

Подготовка у универсиаде 2012 / Генетика (Жимулев) / 5ver7
.pdfГлава 5 |
Генетический анализ |
|
|
Глава 5. Генетический анализ: картирование генов
5.1. Цели и задачи генетического анализа
Определяя понятие генетического анализа, А.С. Серебровский писал: “Генетическим анализом мы называем систему опытов, наблюдений и вычислений, имеющих целью разложение свойств (признаков) организма на отдельные наследственные элементы, “отдельные признаки”, и изучение свойств соответствующих им генов” (Серебровский, 1970, стр. 5).
ПомнениюМ.Е.Лобашева,спомощью генетического анализа исследуется качественный и количественный состав генотипа,проводитсяанализегоструктурыи функционирования.
Методы генетического анализа разнообразны, но, главным образом, это системавсевозможныхскрещиваний,причем любая работа проходит следующие этапы:
1.Выясняется-наследуетсялипризнак,имеет лионконтрастныеформы.
2.Выясняетсячислогенов,контролирующих развитиеданногопризнака.
3.Выясняется - есть ли взаимодействие междуэтимигенами.
4.Определение группы сцепления (хромосомы) и картирование гена в хромосоме.
5.Характеристикагенов.
Âнастоящее время в понятие генетического анализа входит клонирование гена, определение последовательности нуклеотидов ДНК, выяснение интронэкзонной структуры гена, экспрессии гена в онтогенезе.
Картированиегеновсначаласточностью до группы сцепления, а затем уже в определенном участке хромосомы, является важнейшейчастьюгенетического анализа.
У дрозофилы анализ проходит следующиеэтапы:
1.Получениемутаций,спонтанныхили индуцированных.
2.Тестмутацийнааллелизм.
3.Картированиегенавгруппе сцепления.
4.Построение кроссоверных карт генов.
5.Картирование генов с помощью хромосомных перестроек.
6.Картирование генов с помощью гибридизацииinsitu.
Необязательнопроводитьвсеэтиэтапы. ЕсливрукахисследователяестьДНКданного гена, то для его хромосомной локализации легче сразу провести гибридизацию in situ. Однако, чаще всего именно для получения ДНК гена, необходимо провести все этапы генетическогоанализа.
Дляизучениянаследованияпризнакову человека гибридологический анализ не применим. Его успешно заменяет генеалогический метод. Генеалогическое древо,илиродословная,строится,начинаяот персоны, обладающей определенным признаком.
Уорганизмов,генетикакоторыхразвита недостаточно хорошо, картирование генов проводят с помощью методов клеточной биологии.
5.2. Получение мутаций
Спонтанно возникшие мутации выявляют, главным образом, в результате обследования диких популяций изучаемого организма.
Индуцированные мутации получают с помощью различных схем скрещиваний, в зависимости от наличия предварительных знаний о локализации гена. Если известна только локализация гена с точностью до хромосомы, то получают новые мутации, картированные в пределах этой хромосомы. У дрозофилы используют такие схемы скрещиваний как ClB для Х-хромосомы или Cy L/Pm для второй хромосомы (см. Раздел 4.2.1.). Известны и другие схемы.
Еслипредварительноизвестныделеции в определенном районе хромосомы, существуют специальные схемы скрещиваний, позволяющие буквально “насытить”мутациямиэтотрайон.Такойметод был разработан в 1938 г. С.И. Алиханяном, а затембылуспешнопримененамериканскими
82

Генетический анализ |
|
|
Глава 5 |
Рисунок 5.1 |
|
|
|
|
|
мутаген |
|
Basc, B |
|
|
|
P |
|
|
|
Basc, B |
|
|
|
|
Basc, B |
Df |
|
F1 |
|
|
|
|
l |
Dp |
|
Basc, B |
l |
Basc, B |
l |
F2 |
|
|
|
Df |
Df |
Dp |
Dp |
Схема скрещиваний для получения мутаций в определенных районах Х-хромосомы дрозофилы (Из: Judd et al., 1972).
учеными Б. Джаддом и его сотрудниками в 1972 году (Рис. 5.1). Самок, гетерозиготных по балансерной линии Basc, несущей доминантную мутацию Bar (B), скрещивают
ñобработанными мутагеном самцами. В потомстве отбирают самок, несущих хромосому Basc и мутагенизированную хромосому,и скрещивают их индивидуально
ñсамцами, имеющими делецию в Х- хромосоме и дупликацию Х-хромосомного материала в Y-хромосоме. Наличие дупликации в Y-хромосоме, “перекрывающей” нехватку генов в Х- хромосоме, позволяет этим самцам выжить.
ÂF2 вкаждойиндивидуальнойпробирке анализируютрасщеплениесредисамок.Если вмутагенизированнойХ-хромосомевозникла летальная мутация в участке, удаляемом
Рисунок 5.2
l1
l2
мутантный фенотип
|
|
|
l |
1 |
+ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
+ |
|
l2 |
||||||
нормальный фенотип
Функциональный тест на аллелизм. Вверху - гетероаллельная комбинация или компаунд, внизу - классическая дигетерозигота.
делецией,тосамкиBar+ гибнут.Втожевремя самцыBar+ являютсяносителямиэтойлетали.
Впоследниегодымутацииудрозофилы индуцируют с помощью мобильного элемента генома (Р-элемента). Наличие мутаций, индуцированных таким образом, позволяет выделить и клонировать ДНК данногогена.
