

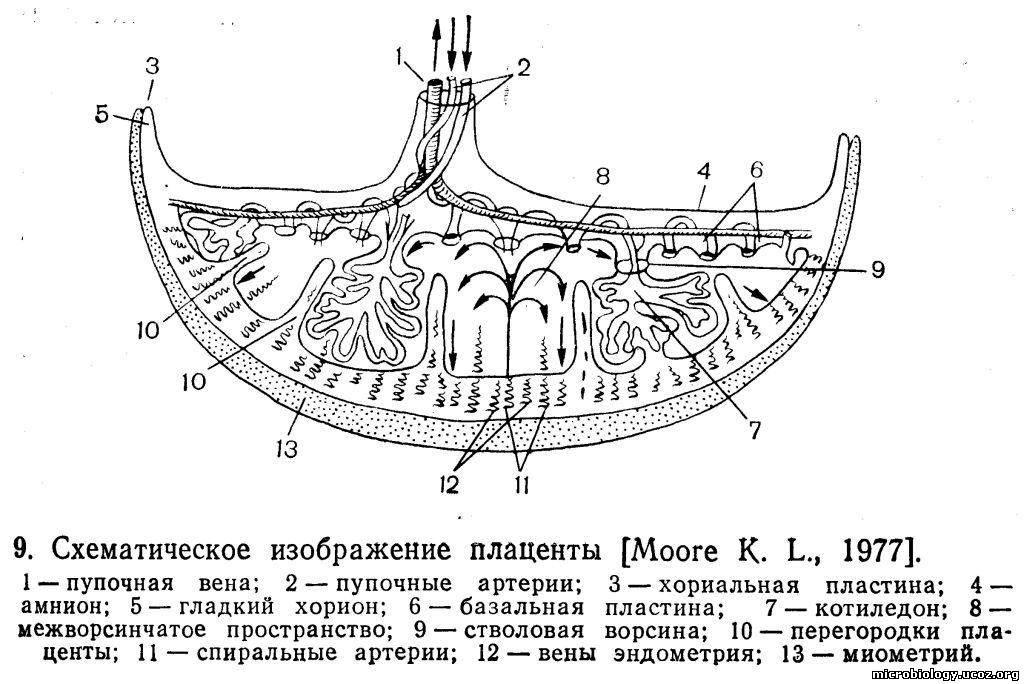
Миксобактерии
К этому порядку относят грамотрицательных бактерий, вегетативные клетки которых обладают большой гибкостью и способны к скользящему движению в слизи, продуцируемой ими в большом количестве. Клетки имеют палочковидную или веретенообразную форму с острыми концами. Большинство миксобактерии - аэробы, гетеротрофы.
Существенным отличием миксобактерии
от истинных бактерий является их
способность передвигаться за счет
активного сгибания тела (жгутики
отсутствуют). Такая особенность строения
(эластичная клеточная стенка) свойственна
простейшим животным организмам. Другой
особенностью миксобактерии является
образование плоских стелющихся по
поверхности слизистых колоний, образующих
выросты - псевдоплазмодий. Многие из
них способны к образованию плодовых
тел, напоминающих плодовые тела
простейших. Эти тела, имеющие причудливую
форму, но небольшие размеры (около 7 мм),
являются разновидностью колониальной
формы (рис. 113) и представляют собой
стадию образования цист.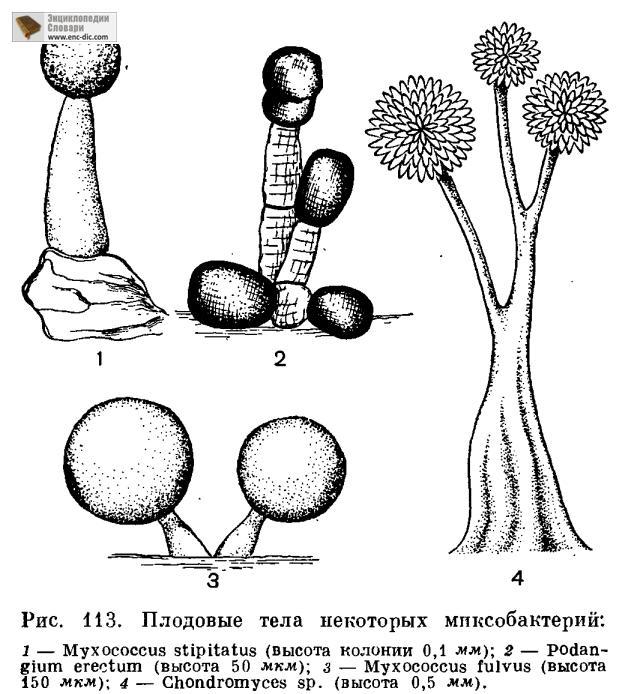
Систематическое положение миксобактерии
до сих пор вызывает споры. Одни специалисты
их считают родственными сине-зеленым
водорослям, другие - простейшим.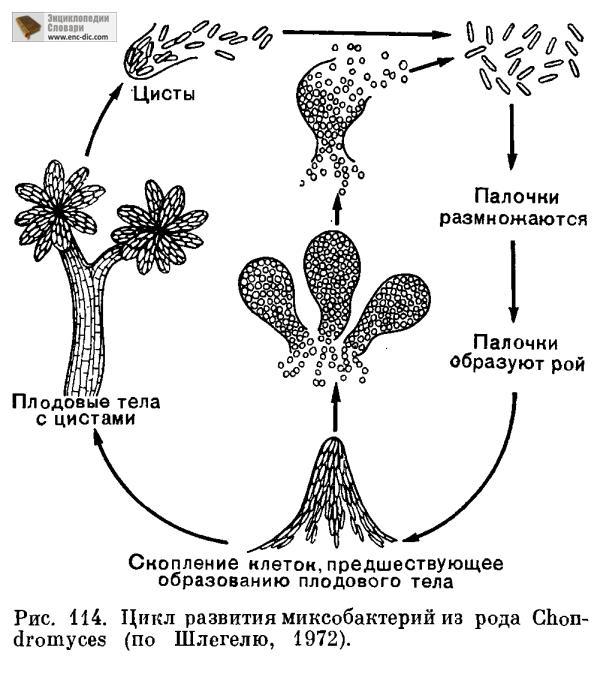
