средней трети периода эмбрионального развития.
Например, у нильского крокодила при температуре от 31,7 °C
до 34,5 °C формируются самцы, при более низкой или более высокой температуре – самки. У некоторых рыб пол может изменяться несколько раз в течение жизни.
В случае генетического определения пола небольшое число генов, которые расположены, как правило, в половых хромосомах (набор которых различается у разных полов), на-
правляет развитие по женскому или мужскому пути (рис. 63).
Система хромосомного определения пола гаплоид-диплоид встречается у некоторых перепончатокрылых – из оплодотво-
ренных яиц развиваются диплоидные самки, а из неоплодо-
творенных – гаплоидные самцы. Биологическое назначение хромосомных механизмов определения пола систем X0, ZW и
114
XY – поддерживать равное соотношение полов для повыше-
ния биологического разнообразия. Система X0 присутствует у некоторых насекомых, самки имеют две половых хромосомы,
самцы – одну. Прямо противоположная картина наблюдается у тли. При наличии двух половых хромосом различают ZW и XY системы определения пола. Если гетерогаметным полом,
т. е. полом, в мейозе которого образуются гаметы с разным набором половых хромосом, является женский (например,
птицы, некоторые рептилии), система называется ZW (рис. 63). При системе XY (например, дрозофила, плацентар-
ные млекопитающие) гетерогаметным полом являются сам-
цы (рис. 63). Интересно, что в последнем случае пол может определяться как присутствием Y-хромосомы, как у плацен-
тарных млекопитающих, так и соотношением числа аутосом и числа X-хромосом (балансовый тип), как у дрозофилы. У
дрозофилы при диплоидном наборе аутосом и наличии од-
ной X-хромосомы формируются самцы безотносительно на-
личия и количества Y-хромосом. Присутствие двух X-
хромосом на диплоидный набор ведет к развитию самок, че-
му не препятствует наличие Y-хромосомы. У плацентарных млекопитающих присутствие одной или более Y-хромосомы приводит к формированию мужского пола, несмотря на лю-
бые иные обстоятельства. Это объясняется наличием в
Y-хромосоме гена TDF-SRY, определяющего развитие по мужскому пути (рис. 63). При транслокациях участка
Y-хромосомы, содержащего этот ген, на X-хромосому у чело-
века встречались мужчины с набором хромосом 46, XX и жен-
щины с набором хромосом 46, XY.
115
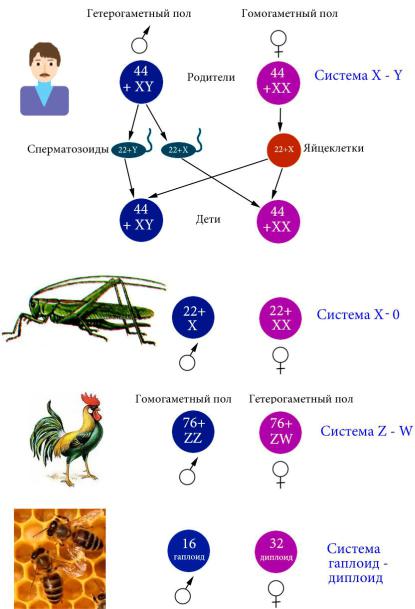
Рис. 63. Типы хромосомного определения пола у животных
116
У человека, подобно всем плацентарным млекопитающим, принято различать четыре уровня половой дифференцировки (рис. 64):
1)хромосомный пол (XX или XY);
2)гонадный пол (яичники или семенники);
3)фенотипический пол (женский или мужской; внешние половые признаки);
4)поведенческий и метаболический пол (характерные мужские или женские особенности поведения и обмена веществ).
Под влиянием TDF-SRY из зачатка гонады формируются семенники. При отсутствии этого гена зачаток гонады развивается по женскому типу. Половые органы формируются из мюллеровых и вольфовых протоков, которые происходят из первичной почки. У женщин мюллеровы протоки развиваются в фаллопиевы трубы и матку, а вольфовы протоки атрофируются. У мужчин вольфовы протоки развиваются в семенные протоки и семенные пузырьки. В клетках Сертоли, расположенных в извитых канальцах семенников, синтезируется антимюллеров гормон, который ингибирует развитие мюллеровых протоков и приводит к их атрофии. Клетки Лейдига, расположенные между канальцами в семенниках, синтезируют стероидные гормоны тестостерон и 5-дигидротестостерон, которые определяют развитие гениталий и, позднее, формирование мужских вторичных половых признаков (мужской тип оволосения, низкий голос, особенности телосложения). Интересно, что для нормального развития по мужскому типу требуется нормальное функционирование всех этих компонентов, а развитие по женскому типу происходит «конститутивно». Поведенческий
иметаболический пол формируются постепенно, достигая полноты проявления в постпубертантный период.
117
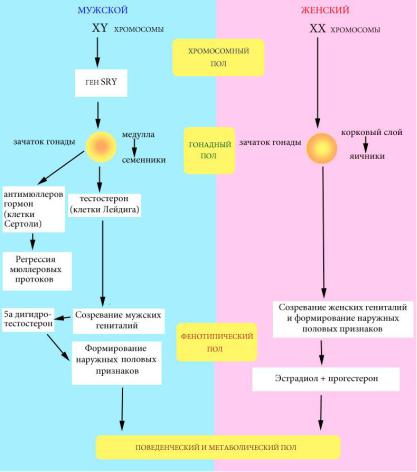
Рис. 64. Схема определения пола у человека
Контрольные вопросы и задания
1. Самка медоносной пчелы Apis mellifera диплоидна, а трутни гаплоидны. Существует, однако, локус с множественными аллелями, влияющий на определение пола у диплоидных особей. Все диплоидные особи, гетерозиготные по аллелям этого локуса – самки, а гомозиготные по любому из аллелей – самцы. Гомозиготные самцы не достигают половозрелости, поскольку обычно поедаются рабочими пчелами на личиночной стадии в первые трое суток после вылупления из яичек. Таким образом, гомозиготный генотип в естествен-
118
ных условиях летален. Однако изъятых из улья личинок таких самцов можно довести до стадии половозрелости. В этом локусе насчитывается около двадцати аллелей, обозначаемых символами A1, A2, ... , A20. Пусть в потомстве некоторой оплодотворенной гаплоидным трутнем матки лишь половина яичек способна к развитию (это хорошо заметно, поскольку половина ячеек в сотах остается пустой, и пустые ячейки распределены по сотам случайным образом). Что можно сказать
огенетической конституции матки и трутня?
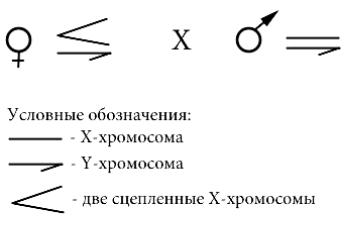
2.Рассчитайте соотношение полов у гибридов первого поколения от скрещивания дрозофил (рис. 65)
3.У тутового шелкопряда хромосомный механизм определения пола ZW. Известно, что самцы дают на 20 % больше шелка, чем самки. Самцы линии А являются сбалансированными гетерозиготами по двум сцепленным с полом летальным мутациям, а самцы линии Б – нормальные. Какую линию лучше использовать в качестве отцовской для получения гибридов для выкормки? Ответ обоснуйте.
Рис. 65. Схема скрещивания (для задачи 2 к гл. 5)
119