

Столбиками обозначен объем секреции сока. Кривая показывает степень свободное соляной кислоты желудочного сока
людях Н. И. Лепорский и Е. А. Нечаева показали,что кровь одного человека, взятая в разгар секреции, вызванной натуральными условными раздражителями, и перелитая другому человеку, обладает свойством возбуждать у последнего желудочную секрецию (рис. 27). Возможно, что усиленное образование ацетилхолинопо-добных веществ в этих случаях происходит преимущественно в центральных ин-первационных приборах, хотя не исключено и образование их на периферии, в частности в желудке.
Регуляция секреторного процесса обусловливается пищевым центром, расположенным в различных отделах головного мозга. Высший регуляторный орган — кора больших полушарий, которая оказывает влияние на работу желудочных желез по типу «пускового» и «корригирующего» механизмов, т. е. вызывая активность желез при их физиологическом покое («пусковой» механизм влияний).
В возникновении секреторной реакции большую роль играет исходная возбудимость желудочных желез, которая определяется функциональным состоянием пищевого центра.
Передача возбуждения с рецепторных полей на желудочные железы осуществляется рефлекторным путем по нервам при участии медиаторов (аце-тилхолина и адреналина). Секреторный процесс возникает вначале в железах малой кривизны, а затем в железах других секреторных полей желудка. Величина латентного периода возбуждения желудочных желез на малой кривизне равняется 1—3 мин, а на большой кривизне — 5—10 мин, если судить по первой кислой капле, выделившейся через фистулу наружу. На самом деле, как показали последние электрофизиологические исследования, он исчисляется от 7 до 20 с,
93


 Рефлекторное
возбуждение желудочных желез легко
тормозится при воздействиях на высшие
отделы центральной
нервной системы и при раздражениях
периферических
нервов. В механизме торможения секреторного
процесса
принимают участие и инкреты желез
внутренней
секреции (адреналин и питуитрин), а также
специфические гормоны, образующиеся
в стенке кишечника (энтерогастрон);
тормозящее действие их на секреторные
клетки
осуществляется через нервную систему.
Рефлекторное
возбуждение желудочных желез легко
тормозится при воздействиях на высшие
отделы центральной
нервной системы и при раздражениях
периферических
нервов. В механизме торможения секреторного
процесса
принимают участие и инкреты желез
внутренней
секреции (адреналин и питуитрин), а также
специфические гормоны, образующиеся
в стенке кишечника (энтерогастрон);
тормозящее действие их на секреторные
клетки
осуществляется через нервную систему.
Желудочная секреция, возникающая при механическом раздражении рецепторов желудка, — сложнореф-лекторная реакция, в которой принимает участие кора больших полушарий головного мозга. «Механическая секреция» возникает вначале в секреторном поле малой кривизны, а затем в других секреторных полях желудка. У человека она характеризуется коротким, по сравнению с животными, латентным периодом и обильным отделением сока в первые 15—30 мин непрерывного действия раздражителя. Количество отделяемого сока зависит, с одной стороны, от функционального состояния высших отделов центральной нервной системы и степени возбудимости нервно-железистого аппарата желудка, с другой — от силы и времени действия раздражителя.
Возбуждение фундальных желез при механическом раздражении стенок дна и тела желудка осуществляется при помощи рефлекторного механизма. Рефлекторная передача возбуждения с механорецепторов желудка на секреторные клетки происходит через блуждающие нервы.
В процессе возбуждения желудочных желез человека большую роль играет кора головного мозга — при ее участии происходит образование условного интероцеп-тивного рефлекса. Поскольку у людей во время сна «механическая секреция» бывает меньше, чем во время бодрствования, то, следовательно, механизм возникновения ее у человека осуществляется помимо условного интероцептивного рефлекса и в порядке безусловнореф-лекторной реакции (рис. 28).