Как бы ни обстояло дело с таксономией, но та группа микроорганизмов со скользящим движением, которая образует плодовые тела, в цикле своего развития претерпевает смену форм, сходную со сменой форм у простейших: стадии роения и образования цист. На стадии псевдоплазмодия миксобактерии размножаются путем поперечного деления, выделяя обильную слизь, в которой живут и перемещаются. Эта стадия продолжается 1-7 дней, после чего палочки начинают образовывать скопления, растущие над поверхностью субстрата. У некоторых видов палочки передвигаются в верхней части этих сложных структур. Там палочки укорачиваются, переходя в состояние покоящихся клеток. У миксококков они, например, становятся овальными, окрашиваются в темный цвет, приобретают толстые оболочки и становятся похожими на хламидоспоры (микроцисты) (рис. 114). При увлажнении покоящиеся вегетативные клетки прорастают и снова переходят к стадии псевдоплазмодия.
Истинные миксобактерии, образующие плодовые тела, представлены в природе следующими родами: миксококкус (Myxococcus), сорангиум (Sorangium), хондромицес (Chondromyces), хондрококкус (Chondrococcus), архангиум (Archangium), полиангиум (Polyangium), мелитангиум (Melittangium), подангиум (Podangium), стелангиум (Stelangium), синангиум (Synangium), ангиококкус (Angiococcus) (рис. 113).
По способности к скользящему движению, форме клеток с миксобактериями сходны два рода целлюлозоразлагающих бактерий - цитофага (Cytophaga) и спороцитофага (Sporocytophaga). Эти бактерии не образуют плодовых тел. Веретенообразные вегетативные клетки Sporocytophaga могут переходить в кокковидные микроцисты.
