

Структурно-функциональная характеристика органелл, составляющих цитоскелет клеток.
ЦИТОСКЕЛЕТ
Трёхмерная цитоплазматическая сеть волокнистых и трубчатых структур различного типа формирует цитоскелет. К элементам цитоскелета относят микротрубочки, промежуточные филаменты, микрофиламенты. Цитоскелет придаётклетке определённую форму и выполняет множество других функций (например, подвижность клетки, внутриклеточный транспорт).
Микротрубочки
Микротрубочки состоят из 13 тубулиновых протофиламентов (нитей), идущих о спирали; нити имеют диаметр 24 нм и длину несколько мкм
Микротрубочки входят в состав аксонемы. базального тельца и центриоли. Микротрубочки участвуют в поддержании формы клетки в транспорте макромолекул и органелл, обеспечивают расхождения хромосом при делении
клеток подвижность жгутиков и ресничек. Для микротрубочек характерна полярность.
Полярность микротрубочек. Микротрубочки — динамичные структуры постоянно растущие с одного конца (полимеризация) и деполимеризующиеся с другого конца. В каждой микротрубочке различают (+)-конец где присоединяются новые СЕ тубулиновых протофиламентов, и (-)-конец где СЕ тубулина отделяются от нити.
Промежуточные нити
Промежуточные нити (филаменты) состоят из белков, специфичных для определённых клеточных типов; имеют диаметр 8—11 нм.). Промежуточные нити создают внутриклеточный каркас, обеспечивают упругость клетки, поддерживают упорядоченность расположения компонентов цитоплазмы. Иммуноцитохимические реакции с АТ конкретных типов промежуточных нитей нашли применение в цитодиагностике генеза опухолей.
Микрофиламенты
Две переплетённые нити Р-актина, составленные из 0-актина. Формируют
микрофиламенты диаметром 6 нм. Микрофиламенты образуют скопления по
периферии клетки и связаны с плазмолеммой посредством белков (кактинин,
винкулин, талин).
Полярность микрофиламентов. Как и микротрубочки, микрофиламенты по-
лярны; присоединение (полимеризация) СЕ 0-актина происходит на (+) конце.
Известны токсины, связывающиеся с актином и блокирующие его полимеризацию, нарушая тем самым подвижность клеток, фагоцитоз и цитокинез. К таким относятся цитохалазины и фаллоидин.
Функции микрофиламентов
Изменение консистенции цитозоля, переход золя в гель и обратно (например, для изменения вязкости примембранной цитоплазмы при образовании псевдоподии).
Строение и значение центриолей, ресничек и жгутиков.
Органеллы, содержащие триплеты микротрубочек (центриоля и базальное тельце) или аксонему (реснички и жгутики), участвуют в расхождении хромосом, мерцании ресничек, движении сперматозоидов.
Центриоли
Центриоли (см. рис. 2-8) между клеточными делениями расположены вблизи ядра, часто рядом с комплексом Гольджи. Центриоль имеет цилиндрическую форму, диаметр 150 им и длину до 500 нм; стенка состоит из 9 триплетов микротрубочек. Центриоль — центр организации митотического веретена — участвует в делении клетки. В ходе фазы 5 клеточного цикла центриоли дуплицируются. Образовавшаяся центриоль расположена под прямым углом к первоначальной. При митозе пары центриолей, каждая из которых состоит из первоначальной и вновь образованной, расходятся к полюсам клетки и участвуют в образовании митотического веретена.
Базальное тельце
Базальное тельце состоит из 9 триплетов микротрубочек, расположенных в основании реснички или жгутика; служит матрицей при организации аксонемы.
Аксонема
Аксонема формируется путём самосборки. Матрицей для сборки служит центриоль или базальное тельце. Аксонема состоит из 9 периферических
Пар микротрубочек и двух расположенных центрально одиночных микротрубочек. В каждой периферической паре различают субфибриллу А, содержащую 10—11 тубулиновых протофиламентов, и субфибриллу В, содержащую 13 протофиламентов.
Ресничка — вырост клетки длиной 5—10 мкм и шириной 0,2 мкм, содержащий аксонему. Реснички присутствуют в эпителиальных клетках воздухопроводящих и половых путей, перемещают слизь с инородными частицами и остатками отмерших клеток и создают ток жидкости около клеточной поверхности.