Нервно-химическая фаза желудочной секреции возникает вслед за сложнорефлекторной и обусловлена возбуждающим действием специфических гормонов, продуктов расщепления пищевых веществ и экстрактивных, содержащихся в мясе, рыбе и овощах и появ-
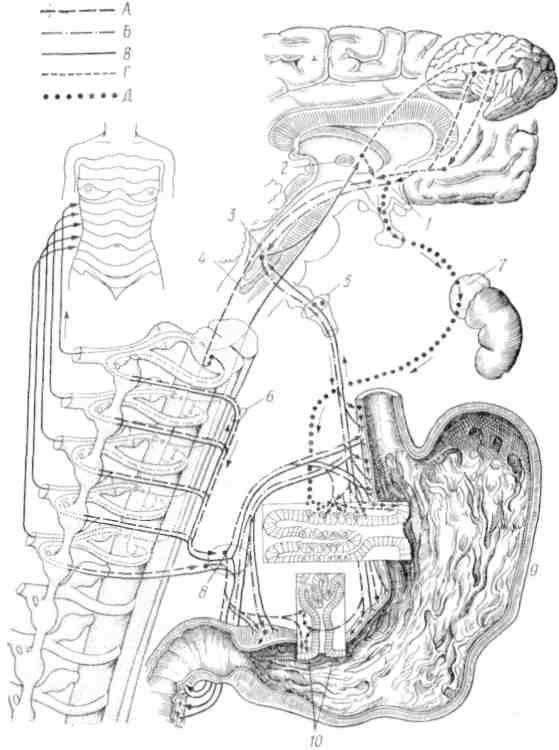
Рис. 28. Нейрогуморальная регуляция функций желудка (по Нет-тер, 1959). А — симпатические эфферентные волокна, Б — парасимпатические эфферентные волокна, В — афферентные волокна, Г— неизвестные пути, Д — гуморальные связи:
/ — гипоталамус; 2 — таламус; 3 — дорзальпые ядра вагуса; 4 — tr. solita-rius; 5 — блуждающий нерв; 6 — чревный нерв; 7 — кора надпочечников; 8— gangl. coeliacum; 9 — фуидальные железы; 10 — пилорические железы


 ляющихся
при варке супов. Основанием для признания
наличия второй фазы желудочной секреции
послужили опыты на собаках с частично
(по Гейденгайну) или полностью (по
Грегори) денервированными изолированными
желудочками. Под влиянием вводимых в
полость большого желудка экстрактивных
веществ в изолированном желудочке
начиналось отделение кислого желудочного
сока. В то же время акт еды подобной
реакции не вызывал. Изучение механизма
развития секреторного процесса в
эту фазу дало много ценных сведений.
Однако до сих пор механизм работы
желудочных желез еще недостаточно ясен
и по этому поводу существует несколько
теоретических воззрений.
ляющихся
при варке супов. Основанием для признания
наличия второй фазы желудочной секреции
послужили опыты на собаках с частично
(по Гейденгайну) или полностью (по
Грегори) денервированными изолированными
желудочками. Под влиянием вводимых в
полость большого желудка экстрактивных
веществ в изолированном желудочке
начиналось отделение кислого желудочного
сока. В то же время акт еды подобной
реакции не вызывал. Изучение механизма
развития секреторного процесса в
эту фазу дало много ценных сведений.
Однако до сих пор механизм работы
желудочных желез еще недостаточно ясен
и по этому поводу существует несколько
теоретических воззрений.
Теория И. П. Павлова. И. П. Павлов утверждал, что «система организма, его бесчисленных частей соединяется в единое целое двояким образом: посредством специфической ткани, которая существует только для поддержания взаимных отношений, а именно нервной ткани, и при помощи тканевых жидкостей, омывающих все тканевые элементы. Эти же самые посредники переносят также и наши раздражители на железистую ткань» .
Механизм второй фазы желудочной секреции И. П. Павлов рассматривал как нервно-химическую. Об этом свидетельствует ряд фактов. Например, введение атропина в разгаре пищеварения полностью прекращает секрецию желудочного сока. Подобный эффект наблюдается и в тех случаях, когда перед введением химических возбудителей в привратник производилась предварительная кокаинизация слизистой оболочки последнего. Установлено также, что у людей и животных после двусторонней перерезки блуждающих нервов значительно уменьшается секреторный эффект на химические возбудители и т. д.
Теория И. С. Эдкинса. Ряд физиологов, стоявших на позициях «гуморализма», рассматривает вторую фазу желудочной секреции как чисто гуморальную, не связанную с нервной системой. Такого же взгляда придерживается американский ученый Эдкинс, выдвинувший теорию гормонального возбуждения желудочных желез. Согласно этой теории возбуждение фундаль-
1 Павлов И. П. Поли. собр. соч., т. II, кн. 2. М., 1951, с. 355—356.
96
ных желез во второй фазе обусловливается действием специального гормона гастрина, который под влиянием химических веществ образуется в стенке пилорического отдела желудка из прогормона и кровяным током подносится к фундальным железам. В опытах было показано, что кровь, взятая у собаки с удаленным привратником в разгаре пищеварения и перелитая другой собаке, не вызывает секреции желудочного сока, в то время как кровь собаки с сохраненным пилорическим отделом желудка всегда вызывает секрецию. Ряд авторов утверждает, что в привратнике образуются гормоны иило-рин и пилороантрумгастрин, которые гуморальным путем возбуждают фундальные железы.
Теория Л. Б. Попельского. Теория гормонального механизма возбуждения желудочных желез получила дальнейшее развитие в исследованиях Л. Б. Попельского, установившего, что в процессе возбуждения желез принимает активное участие гистамип, образующийся в желудке. Является ли гистамин действующим началом гастрина Эдкинса, как это считал По-пельский и другие исследователи, или он представляет самостоятельный гормон в настоящее время точно не установлено. По мнению некоторых авторов, гистамин никакого отношения к гастрину не имеет. Возможно, гистамин— медиатор, выделяющийся при раздражении блуждающих нервов.