По ряду признаков подобны миксобактериям флексибактерии - обитатели пресной и морской воды. Клетки представителей рода Flexibacter имеют вид длинных гибких нитей толщиной менее 1 мкм, способных к скользящему движению.
Миксобактерий в природе играют очень важную роль. Многие представители этой группы активно разрушают мертвые растительные остатки (целлюлозу) и соединения, содержащиеся в наружных покровах насекомых и ракообразных (хитин), превращая их в вещества, пригодные для питания растений.
Многие плодоносящие бактерии способны к внеклеточному паразитизму. Не проникая внутрь клеток жертв других бактерий и некоторых высших грибов, они вызывают их разрушение (лизис), используя освобождающиеся при этом вещества как пищу. Известны также случаи паразитизма флексибактерии на водорослях.
|
Спирохеты
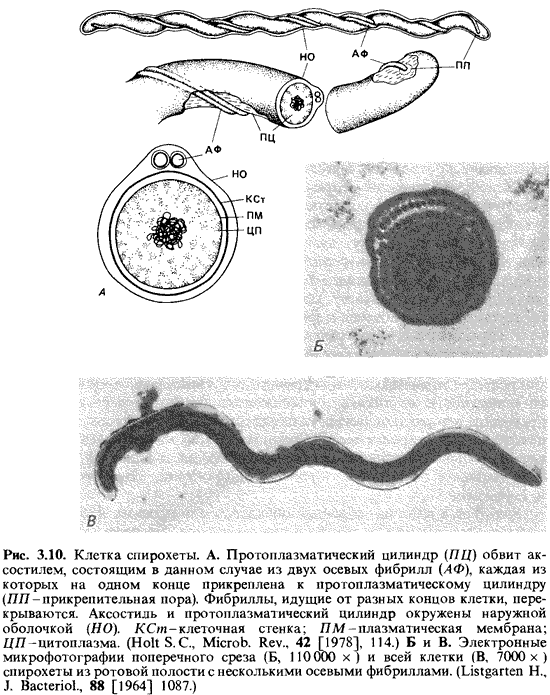
Спирохеты - группа одноклеточных хемогетеротрофных бактерий с очень характерной морфологией. Строением своих клеток и способом передвижения они отличаются от всех других бактерий. Клетка спиралевидная, как у спириллы, но не ригидная, а чрезвычайно гибкая. В сравнении с длиной (5-500 мкм) толщина ее необычно мала (0,1-0,6 мкм). Поэтому спирохеты проходят через мелкопористые фильтры (с отверстиями 0,2-0,45 мкм), задерживающие большинство бактерий, и путем фильтрования можно получать их накопительные культуры. Из-за малой толщины клетки спирохет трудно увидеть в светлом поле, и их обычно наблюдают в условиях фазового контраста или в темном поле. Строение клетки. В клетках спирохет можно различать три главных компонента: протоплазматический цилиндр, осевые фибриллы и наружную оболочку (рис. 3.10). Спирально извитой протоплазматический цилиндр снаружи окружен комплексом из плазматической мембраны и клеточной стенки. Вокруг цилиндра обвиваются нити, называемые по отдельности осевыми фибриллами, а в совокупности-аксостилем. Каждая фибрилла одним концом прикреплена вблизи конца клетки, а другой ее конец свободен. Число фибрилл варьирует от вида к виду: у Treponema pallidum и Leptospira имеется чаще всего 4 фибриллы, уBorrelia до 18, а у Cristispira более сотни. На обоих концах клетки прикреплено примерно одинаковое число фибрилл; в середине или по всей длине клетки фибриллы перекрывают друг друга. Наружная мембрана охватывает протоплазматический цилиндр и фибриллы. Способ
передвижения. Хотя
спирохеты не обладают жгутиками, они
способны плавать, не соприкасаясь с
твердыми поверхностями и не скользя
по ним. Вероятнее всего, в движении
участвуют фибриллы. Фибриллы сходны
с жгутиками по своему составу (состоят
из белка флагеллина), способу прикрепления
к телу клетки и спиральному расположению
рядов субъединиц. По-видимому, фибриллы,
вращаясь или же сокращаясь, обусловливают
характерное для спирохет движение:
эти бактерии могут изгибаться и
