

48. Генотип, геном, фенотип. Взаимодействие аллелей в детерминации признаков: доминирование, промежуточное проявление, рецессивность, кодоминирование.
Геноти́п — совокупность генов данного организма. генотип определяет норму реакции организма на все возможные условия среды. Взаимодействие генотипа с комплексом факторов внутренней и внешней среды организма обусловливает фенотипическое проявление признаков. Примером влияния среды на фенотипическое проявление генотипа может служить окраска меха у кроликов так называемой гималайской линии: при одном и том же генотипе эти кролики при выращивании на холоде имеют чёрный мех, при умеренной температуре — «гималайскую» окраску (белая с чёрными мордой, ушами, лапами и хвостом), при повышенной температуре — белый мех. Фенотип — совокупность внешних и внутренних признаков организма, приобретённых в результате онтогенеза (индивидуального развития).людается корреляции между уровнем эволюционной сложности биологического вида и размером его генома.Большинство генов проявляются в фенотипе организма, но фенотип и генотип различны по следующим показателям:1. По источнику информации (генотип определяется при изучении ДНК особи, фенотип регистрируется при наблюдении внешнего вида организма).2. Генотип не всегда соответствует одному и тому же фенотипу. Некоторые гены проявляются в фенотипе только в определенных условиях. С другой стороны, некоторые фенотипы, например, окраска шерсти животных, являются результатом взаимодействия нескольких генов .Примером различия генотипа и фенотипа служит наследование гемофилии. Иногда в семье, в которой оба родителя здоровы, рождается больной ребенок. То есть хотя болезнь не проявилась в фенотипе родителей, в их генотипе присутствовал один нормальный аллель и один мутированный аллель гена, то есть они являлись носителями заболевания. В данном случае фенотип здоровых людей и носителей заболевания одинаков.Гено́м —совокупность всех групп сцепленных хромосомных и внехромосомных ДНК в клетке организма данного вида, генетическая характеристика вида. Homo sapiens включает: 24 Днк хромосом( 22 аутосомы, Х и У хромосомы).Геномы про- и эукариот:1)Хромосомная ДНК: эукариоты- в нуклеотиде, прокариоы- в ядре ; 2)внехромосомная ДНК: прокариоты-плазмиды, эукариоты- ДНК митохондрий и хролопластов 5%.«Геном- гаплоидное число хромосом, специфичное для организмов каждого вида»( Винклер.Г 1920).Следовательно, под геномом организма понимают суммарную ДНК гаплоидного набора хромосом и каждого из внехромосомных генетических элементов, содержащуюся в отдельной клетке зародышевой линии многоклеточного организма. В определении генома отдельного биологического вида необходимо учитывать, во-первых, генетические различия, связанные с полом организма, поскольку мужские и женские половые хромосомы различаются. Во-вторых, из-за громадного числа аллельных вариантов генов и сопутствующих последовательностей, которые присутствуют в генофонде больших популяций, можно говорить лишь о некоем усреднённом геноме, который сам по себе может обладать существенными отличиями от геномов отдельных особей. Размеры геномов организмов разных видов значительно отличаются друг от друга, и при этом часто не наб..Кодоминирование и неполное доминирование:Некоторые противоположные признаки находятся не в отношении полного доминирования (когда один всегда подавляет другой у гетерозиготных особей), а в отношении неполного доминирования. Например, при скрещивании чистых линий львиного зева с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. При неполном доминировании гетерозиготы имеют признаки, промежуточные между признаками рецессивной и доминантной гомозигот.При кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки проявляются одновременно (смешанно). Типичный пример кодоминирования — наследование групп крови системы АВО у человека. Всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвертая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В).Явления кодоминирования и неполного доминирования признаков слегка видоизменяет первый закон Менделя: «Гибриды первого поколения от скрещивания чистых линий особей с противоположными признаками всегда одинаковы по этому признаку: проявляют доминирующий признак, если признаки находятся в отношении доминирования, или смешанный (промежуточный) признак, если они находятся в отношении кодоминирования (неполного доминирования)».Для своих первых опытов Мендель выбирал растения, чётко различающиеся по какой-либо паре признаков, например, по расположению цветов («пазушные» или «верхушечные»). Выращивая растения каждого типа на протяжении нескольких поколений, Мендель убедился в их пригодности для проведения эксперимента. Мендель проводил скрещивание – опылял растения одного типа пыльцой растений другого типа. Ряд предосторожностей (например, удаление тычинок у цветков, которые впоследствии опылялись, и надевание колпачков на цветы, чтобы избежать дополнительного опыления со стороны других растений) позволили получить достоверные результаты. Во всех случаях из семян, собранных с этих гибридов, вырастали растения с пазушными цветками. Признак «пазушные цветки», наблюдаемый у гибридов первого поколения, был назван доминантным, признак «верхушечные цветки» – рецессивным.Далее растениям первого гибридного поколения была предоставлена возможность самоопылиться. Во втором гибридном поколении у части растений образовались пазушные цветки, а у другой части – верхушечные. Мендель предположил, что признак «верхушечные цветки» присутствовал и в первом поколении, но в скрытом виде. Во всех подобных опытах, проведённых с какой-либо парой признаков, примерно три четверти гибридов второго поколения обладали признаком, проявлявшимся и в первом поколении гибридов (его назвали доминантным), а четверть потомства второго поколения обладала признаком, не проявившимся у гибридов первого поколения (рецессивным). Важно, что чем больше опытов было поставлено, тем ближе был полученный результат к отношению 3 : 1.