Эндокринные теории. Кроме гастрина, пило-рина, пилороантрумгастрина и гистамина в последнее время были обнаружены в слизистой оболочке верхних и средних отделов тонкого кишечника собак особые гормоны, обладающие способностью возбуждать желудочную секрецию («желудочные секретины»). Помимо названных гормонов, в желудке был недавно найден еще один гормон, обозначенный субстанцией «Р», который также обладает свойством возбуждать желудочные клетки. В механизме желудочной секреции принимают участие и такие гормоны желез внутренней секреции, как инсулин, адреналин, паратиреоидин и гидрокортизон. Особая роль в этом отношении принадлежит инсулину, который возбуждает желудочные железы через систему блуждающих нервов, и адренокортикотропному гормону (АКТГ), который стимулирует образование соляной кислоты желудочного сока через кортикостероид-ные гормоны надпочечника.
97

 Теория
С. Окада.
Участие инсулина и адреналина в
регуляции концентрации сахара крови
дало основание японскому ученому
Окада и некоторым другим исследователям
высказать теорию о механизме регуляции
деятельности желудочных желез. Согласно
этой теории при повышении уровня сахара
крови (гипергликемия) угнетаются, а
при понижении уровня сахара крови
(гипогликемия) возбуждаются секреторные
клетки желудка. Однако эта теория
встретила серьезные возражения.
Вероятно, участие сахара крови в
процессе возбуждения желудочных
желез следует рассматривать в связи
с общей нервно-гуморальной регуляцией,
осуществляемой пищевым центром.
Теория
С. Окада.
Участие инсулина и адреналина в
регуляции концентрации сахара крови
дало основание японскому ученому
Окада и некоторым другим исследователям
высказать теорию о механизме регуляции
деятельности желудочных желез. Согласно
этой теории при повышении уровня сахара
крови (гипергликемия) угнетаются, а
при понижении уровня сахара крови
(гипогликемия) возбуждаются секреторные
клетки желудка. Однако эта теория
встретила серьезные возражения.
Вероятно, участие сахара крови в
процессе возбуждения желудочных
желез следует рассматривать в связи
с общей нервно-гуморальной регуляцией,
осуществляемой пищевым центром.
Теория И. П. Разенкова. И. П. Разенков в 1925 г. выдвинул теорию чисто химического возбуждения желудочных желез. Согласно ей механизм второй Фазы желудочной секреции не гуморальный, с точки зрения специфического гормона, а чисто химический, поскольку химические возбудители, т. е. продукты переваривания пищи, попав в круг кровообращения, непосредственно сами возбуждают работу желудочных желез. Проведенный анализ «сытой» и «голодной» крови показал существенную разницу биохимических свойств той и другой. На собаках с удаленным привратником было установлено, что пищевые и химические вещества (молоко, либиховский экстракт, гемоза, соляная кислота и др.), введенные в тонкую кишку, возбуждают деятельность желудочных желез.
Подобный эффект наблюдается и у больных с удаленным привратником.
В 1948 г. И. П. Разенков пришел к заключению, что вторая фаза секреторной деятельности желудочных желез обусловливается не только действием составных частей пищи и продуктов ее переваривания, но и специфически гормональными веществами, образующимися в слизистой оболочке пилорической части желудка или кишечника.
Итак, вторая фаза желудочной секреции является очень сложной; в ней принимают участие, помимо рефлекторных рлияний различных веществ с привратника и двенадцатиперстной кишки, специфические вещества гормонального происхождения (гастрин, пилорин, пило-роантрумгастрин, гистамин, субстанция «Р» и т. д.) и продукты переваривания самой пищи, которые через
кровь поступают в желудок и возбуждают его секреторные клетки.
Объяснить механизм возбуждения желудочных желез во вторую фазу секреции какой-либо одной из предложенных гуморальных теорий, с точки зрения современных представлений, основанных на принципах павловского «нервизма», невозможно.
Установленные И. П. Павловым и его сотрудниками, а позднее А. В. Соловьевым, Е. М. Матросовой, А. Н. Ба-курадзе факты о зависимости работы желудочных желез во вторую фазу секреции от нервной системы (симпатической и парасимпатической) дают право считать, что в механизме возбуждения желудочных желез гормонами и химическими веществами принимает участие нервная система. Если еще принять во внимание, что в механизме возбуждения желудочных желез во второй фазе их деятельности принимают большое участие и медиаторы, то станет вполне очевидным, что механизм возбуждения желез желудка во вторую фазу является нервно-гуморальным. Такой взгляд полностью отражает принципиальное положение И. П. Павлова о единстве нервного и гуморального механизмов регуляции при ведущем значении нервного механизма.