передвигаться змееобразно или толчками. Распространение, места обитания и важнейшие представители. Свободноживущие спирохеты могут быть обнаружены во многих водоемах (в лужах, прудах и в море). Другие относятся к нормальной автохтонной микрофлоре животных. Они встречаются в кишечнике млекопитающих, на поверхности жгутиковых, в кишечнике термитов, переваривающих древесину, и тараканов, в кристаллическом стебельке моллюсков, в рубце жвачных и в других местообитаниях. Лишь немногие из них патогенны (вызывают сифилис, возвратный тиф, лептоспирозы). Различают пять родов: Spirochaeta, Cristispira, Treponema, Borrelia и Leptospira. Spirochaeta plicatilis- обычный обитатель ила пресных и соленых водоемов. Она никогда не растет в очистных илах, но неизменно присутствует в мезосапробных зонах водоемов, в деревенских прудах и лужицах, а также в иле с большим количеством пурпурных бактерий. S. plicatilisобращает на себя внимание своим характерным безостановочным движением. В лаборатории ее удается культивировать лишь в течение короткого времени. Spirochaeta zuelzerae встречается в гниющих илах водоемов. Она строго анаэробна, может расти на питательной среде с дрожжевым экстрактом и глюкозой, способна использовать многие сахара и крахмал; лучше всего она растет при 37-40°С. Как по внешнему виду, так и по антигенной структуре эта бактерия сходна с Treponema pallidum и дает положительную реакцию связывания комплемента с сифилитической сывороткой. S. zuelzerae, так же как и спирохеты из рубца жвачных, сбраживает глюкозу до лактата, ацетата, сукци-ната, С02 и Н2, т.е. сходна по типу брожения сEscherichia coli. Cristispira живет в кристаллическом стебельке и желудочно-кишечном тракте пресноводных и морских двустворчатых моллюсков {Anodonta, Pecten, Venus и др.). Витки спирали более крутые, чем у Spirochaeta plicatilis; толщина тела клетки составляет 0,5-3 мкм; вокруг клетки - нежная оборка, винтообразно обвивающая цилиндр. Эта оборка (crista) по своему расположению соответствует аксиальной нити. Она состоит более чем из 100 отдельных фибрилл. К Treponemaceae относятся более мелкие формы спирохет, в том числе Treponema pallidum(возбудитель сифилиса) и Т. pertenue (возбудитель тропической болезни фрамбезии). Некоторые трепонемы-относительно безобидные паразиты ротовой полости; Т. macrodentumлегко можно выделить из слюны и налета на зубах. Т. pallidum еще никогда не удавалось надежно выращивать на искусственных средах. К роду Borrelia относят анаэробных спирохет, легко окрашиваемых анилиновыми красителями. Это паразиты различных членистоногих, а также возбудители болезней человека и других позвоночных («спирохеты крови»). В. recurrentis - возбудитель возвратного тифа. На В. anserinaЭрлих и Гата испытывали мышьяковистые препараты против сифилиса, пока не достигли успеха, найдя весьма действенное химиоте-рапевтическое средство - сальварсан. К роду Leptospira относятся самые мелкие аэробные спирохеты; толщина их 0,1-0,25 мкм, а длина 4-8 мкм. Они отличаются крючкообразно загнутыми концами клеток. L. biflexa- аэробная бактерия, обитает в пресной воде (прудах, лужах; ее можно выделить из водопроводной воды), растет на обычных питательных средах. Из патогенных лептос-пир были исследованы Leptospiraicterohaemorrhagiae (возбудитель «болезни Вейля»), L. ротопа (возбудитель болезни свинопасов) и L. canicola (возбудитель инфекционной желтухи). Эти лептоспиры попадают в организм с водой или пищей, проникают в кровь, почки и печень и нарушают функцию этих органов, что приводит к кровоизлияниям и желтухе. |