5.3.Тест мутаций на аллелизм
Âрезультатемутагенизации,какправило, получают большое число мутаций. Для того, чтобы определить, сколько и какие гены они затрагивают, проверяют все мутации в гетерозиготе во всех парных сочетаниях с цельюустановить,нарушаютмутацииоднуи ту же функцию или разные. Согласно этому функциональному критерию, если не поврежденные мутациями участки генов взаимодействуют комплементарно и образуется гибрид дикого типа, то мутации затрагивают разные гены. Если же у гибрида получаетсямутантныйфенотип,этоозначает, что обе мутации затрагивают один и тот же
ãåí (Ðèñ. 5.2).
5.4. Межаллельная комплементация
Этотакаяситуация,когдадвамутантных аллеля совместно способны обеспечить
83

Глава 5 |
Генетический анализ |
|
|
образование нормально функционирующего продукта, а в отдельности ни один из них это сделать не в состоянии. Как находят факты межаллельнойкомплементации?
В генетических экспериментах очень часто выделяют ряд мутаций, имеющих, по крайней мере, на первый взгляд, сходное фенотипическоевыражение,локализующихсяв одном и том же небольшом генетическом сегменте и не дающих комплементации при проверке на аллелизм. Такие мутации приписывают одному и тому же гену. Затем индуцируютвсеновыеиновыемутациитакого жетипаи,когдаихчислостановитсядостаточно большим, выясняется, что хотя почти все они по-прежнемунекомплементарны,внекоторых комбинациях можно обнаружить комплементарность. Явление это широко распространено.Частобываеттак,чтонарядус классами мутаций, комплементарных между собойвлюбыхпарныхсочетаниях,существуют и другие мутации, не комплементарные мутациям двух или более комплементарных классов (перекрывающиеся с ними на комплементационныхкартах),атакжемутации (они обычно составляют абсолютное большинство), перекрывающиеся со всеми остальными. Комплементационные взаимоотношениявсейсовокупностимутантов можно представить в виде схемы, или комплементационной карты, которая очень часто, хотя и не всегда, бывает линейной. В основупостроениякомплементационнойкарты положен следующий принцип: все взаимно комплементарныемутацииизображаютсяввиде неперекрывающихся линий, а все некомплементарные-ввидеперекрывающихся (Рис.5.3).Группамутаций,обнаруживающих одинаковыекомплементационныеотношениясо всемидругимимутациями,называетсягруппой комплементации.
Предпосылками межаллельной комплементации являются независимое действиеаллелейиналичиегибридногобелка - продукта данного гена. Согласно гипотезе КэтчсайдаиОвертона(см.Финчем,1968,стр. 93), средипродуктовгенасуществуетдватипа полипептидных цепей. Предполагается, что
одной из стадий в образовании фермента является агрегация не менее двух полипептидных цепей. В некоторых случаях объединяются гетерологичные продукты. Затем, по-видимому, происходит специфическоесвертывание полипептидных цепей. У каждого мутанта имеется какой-то дефект, обусловливающий не совсем правильное свертывание и, как следствие, образование белка, достаточно сильно отличающегося от нормального фермента и потомулишенногоспецифическойактивности. Однаизпричин,покоторымразныемутантные полипептидные цепи могут оказаться комплементарными, - это способность к кооперативному свертыванию в гетерологичныхагрегатах(Рис.5.4).
Одной из важных характеристик межаллельной комплементации является изменение карты комплементации при варьированиитемпературы(Рис.5.5).
Существование межаллельной комплементации требует более тщательного анализа аллелизма мутаций. Скрещивания необходимо проводить при разных температурах:у дрозофилы это 180C, 250C è 290C.
Рисунок 5.3
51
|
2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2 |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5 |
|
|
|
3 |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
||
|
1 |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6 |
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
||
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
||
|
1 |
|
1 |
|
3 |
|
||
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
1 |
3 |
1 |
5 |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Карта комплементации мутаций гена ad-4 у Neurospora crassa (Из: Финчем, 1968, стр. 54). Цифры указывают число ауксотрофных мутаций в каждой группе коплементации.
84

|
Генетический анализ |
|
|
|
|
|
|
|
|
|
|
Глава 5 |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рисунок 5.4 |
|
|
|
Рисунок 5.5 |
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
m1 |
|
m2 |
m3 |
m1 |
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
m4 |
|
|
m3 |
m2 |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
m5 |
|
|
m4 |
m5 |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
m6 |
|
|
|
m6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
1 |
2 |
|
|
|
|
250C |
|
|
180C |
|||||
|
|
|
|
|
Карты комплементации одних и тех же |
||||||||||
|
|
|