Жгутик как правило, не встречается в количестве более двух на клетку. В сперматозоиде человека имеет длину 50-55 мкм и толшину 0,2—0,5 мкм, содержит аксонему.
9. Основные положения клеточной теории. Вклад Пуркенье, Шванна, Вирхова и др. ученых о клетке. Определение клетки. Включения цитоплазмы: понятие, классификация, химическая и морфофункциональная характеристика.
Современная клеточная теория включает такие положения;
1. Клетка является наименьшей единицей живого.
2. Клетки разных организмов имеют похожее строение.
3. Размножение клеток происходит путем деления материнской клетки
(omnia cellula e cellula — каждая клетка — из клетки).
4. Многоклеточные организмы состоят из сложных ансамблей клеток
и их производных.
Значение клеточной теории состоит в следующем:
1. Она явилась фундаментом для развития многих биологических дисциплин, прежде всего цитологии, гистологии, эмбриологии, физиологии, а также патологии.
2. Позволила понять механизмы онтогенеза — индивидуального развития организмов.
3. Явилась основой для материалистического понимания жизни, окружающего мира.
4. Явилась основой для объяснения эволюции организмов.
Клетка — это элементарная структурная единица организма, состоящая из ядра, цитоплазмы и ограниченная клеточной оболочкой, способная выполнять все функции, характерные живому: обмен веществ и энергии, размножение, рост, раздражимость, сократимость, хранение генетической информации и ее передачу.
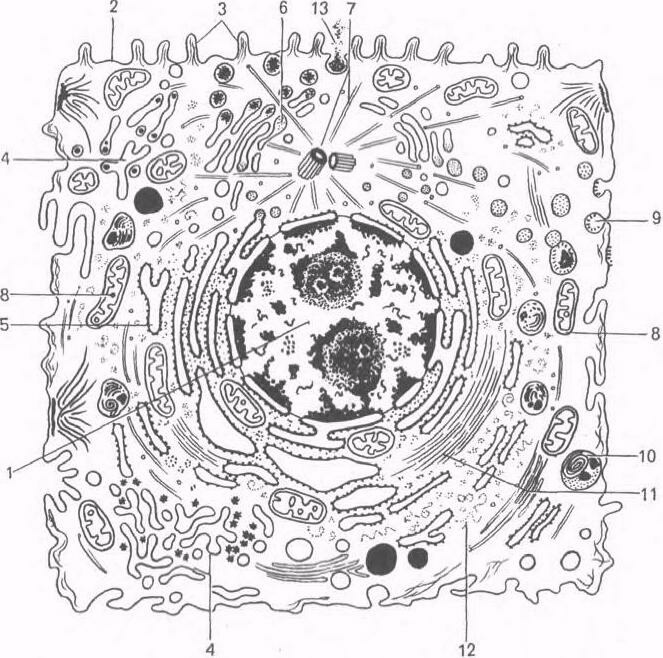
Ультрамикроскопическое строение клетки животных организмов (схема).
1 — ядро; 2 — плазмолемма; 3 — микроворсинки; 4 — агранулярная эндоплазматическая сеть; 5 — гранулярная эндоплазматическая сеть; 6 — аппарат Гольджи; 7 — центриоль и микротрубочки клеточного центра; 8 — митохондрии; 9 — цитоплазматические пузырьки, 10 — лизосомы; 11 — микрофиламенты; 12 — рибосомы; 13 — выделение гранул секрета.
ВКЛЮЧЕНИЯ
Включения — это непостоянные компоненты клетки, не имеющие строго постоянной структуры (их структура может меняться). Выявляются в клетке только в определенные периоды жизнедеятельности или жизненного цикла.
КЛАССИФИКАЦИЯ ВКЛЮЧЕНИЙ.
1. Трофические включения представляют собой депонированные питательные вещества. К таким включениям относятся, например, включения гликогена, жира.