Микоплазмы
Микоплазмы - мельчайшие частицы. Они являются самыми мелкими самореплицирующими прокариотами. Морфология и размеры микоплазм варьируют в зависимости от возраста культуры, условий и сред культивирования. Микоплазмы полиморфны. Клетки микоплазм ограничены лишь трехслойной плазматической мембраной (внутриплазматические мембраны у микоплазм не обнаружены). В 1935 году из бактерий Streptobacillus moniliformis были изолированы фильтрующиеся формы, не имеющие клеточной стенки, которые удивительно напоминали микоплазмы. В последнее время их назвали L-формами бактерий. Состояние L-формы обусловлено влиянием неблагоприятных факторов внешней среды (например, применение антибиотиков, действующих на клеточную стенку). При их отсутствии L-форма обратима. У микоплазм, в отличие от других бактерий, состояние L-формы, т.е. отсутствие клеточной стенки, является их обычным состоянием [15,20].
Отсутствие клеточной стенки у микоплазм определяет их пластичность, которая позволяет этим микроорганизмам проникать через поры фильтров диаметром 0,22 - 0,45мкм. Из-за фильтруемости микоплазмы долгое время путали с вирусами. Сферическая форма клеток характерна для большинства видов микоплазм. При этом клетки одной и той же микоплазмы могут быть сферической (или несколько вытянутой) формы 0,3 - 0,8 мкм в диаметре, но могут образовывать длинные (до 100 мкм), иногда ветвящиеся тяжи, которые, проходя фазу коккоидных структур, распадаются на ряд сферических клеток, что изображено на рисунке 2. Коккоидные структуры иногда образуют кольцо.
Микоплазмы не образуют так называемых покоящихся форм или спор. Подобно другим неспорообразующим бактериям микоплазмы в неблагоприятных условиях переходят в некультивируемое состояние, а также образуют "минимальные тела", не способные к размножению, так как они, вероятно, не содержат ДНК [23].
Рисунок 2 - Морфология M. hominis а) - морфологические превращения при оптимальных условиях культивирования in vitro б) - морфологические формы при неоптимальных условиях, на стационарной фазе роста культуры
Некоторые виды микоплазм обладают скользящей подвижностью. Клетки таких микоплазм имеют особые структуры и цитоскелетоподобные образования. Так, клетки M. gallisepticum имеют грушевидную форму, M. pneumoniae также грушевидны, но более вытянуты, а M. mycoides чаще имеют форму тяжей.
Клетки большинства бактерий могут быть покрыты оболочкой - полимерной субстанцией, которая имеет множество свойств и функций. Эта оболочка, или капсула, отлична от двухслойной мембраны и располагается над ней. У бактерий термин "капсула" используется для определения высокомолекулярных полимеров, которые "прикрепляются" к поверхности бактерий. Хотя пептидогликановая клеточная стенка, которая характерна для большинства прокариот, и отсутствует у представителей класса Mollicutes, однако для некоторых видов микоплазм описаны капсулы или капсулоподобные структуры. Ими обладают Mycoplasma mycoides, M. gallisepticum, M. hyopneumoniae, M. meleagridis, M. dispar, M. pneumoniae, M. pulmonis, M. synoviae, M. hominis. Среди уреаплазм капсулы способны образовывать лишь некоторые штаммы Ureaplasma urealitycum.