|
|
мутаций при разных температурах. |
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
среди прочих гемофилия и дальтонизм у |
|||||||||||
|
|
|
|
человека. |
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
Для локализации гена в аутосоме |
|||||||||
|
|
|
|
используюттесторныелинии. |
|
|
|
|
|||||||
|
|
|
|
5.5.1. Определение группы |
|
|
|||||||||
|
3 |
4 |
|
сцепления с помощью |
|
|
|||||||||
|
Схема комплементации двух полипептидных |
|
рецессивных маркеров |
|
|
|
|||||||||
|
цепей молекулы белка, состоящей из двух |
|
|
|
В качестве линии-тестора можно взять |
||||||||||
|
одинаковых цепей (Из: Гершензон, 1983, стр. |
|
|
|
|||||||||||
|
|
линию,вкотороймаркированывсехромосомы |
|||||||||||||
|
399). 1 - нормальная конфигурация цепи; 2 и |
|
|||||||||||||
|
|
дрозофилы, кроме половой: мутация brown |
|||||||||||||
|
3 - обе цепи имеют по-разному искаженную |
|
|||||||||||||
|
|
(bw,коричневые глаза)-вовторойхромосоме, |
|||||||||||||
|
конфигурацию вследствие двух мутаций; 4 - |
|
|||||||||||||
|
|
мутация ebony (e, черное тело) - в третьей |
|||||||||||||
|
при сборке молекулы белка цепи взаимно |
|
|||||||||||||
|
|
хромосоме и eyeless (ey, отсутствие глаз) - |
|||||||||||||
|
исправляют свою конфигурацию и молекула |
|
|||||||||||||
|
оказывается функционально нормальной. |
|
в четвертой хромосоме. |
|
|
|
|
||||||||
|
Стрелки указывают положение аминокислот, |
|
|
|
Самок тесторной линий скрещивают с |
||||||||||
|
измененных мутацией. |
|
|
носителем картируемой мутации – в данном |
|||||||||||
|
|
|
|
случае это доминантная мутация Mut, |
|||||||||||
5.5. Определение группы |
|
||||||||||||||
|
локализованная в третьей хромосоме (Рис. |
||||||||||||||
сцепления |
|
|
|||||||||||||
|
|
5.6). Самцов F1 нужно скрестить с самками, |
|||||||||||||
|
Если признак переда¸тсятолькоототца |
||||||||||||||
|
взятыми вновь из тестерной линии. Если в F |
||||||||||||||
к сыну, то весьма очевидно, что данный ген |
|
|
|
|
|
|
|
|
|
|
2 |
||||
мутация Mut проявляется только с маркером |
|||||||||||||||
картирован в Y-хромосоме. Например, у |
второй и четвертой хромосом (bw и ey наРис. |
||||||||||||||
человека встречается такой признак, как |
5.6) и никогда – с маркером третьей, значит |
||||||||||||||
волосатыеуши.Онбываеттолькоумужчини |
эта мутация располагается в третьей |
||||||||||||||
передаетсяпомужскойлинии.Генкартирован |
хромосоме. Если картируемая мутация |
||||||||||||||
в Y-хромосоме. |
|
|
никогда не проявляется одновременно с |
||||||||||||
|
Многочисленные“сцепленныесполом” |
||||||||||||||
|
маркерами второй и четвертой хромосом, то |
||||||||||||||
генылокализованыв Х-хромосоме.Характер |
|
она локализована, соответственно во второй |
|||||||||||||
их наследования отличается от всех других: |
или четвертой хромосомах. |
|
|
|
|
||||||||||
1. Реципрокные скрещивания дают разные |
|
|
|
|
|
|
|
|
|
|
|
|
|||
результаты. 2. В одном направлении |
5.5.2. Определение группы |
|
|
||||||||||||
скрещивания потомки F1 единообразны, в |
сцепления с помощью |
|
|
||||||||||||
другом - наблюдается расщепление, причем |
доминантных маркеров |
|
|
||||||||||||
дочери похожи на отца, а сыновья на мать |
|
|
Можноиспользоватьлиниюсчетырьмя |
||||||||||||
(наследование крест-накрест, или крисс- |
|
|
|||||||||||||
доминантнымимаркерами:L(Lobe,дольчатые |
|||||||||||||||
кросс). К числу таких признаков относятся |
|
|
|
|
|
|
|
|
|
|
|
|
|||
85
Глава 5 |
|
|
|
|
|
|
|
|
|
|
|
|
Генетический анализ |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
признаком L и у мух без |
||
Рисунок 5.6 |
|
|
|
|
|
|
|
|
|
|
||||||
|
bw |
e |
|
ey |
+ |
|
Mut |
+ |
|
маркерных признаков, но не |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
проявится у мух с маркерами |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
третьей хромосомы. |
|
|
|
bw |
e |
|
ey |
+ |
+ |
+ |
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3. Åñëè |
мутация |
mut |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
локализована в хромосоме 4, то |
||
|
bw |
e |
|
ey |
bw |
|
Mut |
|
ey |
она проявится у половины мух с |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
признаком L и у половины мух с |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
признакомSb, и у половины мух |
||
|
bw |
e |
|
ey |
+ |
|
e |
+ |
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
без маркеров. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.6. Локализация гена |
|||||
|
|
bw |
|
Mut |
|
ey |
|
+ |
|
|
Mut |
|
|
ey |
|
|||||||||
|
|
|
|
|
|
|
|
|
|
в группе сцепления |
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
bw |
|
e |
|
ey |
|
|
bw |
e |
|
|
ey |
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.6.1. Классический |
||||||
|
|
bw |
|
e |
|
ey |
+ |
|
|
e |
|
|
ey |
|||||||||||