49. Закономерности наследования при моногибридном скрещивании.Моногибридное скрещивание — скрещивание форм, отличающихся друг от друга по одной паре альтернативных признаков. При этом скрещиваемые предки являются гетерозиготными по положению хромосомы в аллели. Например, при скрещивании видов с признаками K=коричневый и к=красный, результат будет КК=коричневый; Кк=коричневый, а не красный; кк=красный. Здесь К — доминантный признак, а к — рецессивный.Моногибридное наследование — наследование единственной особенности. Различные формы признаков обычно управляются различными аллелями того же самого гена. Например, при моногибридном скрещивании между двумя чистыми линиями растений, (гомозиготных по соответствующим им характерным признакам) одного с жёлтыми семенами (доминантный признак), а другого с зелёными семенами (рецессивный признак), можно ожидать, что первое поколение будет только с жёлтыми семенами, потому что аллель жёлтых семян является доминирующим над зелёными. При моногибридном скрещивании сравнивают только один характерный признак.Условия выполнения закона расщепления при моногибридном скрещивании.Расщепление 3 : 1 по фенотипу и 1 : 2 : 1 по генотипу выполняется приближенно и лишь при следующих условиях:Изучается большое число скрещиваний (большое число потомков). Гаметы, содержащие аллели А и а, образуются в равном числе (обладают равной жизнеспособностью).Нет избирательного оплодотворения: гаметы, содержащие любой аллель, сливаются друг с другом с равной вероятностью.Зиготы (зародыши) с разными генотипами одинаково жизнеспособны.
50. Независимое комбинирование неаллельных генов и его цитологические основы.Наследование признаков, сцепленных с поломВ том случае, когда гены, контролирующие формирование того или иного признака, локализованы в аутосомах, наследование осуществляется независимо от того, кто из родителей (мать или отец) является носителем изучаемого признака. Если же гены находятся в половых хромосомах, характер наследования признаков резко изменяется. Например, у дрозофилы гены, локализованные в X-хромосоме, как правило, не имеют аллелей в У-хромосоме. По этой причине рецессивные гены в X-хромосоме гетерогаметного пола практически всегда проявляются, будучи в единственном числе.Признаки, гены которых локализованы в половых хромосомах, называются признаками, сцепленными с полом. Явление наследования, сцепленного с полом, было открыто Т. Морганом у дрозофилы.Х- и У-хромосомы у человека имеют гомологичный (псевдоаутосомный) участок, где локализованы гены, наследование которых не отличается от наследования аутосомных генов.Помимо гомологичных участков, X- и У-хромосомы имеют негомологичные участки. Негомологичный участок У-хромосомы, кроме генов, определяющих мужской пол, содержит гены перепонок между пальцами ног и волосатых ушей у человека. Патологические признаки, сцепленные с негомологичным участком У-хромосомы, передаются всем сыновьям, поскольку они получают от отца У-хромосому.Негомологичный участок X-хромосомы содержит в своем составе ряд важных для жизнедеятельности организмов генов. Поскольку у гетерогаметного пола (ХУ) X-хромосома представлена в единственном числе, то признаки, определяемые генами негомологичного участка X-хромосомы, будут проявляться даже в том случае, если они рецессивны. Такое состояние генов называется гемизиготным. Примером такого рода X-сцепленных рецессивных признаков у человека являются гемофилия, мышечная дистрофия Дюшена, атрофия зрительного нерва, дальтонизм (цветовая слепота) и др.Гемофилия — это наследственная болезнь, при которой кровь теряет способность свертываться. Ранение, даже царапина или ушиб, могут вызвать обильные наружные или внутренние кровотечения, которые нередко заканчиваются смертью. Это заболевание встречается, за редким исключением, только у мужчин. Было установлено, что обе наиболее распространенные формы гемофилии (гемофилия А и гемофилия В) обусловлена рецессивными генами, локализованными в X-хромосоме. Гетерозиготные по данным генам женщины (носительницы) обладают нормальной или несколько пониженной свертываемостью крови.