Вирусы
Вирусы (лат. virus — яд животного происхождения) — мельчайшие организмы, которые по своей морфологии, биологическим свойствам, химическому составу и способу размножения отличаются от других микроорганизмов. Вирусы являются строгими (облигатными) внутриклеточными паразитами и не размножаются вне живых клеток. Размеры вирусов очень малы: от 25 до 250 нм, их определяют размерами пор фарфоровых фильтров, через которые они проходят при фильтрации. Существуют мелкие вирусы: полиомиелита, ящура, желтой лихорадки, размер которых до 25 нм, и более крупные: вирусы оспы и осповакцины, размер которых приближается к 250 нм. Самые крупные мантийные вирусы — орнитоза, трахомы и венерической лимфогранулемы — имеют размер до 300—350 нм. Однако их химический состав сложнее, чем у остальных вирусов. Поэтому мантийные вирусы выделены в особую группу микроорганизмов «хламидозоа», или хламидии (по классификации Берджи группа 18). Изучение формы вирусов и их строения возможно только в электронном микроскопе при увеличении в 50 000—300 000 раз. Крупные вирусы размером более 150 нм можно увидеть в обычном световом микроскопе при специальных методах окраски и увеличении в 900— 1000 раз. Форма вирусов может быть различной: шаровидной, овоидной, палочковидной, нитевидной, многогранной и булавовидной (рис. 14).
Зрелые частицы вируса называют вирионами. Вирион состоит из нуклеиновой кислоты, заключенной в белковую оболочку — капсид. Тип и свойства нуклеиновой кислоты имеют важное значение в классификации вирусов. Характерными признаками вирусов является содержание в вирионе только одной из нуклеиновых кислот: либо ДНК, либо РНК- Все остальные живые организмы содержат одновременно и ДНК, и РНК. В зависимости от типа нуклеиновой кислоты вирусы можно разделить на две большие группы: ДНК-содержащие и РНК-содержащие. Нуклеиновая кислота вирусов может состоять из одной нити (однонитчатая) или двух нитей (двунитчатая). Почти все РНК-содержащие вирусы имеют в своем геноме однонитчатую РНК. ДНК-содержащие вирусы чаще имеют двунитчатую ДНК и редко — однонитчатую.
У многих вирусов нуклеиновая кислота и белок (нуклеопротеид) находятся внутри вириона (сердцевина — core), а у некоторых вирусов нуклеиновая кислота непосредственно заключена в капсид (нуклеокапсид). Капсид состоит из повторяющихся белковых субъединиц, которые образованы одной или несколькими белковыми молекулами. Группы из нескольких белковых молекул можно видеть на электронных микрофотографиях. Такие структурные единицы, образующие часть капсида, называют капсомерами. Способ укладки капсомеров (тип симметрии) и количество их в капсиде неодинаковы у разных вирусов. Капсомеры могут быть уложены в капсидах в виде многогранника с равными симметричными гранями (кубический тип симметрии) или по спирали (спиральный тип симметрии). Спиральный тип симметрии имеют вирусы гриппа, а кубический — адено-, герпес- и энтеровирусы. Вирусы с кубическим типом симметрии называют также изометрическими. У вирусов группы оспы и бактериофагов сложный тип симметрии: например, головка бактериофага кубического типа, а отросток- хвост— спирального (рис. 15).
Многие вирусы животных и
человека обладают внешней оболочкой
(пеплос), окружающей их капсид. В состав
этих оболочек входят липиды или
липопротеиды. Наличие внешних оболочек
характерно для вирусов, созревание
которых происходит на внутренней
поверхности цитоплазматической мембраны
клетки хозяина. Проходя через поверхность
пораженной клетки, вирусный капсид как
бы обволакивается этой мембраной,
формируя один или несколько слоев своей
внешней оболочки. Внешнюю оболочку
имеют миксо-, герпес- и рабдовирусы
(вирус бешенства), а также тогавирусы
(вирусы энцефалитов). Внешние оболочки
этих вирусов, содержащие фосфолипиды,
разрушаются эфиром, что служит
отличительным признаком от вирусов,
которые имеют голый капсид. Вирусы
группы оспы также имеют внешние оболочки,
но формирование их происходит внутри
пораженной клетки и к эфиру они
нечувствительны. У некоторых вирусов
(миксо- и тогавирусы) из внешнего липидаого
слоя выступают наружу вирусспецифические
гликопротеиды. Эти выступающие капсомеры
называют шипами (пепломеры), хотя концы
их не острые, а тупые.