|
|
|
|
|
|
|
|
метод |
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Допустим, |
нужно |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
bw |
|
e |
|
ey |
|
bw |
e |
|
|
ey |
|
|
||||||||||
|
|
|
|
|
|
|
|
прокартировать |
мутацию, |
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
bw |
|
e |
+ |
|
|
+ |
|
|
e |
+ |
|
|
определяющую у дрозофилы |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
желтыйцветтела-yellow (y).Для |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
bw |
|
e |
|
ey |
|
bw |
e |
|
|
ey |
|
локализации y,этулиниюнужно |
||||||||||
|
|
bw |
|
Mut |
+ |
|
|
+ |
|
|
Mut |
+ |
|
|
скрестить с тесторной, несущей |
|||||||||
|
|
|
|
|
|
|
|
какминимумдвамаркерныхгена: |
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
sn -singed - опаленные щетинки |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
bw |
|
e |
|
ey |
|
bw |
e |
|
|
ey |
|
|||||||||||
|
|
|
|
|
|
|
|
и m - miniature - маленькие |
||||||||||||||||
|
Схема скрещиваний для картирования гена в группе |
|
||||||||||||||||||||||
|
|
крылья, поскольку вместе с y их |
||||||||||||||||||||||
|
сцепления с |
помощью |
рецессивных |
маркеров у |
|
|||||||||||||||||||
|
будеттриводнойхромосоме.Это |
|||||||||||||||||||||||
дрозофилы. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
как раз тот минимум, который |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
глаза), Cy (Curly, загнутые вверх крылья, D |
необходимдляточнойлокализациигена. |
|||||||||||||||||||||||
(Dichaete, растопыренные крылья, см. Рис. |
|
|
Ход эксперимента имеет такой вид: |
|||||||||||||||||||||
4.1), Sb (Stubble, укороченные толстые |
|
|
|
|
|
|
|
|
|
|
||||||||||||||
щетинки).Всечетыремутациидоминантны,но |
|
|
|
y sn m |
y sn m |
|
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
с рецессивным летальным действием. |
+ |
+ |
+ |
|
|
|
||||||||||||||||||
Хромосомы 4 и Х в данной схеме не метятся. |
|
|
|
|||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
Затем скрещивают особей этой линий с |
|
|
|
|
|
|
|
|
|
|
||||||||||||
гомозиготой по картируемой рецессивной |
|
|
|
|
Фенотипы |
|
|
|
||||||||||||||||
Некроссоверные |
y sn m |
7337 |
|
|||||||||||||||||||||
мутации.Назовемее mut ипредположим,что |
14671 (66.4%) |
|||||||||||||||||||||||
потомки |
|
|
|
+ |
+ + |
7334 |
||||||||||||||||||
|
|
|
|
|||||||||||||||||||||
она во второй хромосоме (Рис. 5.7). |
|
|
|
Одиночные |
|
|
|
|
|
|
||||||||||||||
|
|
ВпотомствеF1 отбираютсамцовлюбого |
|
|
y |
+ + |
1994 |
|
||||||||||||||||
|
|
кроссоверы |
|
|
4066 (18.4%) |
|||||||||||||||||||
|
|
|
|
+ sn m |
2072 |
|||||||||||||||||||
из классов, затем ставят анализирующее |
на участке y-sn |
|
||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||
скрещивание. |
|
|
|
|
|
|
|
|
|
Одиночные |
|
|
y |
sn + |
1360 |
|
||||||||
1. Если мутация mut локализована в |
кроссоверы |
|
|
2674 (12.1%) |
||||||||||||||||||||
|
|
+ |
+ m |
1314 |
||||||||||||||||||||
на участке sn-m |
|
|||||||||||||||||||||||
хромосоме 2, то она проявится у мух с |
|
|
|
|
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
признаком Sb или у мух без маркерных |
Двойные |
|
|
|
y |
+ m |
335 |
685 (3.1%) |
||||||||||||||||
признаков и не проявится у мух с маркерами |
кроссоверы |
|
|
+ sn + |
350 |
|
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
второй хромосомы. |
|
|
|
|
|
|
|
Всего потомков |
|
|
22096 |
|
||||||||||||
2. Если мутация mut локализована в |
|
|
|
|
|
|
|
|
|
|
||||||||||||||
хромосоме 3, то она проявится у мух с |
|
|
|
|
|
|
|
|
|
|
||||||||||||||
86

|
Генетический анализ |
|
|
|
|
|
|
|
|
|
|
|
Глава 5 |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
появления рекомбинантов в анализирующем |
|||||
|
Рисунок 5.7 |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
скрещивании (rf) и частотой собственно |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
кроссинговера. Однако, эти величины |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
совпадают лишь в том случае, если обе |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
составляющие (rfab è rfbc) не превышают |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
десятка единиц (морганид). На более |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
протяженных |
участках |
хромосом |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
экспериментально определяемое значение rfañ |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
оказывается всегда меньше “теоретически |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ожидаемого”(rfab +rfbc).Основываясьназаконе |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
аддитивности,расстояниеrfay можноопределить |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