Фенотипическое проявление гемофилии у девочек будет наблюдаться в том случае, если мать девочки является носительницей гена гемофилии, а отец — гемофиликом. Подобная закономерность наследования характерна и для других рецессивных, сцепленных с полом признаков.Сцепленное наследованиеНезависимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы до эры молекулярной биологии было изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. То, что число генов у высших организмов составляет несоклько тысяч, было ясно уже У. Сэттону в начале XX века. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе.Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием. Число групп сцепления соответствует гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. (У особей гетерогаметного пола, например, у самцов млекопитающих, групп сцепления на самом деле на одну больше, так как X- и У-хромосомы содержат разные гены и представляют собой две разные группы сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин — 24).Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и ab) в равных количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.Было установлено, однако, что кроме обычных (некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми комбинациями генов — Ab и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной возникновения таких гамет является обмен участками гомологичных хромосом, или кроссинговер.Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
51. Дигибридное и полигибридное скрещивания. Общая формула расщепления при независимом наследовании.Сущность дигибридного скрещивания. Организмы различаются по многим генам и, как следствие, по многим признакам. Чтобы одновременно проанализировать наследование нескольких признаков, необходимо изучить наследование каждой пары признаков в отдельности, не обращая внимания на другие пары, а затем сопоставить и объединить все наблюдения. Именно так и поступил Мендель. Скрещивание, при котором родительские формы отличаются по двум парам альтернативных признаков (по двум парам аллелей), называется дигибридным. Гибриды, гетерозиготные по двум генам, называют дигетерозиготными, а в случае отличия их по трем и многим генам —три- и полигетерозиготными соответственно. Результаты дигибридного и полигибридного скрещивания зависят от того, располагаются гены, определяющие рассмотренные признаки, в одной хромосоме или в разных. Независимое наследование (третий закон Менделя). Для дигибридного скрещивания Мендель использовал гомозиготные растения гороха, различающиеся одновременно по двум парам признаков. Одно из скрещиваемых растений имело желтые гладкие семена, другое — зеленые морщинистые (рис. 3.3).
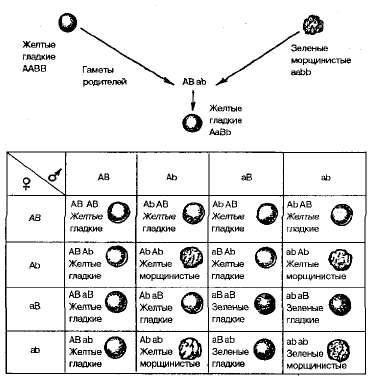
Рис 3.3. Дигибридное скрещивание растений гороха, различающихся по форме и окраске семян. Все гибриды первого поколения этого скрещивания имели желтые гладкие семена. Следовательно, доминирующими оказались желтая окраска семян над зеленой и гладкая форма над морщинистой. Обозначим аллели желтой окраски А, зеленой — а, гладкой формы— В, морщинистой— b. Гены, определяющие развитие разных пар признаков, называются неаллельпыми и обозначаются разными буквами латинского алфавита. Родительские растения в этом случае имеют генотипы АА ВВ и aabb, а генотип гибридов F1 —АаВb ,т. е. является дигетерозиготным. Во втором поколении после самоопыления гибридов F1 в соответствии с законом расщепления вновь появились морщинистые и зеленые семена. При этом наблюдались следующие сочетания признаков: 315 желтых гладких, 101 желтое морщинистое, 108 зеленых гладких и 32 зеленых морщинистых семян. Это соотношение очень близко к соотношению 9:3:3:1. Чтобы выяснить, как ведет себя каждая пара аллелей в потомстве дигетерозиготы, целесообразно провести раздельный учет каждой пары признаков — по форме и окраске семян. Из 556 семян Менделем получено 423 гладких и 133 морщинистых, а также 416 желтых и 140 