2. Пигментные включения. Примером таких включений являются гемоглобин в эритроцитах, меланин в меланоцитах. В некоторых клетках (нервные, печеночные, кардиомиоциты) при старении в лизосомах накапливается пигмент старения коричневого цвета липофусцин, не несущий, как полагают, определенной функции и образующийся в результате изнашивания клеточных структур. Следовательно, пигментации включения представляют собой химически, структурно и функционально неоднородную группу. Гемоглобин участвует в транспорте газов, меланин выполняет защитную функцию, а липофусцин является конечным продуктом обмена. Пигментные включения, за исключением липофусциновых, не окружены мембраной.
3. Секреторные включения выявляются в секреторных клетках и состоят из продуктов представляющих собой биологически активные вещества и другие необходимые для осуществлении функций организма вещества (включения белка, в том числе и ферментов, слизистые включения в бокаловидных клетках и др.). Эти включения имею вид окруженных мембраной пузырьков, из которых секретируемый продукт может иметь различную электронную плотность и часто окружен светлым бесструктурным ободком.
4. Экскреторные включения - включения, подлежащие выведению из клетки, поскольку состоят из конечных продуктов обмена. Примером являются включения мочевины в клетках почки и т.д. По структуре похожи на секреторные включения.
5. Специальные включения — фагоцитированные частицы (фагосомы), поступающие в клетку путем эндоцитоза.
10. Ядро, характеристика как генетического центра, роль ядра в хранении и передачи наследственной информации и в синтезе белка. Общий план строения неделящего ядра: хроматин, ядрышко, ядерная оболочка, нуклеоплазма.
ЯДРО
Ядро клетки — структура, обеспечивающая генетическую детерминацию и регуляцию белкового синтеза.
23 пары хромосом, ядра соматической клетки содержат ядерный геном. Реализация генетической информации (транскрипция, процессинг, трансляция, посттрансляционная модификация) и другие функции ядра происходят при участии ДНК и разных видов РНК. Структурные компоненты ядра: хроматин, ядерная оболочка, ядрышко, нуклеоплазма.
ХРОМАТИН
Термином «хроматин» обозначают комплекс ядерной ДНК с белками (гистоны, негистоновые). Различают гетеро- и эухроматин.
Гетерохроматин. Гетерохроматин — транскрипционно неактивный, конденсированный хроматин интерфазного ядра. В СМ — базофильные глыбки. в ЭМ —скопления плотных гранул Располагается преимущественно по периферии ядра и вокруг ядрышек. Типичный пример гетерохроматина — тельце Барра.
Эухроматин. Эухроматин — транскрипционно активная и менее конденсированная часть хроматина, локализуется в более светлых участках ядра между гетерохроматином.
ЯДЕРНАЯ ОБОЛОЧКА
В состав ядерной оболочки входят наружная и внутренняя ядерные мембраны перинуклеарные цистерны ядерная пластинка, ядерные поры. На поверхности наружной ядерной мембраны расположены рибосомы, где синтезируются белки, поступающие в перинуклеарные цистерны, рассматриваемые как часть гранулярной эндоплазматической сети. Внутренняя ядерная мембрана отделена от содержимого ядра ядерной пластинкой.
Ядерная пластинка толщиной 80—300 нм участвует в организации ядерной
оболочки перинуклеарного хроматина, может разделять комплексы ядерных
пор и дезинтегрировать ядро в ходе митоза; содержит белки промежуточных филаментов — ламины А, В и С.
Ядерная пора имеет диаметр 80—150 нм, содержит канал поры и комплекс ядерной поры. Содержимое ядра сообщается с цитозолем через 3—4
тысячи специализированных коммуникаций — ядерных пор, осуществляющих диффузию воды, ионов и транспорт множества макромолекул (в т.ч. молекул РНК) между ядром и цитоплазмой. Перенос макромолекул через ядерные поры осуществляют специальные транспортные белки кариоферины, которые специфически распознают и связывают свои молекулы и курсируют между ядром и цитоплазмой, перенося связанную молекулу в одном направлении: из цитоплазмы в ядро (импортины) или из ядра в цитоплазму (экспортины).
ЯДРЫШКО
Ядрышко — компактная структура в ядре интерфазных клеток. В ядрышке различают фибриллярный центр слабоокрашенный компонент (ДНК, кодирующая рРНК), фибриллярный компонент,где протекают ранние стадии образования предшественников рРНК, состоит из тонких (5 нм) рибонуклеопротеиновых фибрилл и транскрипционно активных участков ДНК.