двумяспособами-непосредственноизмеривего |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
в соответствующем скрещивании (a × |
y), èëè |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
суммировав значения rf, определяемые на |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
небольшихинтервалахмеждуданнойпаройгенов |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
aиyприналичиипромежуточныхмаркеровb,c,... |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
x. Понятно, что при таком последовательном |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
суммированиидаженебольшихрасстояниймы |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
можем получить значения общей длины карты |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
даннойхромосомы(еслиaиyрасположенынае¸ |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
концах)всотниитысячиморганид.Втожевремя |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
частота рекомбинации между a и y не может |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
превышать 50% - значения, наблюдаемого при |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
независимом |
наследовании |
признаков. |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
Схема скрещиваний для картирования гена |
|
Отступления от аддитивности на больших |
|||||||||||||||||
|
|
расстояниях Стертевант приписал наличию |
||||||||||||||||||
|
в группе сцепления с помощью доминантных |
|
||||||||||||||||||
|
|
двойныхкроссинговеров.Второйкроссинговер |
||||||||||||||||||
|
маркеров у дрозофилы. |
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
между a и с ликвидирует последствия первого: |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
Этиданныесвидетельствуютотом,чтовсего |
||||||||||||||||||||
рекомбинанты по a и с не появляются, хотя |
||||||||||||||||||||
кроссоверов на участке y-sn было 21,5% |
рекомбинацияимеламесто(см. Раздел 3.3.2.3.). |
|||||||||||||||||||
(18,4+3,1), а на участке sn-m – 15,2% |
Еслиинтервалмеждугенамидостаточнодлинный, |
|||||||||||||||||||
(12,1+3,1). |
|
|
|
|
|
|
|
|
|
|
|
|
точислочетныхобменовравночислунечетных, |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
Чтобы данные не противоречили друг |
инаступаетрекомбинационноеравновесие(rf= |
|||||||||||||||||||
другу гены на карте хромосомы можно |
50%). Таким образом на длинных отрезках |
|||||||||||||||||||
расположить следующим образом: |
хромосомывероятнотьрекомбинациинеявляется |
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
y |
sn |
|
|
m |
болеемеройгенетическогорасстояния. |
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Прикартированиигенов,расположенных |
||||
0.0 |
|
|
|
21.5 |
36.7 |
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
далеко друг от друга, выделяют два понятия: |
||||
21.5 |
|
|
|
15.2 |
|
|
|
|
|
расстояниенакарте(d)ичастотурекомбинации |
||||||||||
|
|
Рассуждая таким образом, Стертевант |
(rf). Расстояние подсчитывается на основании |
|||||||||||||||||
приш¸лквыводу,чторасстояниемеждудвумя |
определения суммарной частоты кроссоверов, |
|||||||||||||||||||
генами(rf)равносумме(в некоторых случаях |
частота рекомбинации учитывает только тех |
|||||||||||||||||||
- разности) расстояниймеждунимиилюбым |
кроссоверов, которые выявляются по обмену |
|||||||||||||||||||
третьимгеном,расположеннымвтойжегруппе |
маркеров. |
|
|
|
||||||||||||||||
сцепления (закон аддитивности): rfac = rfab + |
Часто взаимоотношения |
между |
||||||||||||||||||
расстоянием (d) и частотой рекомбинаций |
||||||||||||||||||||
rf . Закон аддитивности показывает, что rf |
||||||||||||||||||||
|
bc |
|
|
|
|
|
|
|
|
|
(rf) выражается картирующей функцией |
|||||||||
естьлинейнаяфункциярасстояния.Стертевант |
||||||||||||||||||||
не проводил различий между частотой |
|
(Ðèñ. 5.8). |
|
|
|
|||||||||||||||
87

Глава 5 |
|
|
|
|
|
|
|
|
Генетический анализ |
|||||
Дополнение 5.1 |
|
|
|
|
Рисунок 5.8 |
|
|
|
|
|
|
|||
|
Идея построения генетических карт по |
rf |
|
|
|
|
|
|
|
|||||
результатам рекомбинационного анализа |
|
|
|
|
|
1 |
|
|
||||||
50 |
|
|
|
|
|
2 |
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|||
принадлежит А. Стертеванту. Вот что он |
|
|
|
|
|
|
|
|
||||||
пишет в своих воспоминаниях. “В 1909 году |
40 |
|
|
|
|
|
|
3 |
||||||
Кастл (Castle) опубликовал диаграммы, |
|
|
|
|
|
|
|
|
||||||
показывающие взаимоотношение между |
30 |
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|||||||
генами, определяющими окраску кролика... |
20 |
|
|
|
|
|
|
|
||||||
Они были восприняты как попытка выявить |
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|||||||
пространственные взаимоотношения в |
10 |
|
|
|
|
|
|
|
||||||
ядре. В конце 1911 года в разговоре с |
|
|
|
|
|
|
|
|
||||||
Морганом об этой попытке... я внезапно |
|
|
|
|
|
|
|
|
||||||
