

Комплементарность
Комплемента́рное (дополнительное) действие генов — это вид взаимодействия неаллельных генов,доминантные аллели которых при совместном сочетании в генотипе обусловливают новоефенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9:6:1, 9:3:4, 9:7, иногда 9:3:3:1. Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлинённую. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдёт расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлинённые.
Эпистаз
Эписта́з — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3, 7:6:3. Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена аллелей гипостатичного гена (i > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7, 13:3.
Полимерия
Полимери́я — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление F2 по фенотипу происходит в соотношении 1:4:6:4:1.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1.
Пример: цвет кожи у людей, который зависит от четырёх генов.
Билет 8
Геномные мутации: эуплоидия, автоплоидия,аллополиплоидия,аэуплоидия.
К геномным мутациям относят мутации, заключающиеся в изменении количества хромосом.Выделяюткратные гаплоидному набору (эуплоидия) и некратные гаплоидному набору (анэуплоидия) изменения числа хромосом . Эуплоидия включает гаплоидию и полиплоидию
При гаплоидии клетки содержат один хромосомный набор(каждая хромосома не имеет пары). У гаплоидных организмов рецессивные аллели всегда проявляются в фенотипе, чем объясняется их сниженная жизнеспособность. Гаплоидия известна у растений (дурман, пшеница, кукуруза). В эксперименте, подвергая икру резким колебаниям температуры, получали гаплоидных тритонов, которые отличались пониженной жизнеспособностью.
При
полиплоидии отмечается увеличенное
число хромосом, кратное гаплоидному
набору: 3n
- триплоид, 4 n - тетраплоид, 5 n - пентаплоид
и т.д. Полиплоидия
широко распространена в растительном
мире (более 1/3 всех покрытосеменных
являются полиплоидами). Ценными оказались
искусственно созданные полиплоидные
формы многих культурных растений:
триплоидный и тетраплоидный сорта
сахарной свеклы, новые полиплоидные
сорта гречихи. Экспериментальным путём
получены полиплоиды у тутового шелкопряда,
тритона, индейки, мыши, кролика.
Полиплоидизация происходит в результате:
- незавершения митоза клетки разделением цитоплазмы (митотическая полиплоидизация), в результате которого возникают тетраплоиды и т.д.;
- нерасхождения хромосом в первом делении мейоза и последующего образования гамет с диплоидным набором хромосом, а затем и триплоидной зиготы (мейотическая полиплоидизация);
- нарушения дробления зиготы (зиготическая полиплоидизация).
Митотическая и мейотическая полиплоидизации являются типамиаутополиплоидии - умножения хромосомного набора одного вида.Наряду с последней выделяют аллополиплоидию, возникающую при межвидовой гибридизации (примером аллоплоида является гибрид редьки и капусты Г.Д. Карпеченко).
Для аутоплоидов характерны нарушения онтогенеза. У мышей, например, большинство триплоидных эмбрионов погибает в первой половине беременности. У человека описаны единичные случаи рождения три- и тетраплоидных людей. Продолжительность жизни новорождённых с триплоидией варьировала от 15 минут до 7 суток. Полиплоиды человека обнаружены также при изучении выкидышей. Мозаичная диплоидно-триплоидная форма выявлялась у жизнеспособных детей в возрасте 9-10 лет.
отдельных органах (например, в печени) у человека и животных встречаются полиплоидные клетки, количество которых с возрастом увеличивается. Это явление получило название избирательной соматической полиплоидии. Оно способствует расширению функциональных возможностей органа, в случаях, когда достичь этого за счёт пролиферации клеток невозможно.
Кроме рассмотренной эуплоидии существует вторая разновидность геномных мутаций -анэуплоидия (гетероплоидия), или изменение числа хромосом, некратное гаплоидному набору (рис. 85). Например, при слиянии нормальной гаметы с гаметой, содержащей две гомологичные хромосомы, оставшиеся в клетке в результате неправильного расхождения хромосом в мейозе, возникает трисомия (рис. 86). Тетрасомия может возникнуть при слиянии двух таких «необычных» гамет.
Полимерноре взаимодействие генов.
Полимерное взаимодействие генов
Полимерия – тип взаимодействия неаллельных генов, при котором несколько пар неаллельных генов действуют на развитие одного признака. Такие гены называются полимерными (от греч. полис – много и мерос – часть). Их обозначают одной буквой, но с разными индексами, которые указывают на число аллельных пар в генотипе, обусловливающих развитие конкретного признака (А1А1А2А2).
Различают два варианта полимерии: с суммирующим действием генов и без усиления генов друг другом.
• Суммирующее
действие полимерных генов. В
этом случае степень проявления признака
зависит от числа доминантных аллелей
в генотипе особи. Так, красная окраска
зерен пшеницы определяется двумя и
более парами генов. Каждый из доминантных
генов этих аллелей определяет красную
окраску, рецессивные гены определяют
белый цвет зерен. Один доминантный
аллель дает не очень сильно окрашенные
зерна. Если в генотипе присутствуют два
доминантных аллеля, интенсивность
окраски повышается. Лишь в том случае,
когда организм оказывается гомозиготным
по всем парам рецессивных генов, зерна
не окрашены. Таким образом, при скрещивании
дигибридов происходит расщепление в
отношении 15 окрашенных к одному белому.
Но из 15 окрашенных один будет иметь
интенсивный красный цвет, т.к. содержит
четыре доминантных аллеля, четыре будут
окрашены несколько светлее, т.к. содержат
в генотипах три доминантных аллеля и
один рецессивный аллель, шесть – еще
светлее с двумя доминантными и двумя
рецессивными аллелями в генотипах,
четыре – еще более светлые, т.к. имеют
лишь один доминантный и три рецессивных
гена, то есть истинное расщепление будет
1:4:6:4:1.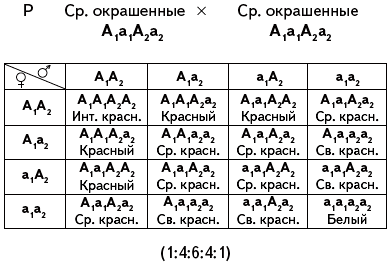
По данному варианту полимерии наследуются окраска кожи, рост и масса у человека. Подобный же механизм наследования характерен для многих количественных, в том числе и хозяйственно-ценных признаков: содержание сахара в корнеплодах свеклы, содержание витаминов в плодах и овощах, длина колоса злаков, длина початка кукурузы, плодовитость животных, молочность скота, яйценоскость кур и др.
• Полимерные гены не усиливают друг друга. В этом случае расщепление в F2 будет 15:1. Так, плоды пастушьей сумки могут быть треугольными (доминантный признак) и овальными (рецессив). Признак контролируется двумя парами полимерных генов. Если в генотипе растения имеется хотя бы один доминантный аллель из первой или второй пары полимерных генов, то форма плода у него будет треугольной (А1А2; А1а2;а1А2). Овальную форму плодов будут иметь лишь те растения, у которых в генотипе нет ни одного доминантного аллеля – а1а1а2а2.
Таким образом, накопление определенных аллелей в генотипе может привести к изменению выраженности признаков.
Билет 9
Хромосомные перестройки ( аберрации)
Хромосомные аберрации (хромосомные мутации, хромосомные перестройки) — изменения структуры хромосом. Классифицируют делеции (удаление участка хромосомы), инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы на другую). Хромосомные перестройки носят, как правило, патологический характер и нередко приводят к гибели организма. Показано значение хромосомных перестроек в видообразовании и эволюции.