Мелкие вирусы,
имеющие форму палочек или шариков, могут
образовывать упорядоченные структуры
в виде кристаллов. Кристаллы состоят
из сотен миллиардов вирусных частиц,
тесно прижатых друг к другу.
Вирусы
оспы, бешенства, хламидии трахомы и
некоторые другие, поражая различные
клетки организма, образуют в них
внутриклеточные включения. Это крупные
и плотные гранулы, которые при определенной
окраске могут быть обнаружены в световом
микроскопе. Внутриклеточные включения
располагаются в ядре клетки или ее
цитоплазме. Происхождение их пока
неясно.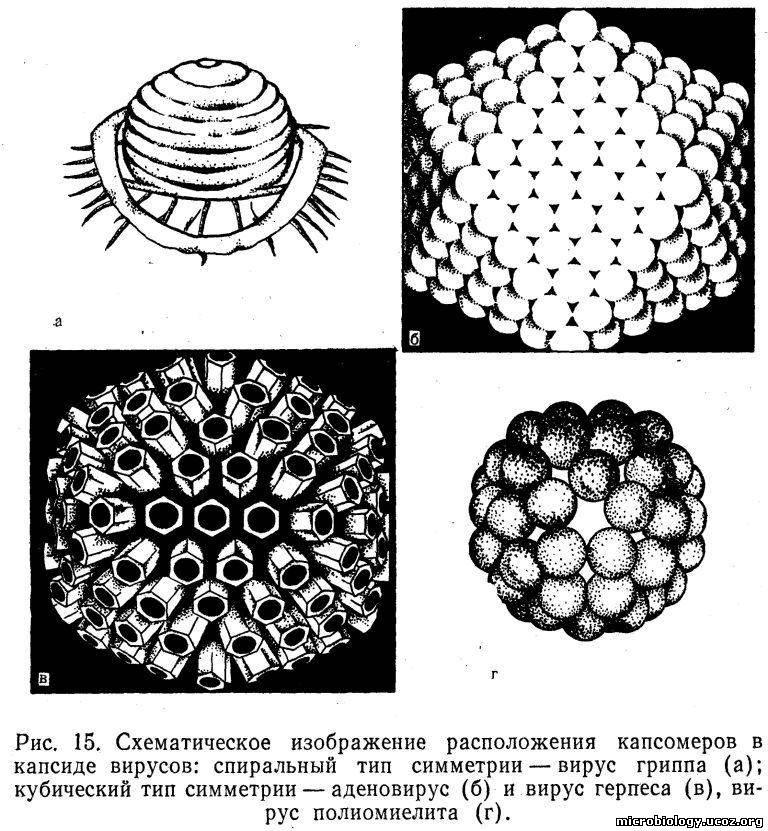
В одних случаях они являются внутриклеточными колониями (скоплениями) элементарных частиц вируса (вирионов), в других — продуктами реакции клетки хозяина на внедрившийся вирус. Обнаружение внутриклеточных включений при некоторых инфекциях служит диагностическим признаком заболевания: например, тельца Негри при бешенстве, тельца Гварниери при оспе.
Используемая литература
1. http://www.krugosvet.ru/enc/nauka_i_tehnika/biologiya/BAKTERII.html?page=0,2
2. http://floralesa.ru/nizshie_rasteniya/klass_aktinomicety_ili_luchistye_griby__actinomycetes
3. http://microbiology.ucoz.org/index/virusy/0-15
4. http://mir-prekrasen.net/referat/3653-miksobakterii.html
5. http://enc-dic.com/enc_biology/Porjadok-miksobakterii-myxobacteriales-1009/