понял, что вариации в силе сцепления, уже |
0 |
10 |
20 |
30 |
40 |
50 |
60 |
70 d |
||||||
приписанные Морганом различиям в |
Картирующие функции. По оси абсцисс: d (в |
|||||||||||||
пространственном расположении генов, |
%), по оси ординат rf (в %). - кривая, |
|||||||||||||
открывают возможность определения |
описываемая функцией Стертеванта (rf = d); |
|||||||||||||
последовательности в линейном измерении |
- эмпирическая кривая, построенная по |
|||||||||||||
хромосомы. Ночью я сделал первую карту” |
точкам, ордината каждой из которых |
|||||||||||||
(Sturtevant, 1965, p. 305, из Кушев, 1971, стр. |
представляет собой частоту рекомбинации |
|||||||||||||
14). |
|
|
|
|
|
для данной пары мутантов, а абсцисса - |
||||||||
Картирующая функция, предложенная |
сумму частот рекомбинаций для небольших |
|||||||||||||
последовательных интервалов (d) между |
||||||||||||||
Холдейном (J.B.S. Haldane), выражается |
||||||||||||||
данной парой маркеров; ! - кривая, |
||||||||||||||
|
|
|
|
|
|
|||||||||
формулой: |
|
|
|
|
описываемая функцией Холдейна (Из: Кушев, |
|||||||||
rf= 1 |
(1-e-2d) èëè |
d=-1ln(1-2rf), ãäå |
1971, ñòð. 17). |
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|||||||
2 |
|
|
2 |
|
|
vermilion, алые глаза). Одновременно ставят |
||||||||
e - основание натуральных логарифмов |
|
|||||||||||||
|
два скрещивания. Одно для того, чтобы |
|||||||||||||
d - расстояние на карте |
|
|
||||||||||||
|
|
определить частоты кроссинговера между |
||||||||||||
rf - частота рекомбинации маркеров |
|
|||||||||||||
|
маркерами, если бы в X-хромосоме не было |
|||||||||||||
Функция Холдейна показывает, что по |
||||||||||||||
летали (скрещивание а на Рис. 5.9), другое – |
||||||||||||||
ìåðå |
увеличения |
расстояния, |
rf |
собственнодлякартирования(скрещиваниеб). |
||||||||||
ассимптотически приближается к 0.5. |
Видно, что в скрещивании “а” число муx в |
|||||||||||||
Значения rf и d практически совпадают лишь |
комплементарных классах кроссоверов и |
|||||||||||||
на отрезке кривой до rf = 0,1 (Из: Кушев, |
некроссоверов равны. В скрещивании “б” |
|||||||||||||
1971, ñòð. 14-17). |
|
|
|
такого равенства нет, и самцы дикого типа |
||||||||||
В настоящее время кроссоверные |
||||||||||||||
встречаютсяоченьредко,чтосвидетельствует |
||||||||||||||
генетические карты составлены для очень |
о наличии летали в Х-хромосоме. Она может |
|||||||||||||
многих организмов (см. справочники: |
располагаться в одном из трех положений: |
|||||||||||||
Захаров, 1979; O’Brien, 1982; Lindsley, Zimm, |
слева от ct, между ct и v, и правее v. В |
|||||||||||||
1992). |
|
|
|
|
скрещивании“а”частотакроссоверовмежду |
|||||||||
|
|
|
|
|
|
|||||||||
5.6.2. Картирование летальных |
ct, и v составляет 13%, а некроссоверов - 86, |
|||||||||||||
приэтомвскрещивании“б”происходитпотеря |
||||||||||||||
мутаций |
|
|
|
|
||||||||||
|
|
|
|
некроссоверного |
класса |
++, |
÷òî |
|||||||
|
|
|
|
|
|
|||||||||
Можно |
рассмотреть |
принцип |
свидетельствуетолокализациилеталимежду |
|||||||||||
картирования леталей на примере мутации, |
двумя маркерами. Более того, кроссоверов |
|||||||||||||
локализованной в X-хромосоме. Используют |
между l и v (ct+) выживаетнесколько больше |
|||||||||||||
тестерную линию, то есть хромосому, |
(7,5%)чеммеждуctиl (v+)–4,5%.Расстояние |
|||||||||||||
имеющую 2 видимых маркера, в данном |
между ct (22,0) и v (33,0) составляет 11 |
|||||||||||||
случае ct и v (ct – cut, вырезки на крыльях, v – |
морганид. Леталь картируется в этом |
|||||||||||||
88

Генетический анализ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Глава 5 |
|||||||
Рисунок 5.9 |
|
|
|
|
|
|
|
|
|
мутациями, часть которых |
||||||||||||
à |
|
|
|
|
|
á |
|
|
|
|
|
расположена дистальнее (m1, |
||||||||||
|
|
|
|
|
|
|
|
|
|
m2)илипроксимальнее(m3, m4) |
||||||||||||
+ |
+ |
|
|
ct |
v |
+ |
l |
+ |
ct + |
v |
||||||||||||
|
|
леталей (Рис. 5.10). |
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
ct |
v |
|
|
|
|
ct |
+ |
v |
|
|
|
|
Получают |
|
|
самок, |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
гетерозиготных |
ïî |
ýòèì |
||||||||
|
|
самцы |
|
|
|
|
|
самцы |
|
леталям, их скрещивают с |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
любым самцом и анализируют |
||||||||||
|
ct |
v |
43.5% |
|
|
|
ct |
v |
87.6% |
|
потомство: подсчитывают |
|||||||||||
|
+ |
+ |
43.5% |
|
|
|
+ |
+ |
0.4% |
|
||||||||||||
|
|
|
|
|
число |
самок, а самцы- |
||||||||||||||||
|
ct |
+ |
6.5% |
|
|
|
ct |
+ |
7.5% |
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
+ |
v |
6.5% |
|
|
|
+ |
v |
4.5% |
|
некроссоверы все гибнут, т.к. |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
обе Х-хромосомы у самки- |
||||||||||
|
|
|
+ |
l |
v |
+ |
|
+ |
v |
|
|
материнесутлетали.Выживают |
||||||||||