НУКЛЕОПЛАЗМА
Ядерный матрикс содержит сеть рибонуклеопротеинов, ядерные рецепторы, ферменты (АТФаза, ГТФаза, НАД-пирофосфатаза, ДНК- и РНК-полимеразы) и множество других молекул, часто образующих ассоциации — ядерные частицы. В матриксе происходят транскрипция и процессинг мРНК и рРНК. Ядерные рецепторы, онкогены, факторы транскрипции, белки теплового шока, вирусные ДНК влияют на транскрипцию и процессинг РНК.
11. Ядро: хроматин, как форма существования хромосом в неделящемся ядре. Строение и химический состав, характеристика хроматиновых фибрилл и гранул, роль белков в стуктуризации и регуляции метаболической активности хроматина. Половой хроматин. Нуклеосомы. Понятие о хромосомах. Строение, функции, классификация.
Ядро клетки — структура, обеспечивающая генетическую детерминацию и регуляцию белкового синтеза.
ХРОМАТИН
Термином «хроматин» обозначают комплекс ядерной ДНК с белками (гистоны, негистоновые). Различают гетеро- и эухроматин.
Гетерохроматин. Гетерохроматин — транскрипционно неактивный, конденсированный хроматин интерфазного ядра. В СМ — базофильные глыбки. в ЭМ —скопления плотных гранул Располагается преимущественно по периферии ядра и вокруг ядрышек. Типичный пример гетерохроматина — тельце Барра.
Эухроматин. Эухроматин — транскрипционно активная и менее конденсированная часть хроматина, локализуется в более светлых участках ядра между гетерохроматином.
Наблюдения за структурой хроматина с помощью электронного мик-роскопа показали, что как в препаратах выделенного интерфазного хроматина или выделенных митотических хромосом, так и в составе ядра на ультратонких срезах всегда видны элементарные хромосомные фибриллы толщиной 20—25 нм.
В химическом отношении фибриллы хроматина представляют собой сложные комплексы дезоксирибонуклеопротеидов (ДНП), в состав которых входят ДНК и специальные хромосомные белки — гистоновые и негистоновые. В составе хроматина обнаруживается также РНК. Количественные отношения ДНК, белка и РНК составляют 1:1,3:0,2. Обнаружено, что длина индивидуальных линейных молекул ДНК может достигнуть сотен микрометров и даже сантиметров. Среди хромосом человека самая большая первая хромосома содержит ДНК с общей длиной до 4 см. Суммарная длина молекул ДНК во всех хромосомах одной клетки человека составляет около 170 см, что соответствует 6 • 10~12 г.
В хромосомах существует множество мест независимой репликации, т.е. удвоения ДНК — репликонов. ДНК эукариотических хромосом представляют собой линейные молекулы, состоящие из тандемно (друг за другом) расположенных репликонов разного размера. Средний размер репликона около 30 мкм. В составе генома человека должно встречаться более 50 000 репликонов, участков ДНК, которые синтезируются как независимые единицы.
Синтез ДНК как на участках отдельной хромосомы, так и среди разных хромосом идет неодновременно, асинхронно. Так, например, в некоторых хромосомах человека репликация наиболее интенсивно начинается в плечах хромосом и заканчивается (при высокой интенсивности включения метки) в центромерном районе (см. ниже). Наиболее поздно репликация заканчивается в хромосомах или в их участках, находящихся в компактном, конденсированном состоянии. Таким примером может являться поздняя репликация генетически инактивированной Х-хромосомы у женщин, формирующей в ядре компактное тельце полового хроматина.
Белки хроматина составляют 60—70 % от его сухой массы. К ним относятся так называемые гистоны и негистоновые белки. Негистоновые белки составляют 20 % от количества гистонов. Гистоны — щелочные белки, обогащенные основными аминокислотами (главным образом лизином и аргинином). Очевидна структурная роль гистонов, которые не только обеспечивают специфическую укладку хромосомной ДНК, но и имеют значение в регуляции транскрипции. Гистоны расположены по длине молекулы ДНК не равномерно, а в виде блоков. В один такой блок входят 8 молекул гистонов, образуя так называемую нуклеосому. Размер нуклеосомы около 10 нм.