|
|
|
ct |
+ |
+ |
ct |
l |
+ |
|
|
только кроссоверы + + + + + +, |
|||||||||||
|
|
|
|
|
ò.å. |
åñëè |
|
кроссинговер |
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
кроссоверы ct + |
кроссоверы + |
v |
|
произошел между леталями и |
||||||||||||||||
|
|
|
â |
|
|
|
|
|
|
|
|
эти летали расположены в том |
||||||||||
|
|
|
ct |
|
l |
|
|
|
v |
|
|
порядке, |
â |
котором |
îíè |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
22.0 |
|
|
|
|
33.0 |
|
|
расположены |
íà |
âûøå |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
4.1 |
|
6.9 |
|
|
|
|
|
приведеннойсхеме(Рис.5.10а). |
||||||||||
|
|
|
|
|
|
|
|
|
|
Åñëè l2 расположена левее чем |
||||||||||||
Схемы скрещивания для картирования летальной мутации в |
||||||||||||||||||||||
l ,выживаюткроссоверы m m |
||||||||||||||||||||||
Х-хромосоме дрозофилы. |
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
1 |
2 |
||||||
|
|
|
|
|
|
+ + m3 m4 (Ðèñ. 5.10á). |
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
промежутке, который нужно поделить в |
Расстояние |
|
между |
леталями |
||||||||||||||||||
соответствии с частотами кроссинговера |
определяетсякакотношениеудвоенногочисла |
|||||||||||||||||||||
4,5:12 и 7,5:12, то есть леталь будет |
выжившихсамцовкчислусамоквпотомстве |
|||||||||||||||||||||
располагатьсяв4,125морганидыот ct и6,875 |
(Èç: Lefevre, 1971; Zhimulev et al., 1981). |
|
||||||||||||||||||||
– îò v. (ñì. Ðèñ. 5.9â). |
|
|
|
|
|
5.6.4. Соотношение |
|
|
|
|
|
|||||||||||
|
Для окончательного картирования |
|
|
|
|
|
||||||||||||||||
леталей следует использовать делеции и |
кроссоверной и молекулярной |
|
||||||||||||||||||||
дупликации(см.Рис.4.15). |
|
|
|
|
карт генов |
|
|
|
|
|
|
|
|
|
||||||||
5.6.3. Селективные схемы |
|
|
|
Учитывая |
òî, |
÷òî |
|
íà |
частоту |
|||||||||||||
|
|
|
кроссинговера влияют разнообразные |
|||||||||||||||||||
скрещивания |
|
|
|
|
|
|
||||||||||||||||
|
|
|
|
|
|
факторы,всегдаостаетсяощущениетого,что |
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
В некоторых случаях особенности |
кроссоверныерасстоянияотносительныиони |
||||||||||||||||||||
организации гена, участка хромосомы или |
не имеют соответствия каким-то реальным |
|||||||||||||||||||||
самойхромосомыпозволяютсоставитьсхемы |
Рисунок 5.10 |
|
|
|
|
|
|
|
|
|
||||||||||||
скрещиваний, позволяющие анализировать |
|
|
|
|
|
|
|
|
|
|||||||||||||
m1 m2 l1 |
+ + + |
|
|
|
|
|
|
|||||||||||||||
огромныевыборкипотомков,врезультатечего |
|
|
+ + + + + + |
|
||||||||||||||||||
à |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
появляетсявозможностькартироватьмутации |
+ |
+ |
+ |
l2 |
m3 |
m4 |
|
|
|
|
|
|
||||||||||
наоченькороткихрасстоянияхидажевнутри |
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
гена.Известнысхемы,позволяющиенадежно |
m1 |
m2 + |
l1 |
+ |
+ |
|
|
+ + + + + + |
|
|||||||||||||
картировать взаимное расположение генов, |
á |
|
|
|
|
|
|
|
|
|
|
|
||||||||||
+ |
+ |
l2 |
+ m3 |
m4 |
|
|
|
|
|
|
||||||||||||
лежащих рядом и определять расстояние |
|
|
|
|
|
|
||||||||||||||||
Селективная схема скрещиваний с помощью |
|
|||||||||||||||||||||
междуними,например,картированиелеталей, |
|
|||||||||||||||||||||
которой картируют близко расположенные |
|
|||||||||||||||||||||
локализованных очень близко друг к другу в |
|
|||||||||||||||||||||
летали. |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
Х-хромосомедрозофилы.Вэтомслучаелетали |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
а. Леталь l1 расположена слева от летали l2. |
|
|||||||||||||||||||||
(l1-l2)маркируютсярецессивнымивидимыми |
|
|||||||||||||||||||||
б. Леталь l расположена правее чем леталь l . |
|
|||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
89 |
|
|

Глава 5 |
|
|
|
|
|
|
|
Генетический анализ |
|
|||
|
|
|
|
|
|
|
||||||
физическим единицам, например, числу |
5.6.6. Картирование генов с |
|
||||||||||
нуклеотидов ДНК. Обычно, чтобы получить |
помощью соматического |
|||||||||||
данные о соотношении между единицей |
кроссинговера |
|
|
|||||||||
кроссинговера и числом нуклеотидов в ДНК, |
Порядок генов в хромосоме можно |
|||||||||||
значениеразмерагеномаделятнаобщуюдлину |
||||||||||||
определить и с помощью митотического |
||||||||||||
кроссовернойкарты(Табл.5.1). |
|
|
кроссинговера. Однако, |
расстояния |
||||||||
|
Данные Таблицы5.1позволяютсделать |
|||||||||||
следующиезаключения: |
|
|
|
получаютсясовершенноиные(Рис.5.11). |
||||||||
|
|
|
|
|
|
|
|
|||||
1. Частота генетической рекомбинации |
Литература к разделам 5.1. - 5.6. |
|||||||||||
значительно выше у прокариот и низших |
||||||||||||
эукариот,чемуэукариот.Так,дрожжиимеют |
Гершензон С.М. Основы современной генетики. |
|||||||||||
кроссовернуюкартуразмером4200единиц,в |
Киев, Наукова Думка, 135-169, 1983. |
|
||||||||||
Захаров И.А. Генетические карты высших |
||||||||||||
то время, как дрозофила или нематода |
||||||||||||
(представитель круглых червей) - 280 и 320, |
организмов. Ленинград, Наука, 1-157, 1979. |
|||||||||||
Инге-Вечтомов С.Г. Генетика с основами |
||||||||||||
соответственно. |
|
|
|
|
|
селекции. Москва, Высшая школа, 144-169, |
||||||
2. Как следствие, у прокариот число |
||||||||||||
1989. |
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
||
нуклеотидов на единицу карты значительно |
Кушев В.В. |
|
Механизмы |
генетической |
||||||||
ниже,чемуэукариот. |
|
|
|
рекомбинации. Ленинград, Наука, 1-247, |
||||||||
|
|
|
|
|
|
|
1971. |
|
|
|
|
|
5.6.5. Картирование генов с |
|
Лобашев М.Е. Генетика. Ленинград, Изд-во ЛГУ, |
||||||||||
помощью хромосомных |
|
226-257, 1967. |
|
|
||||||||
перестроек |
|
|
|
|
Серебровский А.С. Генетический анализ. |
|||||||
|
|
|
|
Москва, Наука, 1-342, 1970. |
|
|||||||
|
Мутации лучше всего картировать, |
|
||||||||||
|
Тихомирова М.М. Генетический анализ. |
|||||||||||
используя наборы делеций и дупликаций. |
Ленинград, Изд-во Ленинградского |
|||||||||||
|
|
|
|
|
|
|
||||||
Картирование гена с помощью делеций |
Университета, 1-280, 1990. |
|
||||||||||
изображено на Рис. 4.14-4.16. С помощью |
Финчем Дж. Генетическая комплементация. |
|||||||||||
транслокаций (изменяется сцепленность |
Москва, Мир, 1-183, 1968. |
|
|
|||||||||
наследования) можно определять группы |
Ashburner M. Drosophila. A laboratory handbook. |
|||||||||||
сцепления (см. Раздел 4.3.2.). |
|
Cold Spring Harbor, Cold Spring Harbor Press, |
||||||||||
|
912, 1989. |
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
Judd B.H, Shen M.W., Kaufman T.C. The anatomy |
|||||
|
|
|
|
|
|
|
andfunctionofasegmentoftheX chromosome |
|||||
Таблица 5.1 |
|
|
|
|
|
|
|
|
|
|
||
|
Соотношение кроссоверных и молекулярных карт (Из: Lewin, 1994, стр. 128). |
|
|
|||||||||
Âèäû |
|
Размер генома в парах |
|
Число кроссоверных |
|
Размер кроссоверной единицы |
|
|||||
|
|
|
|
|||||||||
|
нуклеотидов (а) |
|
единиц карты в геноме (б) |
|
в парах нуклеотидов (а/б) |
|
||||||
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
||
|
Ôàã T4 |
|
1.6 × |
105 |
|
|
800 |
|
200 |
|
||
|
|
|
|
|
|
|
|
|
|
|
||
|
E. coli |
|
4.2 × |
106 |
|
|
1750 |
|
2400 |
|
||
|
|
|
|
|
|
|
|
|
|
|
||
Дрожжи |
|
2.0 × |
107 |
|
|
4200 |
|
5000 |
|
|||
|
|
|
|
|
|
|
|
|
|
|
||
Грибы |
|
2.7 × |
107 |
|
|
1000 |
|
27000 |
|
|||
|
|
|
|
|
|
|
|
|
|
|
||
Нематоды |
|
8.0 × |
107 |
|
|
320 |
|
250000 |
|
|||
|
|
|
|
|
|
|
|
|
|
|
||
Дрозофила |
|
1.4 × |
108 |
|
|
280 |
|
500000 |
|
|||
|
|
|
|
|
|
|
|
|
|
|
||
|
Ìûøü |
|
3.0 × |
109 |
|
|
1700 |
|
1800000 |
|
||
|
|
|
|
|
|
|
|
|
|
|
||
Человек (муж.) |
|
3.3 × |
109 |
|
|
2809 |
|
1200000 |
|
|||
|
|
|
|
|
|
|
|
|
|
|
||
Человек (жен.) |
|
3.3 × |
109 |
|
|
4782 |
|
700000 |
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
90

Генетический анализ |
Глава 5 |
Рисунок 5.11 |
|
Сопоставление расстояний между генами третьей хромосомы дрозофилы, выявленных с |
|
помощью мейотического и митотического кроссинговера (Из: Ashburner, 1989, p. 912). |
|
of Drosophila melanogaster.Genetics % : 139156, 1972.
Lefevre G. Jr. Salivary chromosome bands and the frequency of crossing over in Drosophila melanogaster. Genetics $%: 497-513, 1971.
LindsleyD.L.,ZimmG.G.ThegenomeofDrosophila melanogaster.AcademicPress,SanDiego,New York, Boston, London, 1-1133, 1992.
O’Brien S. Genetic maps. Cold Spring Harbor, Cold Spring Harbor Laboratory Press, vol. 2, 1-405, 1982.
ZhimulevI.F.,PokholkovaG.V.,BgatovA.V.,Semeshin V.F., Belyaeva E.S. Fine cytological analysis of theband10A1-2andtheadjoiningregionsinthe
Drosophila melanogaster X chromosome. II.
Geneticalanalysis.Chromosoma& :25-40,1981.
5.7. Метод анеуплоидных тесторов
Анеуплоидные особи - это особи, геномы которых не содержат какое-то число хромосом. Если недостает двух гомологичных хромосом - такие особи называют нуллисомиками, одной гомологичной хромосомы - моносомиками. Особей с одной лишней гомологичной хромосомой называют трисомиками (см. раздел 4.4.4).
5.7.1. Нуллисомия
Если скрещивание линии, содержащей картируемуюмутацию r, с тесторной линией нулли-А дает в F1 нормальных потомков,
P rr  0A0A
0A0A
F1 Дикий тип
то в этом случае мутация r не картируется в хромосоме А и нужно взять следующую линию-тестор, например,нулли-В.
P |
rr |
|
0B0B |
|
|||
|
|
|
|
F1 Мутантный фенотип r
Во этом скрещивании проявляется мутантный фенотип r, значит, мутация картирована в хромосоме B. Полностью генотипы для первого скрещивания записываются следующим образом:
P AABrBr 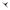 0A0ABRBR
0A0ABRBR
F1 A0ABRBr
Мутация не проявляется
Для второго скрещивания:
P AABrBr  AA0B0B
AA0B0B
F1 AABr0B
Проявляется мутантный фенотип r
91