При образовании нуклеосом происходит компактизация, сверхспирализация ДНК, что приводит к укорачиванию длины хромосомной фибриллы примерно в 7 раз. Сама же хромосомная фибрилла имеет вид нитки бус или четок, где каждая бусина — нуклеосома. Такие фибриллы толщиной 10 нм дополнительно продольно конденсируются и образуют основную элементарную фибриллу хроматина толщиной 25 нм.
В интерфазе фибриллы хроматина образуют петли. Эти петли собраны в розетки, где основания нескольких петель связаны друг с другом негистоновыми белками ядерного матрикса. Такие петлевые группы (петлевые домены) при падении активности хроматина могут конденсироваться, уплотняться, образуя так называемые хромомеры или хромоцентры интерфазных ядер. Хромомеры выявляются также в составе митотических хромосом.
В ядрах, кроме хроматиновых участков и матрикса, обнаруживаются
перихроматиновые фибриллы, перихроматиновые и интерхроматиновые гранулы. Они содержат РНК и встречаются практически во всех активных ядрах, представляют собой информационные РНК, связанные с белками, — рибонуклеопротеиды (информосомы). Матрицами для синтеза этих РНК являются разные гены, разбросанные по де конденсированным участкам хромосомных (хроматиновых) фибрилл.
Особый тип матричной ДНК, а именно ДНК для синтеза рибосомной РНК, собран обычно в нескольких компактных участках, входящих в состав ядрышек интерфазных ядер.
ХРОМОСОМЫ.
Хромосомы видны полностью только в митозе. Наиболее удобно изучать их и метафазе (метафазные хромосомные пластинки). Основными химическими элементами хромосомы являются ДНК и белки. Комплекс ДНК с белками (в основном с гистонами) формирует фибриллярную структуру — элементарную хромосомную фибриллу, имеющую нуклеосомную организацию. Каждая нуклеосома представляет собой комплекс из 8 молекул гистонов (гистоновый октамер). Вокруг него молекула ДНК образует около 2 оборотов. Участки ДНК, связывающие соседние нуклеосомы, называются линкерной ДНК. Следующим уровнем организации хромосомы является нуклеомерная организация, или уровень хроматиновой фибриллы. В ней нуклеосомы объединяются в нуклеомеры, причем каждый нуклеомер состоит из 8--10 нуклеосом и имеет диаметр около 30 им. Хромосомы образованы хроматиноными фибриллами (хроматидами) в интерфазе. В ходе последующей упаковки нуклеомер подвергается суперспирализации и превращается в хрономер, содержащий петельные домены. Петельный домен имеет диаметр до 300 нм и соответствует одному или нескольким генам. Хромомер далее за счет суперспирализации
укорачивается, образуются конденсированные хромосомы, видимые только в митозе клетки.
МОРФОЛОГИЯ И КЛАССИФИКАЦИЯ ХРОМОСОМ
Каждая хромосома в световой микроскоп выглядит в виде палочки. У большинства хромосом можно увидеть первичную перетяжка - центромер, или кинетохор. Он делит хромосомы па два плеча. Если длина плеч одинакова, такие хромосомы называются метацентрическими. Если одно плечо больше, то хромосомы являются субметацентрическими. Хромосомы с очень коротким одним плечом называются акроцентрическими. Иногда на хромосомах есть вторичные перетяжки, отделяющие от хромосомы маленький участок - сателлит. В области вторичных перетяжек находятся ядрышковые организаторы.
Согласно Денверской классификации хромосом (Денвер, США. 1960), учитывающей размеры хромосом, расположение первичных, вторичных перетяжек и наличие спутника, все хромосомы делятся на 7 групп (А, В, С, D, E, F, G). Существует также Парижская (1971) классификация хромосом. В ее основу положена дифференциальная окраска хромосом некоторыми красителями. Эта окраска выявляет в хромосомах чередующиеся светлые и темные полосы (гетеро- и эухроматиновые районы), уникальной для каждой пары хромосом. Дифференциальная окраска позволяет достоверно отличить одну пару хромосом от другой.