зеленых. Таким образом, и в этом случае соотношение доминантных и рецессивных форм по каждой паре признаков свидетельствует о моногибридном расщеплении по фенотипу 3:1. Отсюда следует, что дигибридное расщепление представляет собой два независимо идущих моногибридных расщепления, которые как бы накладываются друг на друга. Проведенные наблюдения свидетельствуют о том, что отдельные пары признаков ведут себя в наследовании независимо. В этом сущность третьего закона Менделя — закона независимого наследования признаков, или независимого комбинирования генов. Он формулируется так: каждая пара аллельных генов (и альтернативных признаков, контролируемых ими) наследуется независимо друг от друга. Закон независимого комбинирования генов составляет основу комбинативной изменчивости (см. § 3.4), наблюдаемой при скрещивании у всех живых организмов. Отметим также, что в отличие от первого закона Менделя, который справедлив всегда, второй закон действителен только для генов, локализованных в разных парах гомологичных хромосом. Это обусловлено тем, что негомологичные хромосомы комбинируются в клетке независимо друг от друга, что было доказано не только при изучении ха- рактера наследования признаков, но и прямым цитологическим методом. Поведение хромосом при дигибридном скрещивании показано на рис. 3.4. Рассуждая аналогично, можно представить расщепление при три- и полигибридном скрещивании, т.е. когда родители различаются по аллелям трех и более генов, а в F1 образуются три- и полигетерозиготы. Соотношение генотипических и фенотипических классов в F2 три- и полигибридных скрещиваний, а также число типов гамет (и число фенотипов) у гибридов F1 определяются простыми формулами: при моногибридном скрещивании число типов гамет равно 2, при дигибридном 4(22), а при полигибридном – 2n; число генотипов равно соответственно 3, 9(32) и 3n.Полигибридное скрещивание – это скрещивание организмов, анализируемых по трем и более парам альтернативных признаков. Механизм наследования двух, трех и многих пар признаков, определяемых генами, лежащими в разных негомологичных хромосомах, в принципе не отличается от механизма наследования одной пары признаков. В основе этих скрещиваний лежит одна и та же закономерность.
52. Принцип анализирующего скрещивания и его использование в генетическом анализе. Анализирующее скрещивание в случае независимого и сцепленного наследования.Анализирующее скрещивание — скрещивание гибридной особи с особью, гомозиготной по рецессивным аллелям, то есть "анализатором". Смысл анализирующего скрещивания заключается в том, что потомки от анализирующего скрещивания обязательно несут один рецессивный аллель от "анализатора", на фоне которого должны проявиться аллели, полученные от анализируемого организма. Для анализирующего скрещивания (исключая случаи взаимодействия генов) характерно совпадение расщепления по фенотипу с расщеплением по генотипу среди потомков. Таким образом, анализирующее скрещивание позволяет определить генотип и соотношение гамет разного типа, образуемых анализируемой особью.При скрещивании с организмом, рецессивным по обоим генам aabb, получается расщепление 1:1
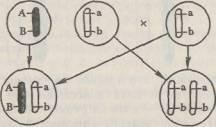
вместо ожидаемого при дигибридном анализирующем скрещивании 1:1:1:1. Такое отклонение от независимого распределения означает, что гены, локализованные в одной хромосоме, наследуются совместно.Явление совместного наследования генов, локализованных в одной хромосоме, называется сцепленным наследованием, а локализация генов в одной хромосоме — сцеплением генов. Сцепленное наследование генов, локализованных в одной хромосоме, установил Морган.
53. Условия менделирования признаков. Менделирующие признаки у человека – примерыМенделирующими признаками называются те, наследование которых про исходит по закономерностям, установленным Г. Менделем. Менделирующие признаки определяются одним геном моногенно (от греч.monos-один) то есть когда проявление признака определяется взаимодействием аллельных генов, один из которых доминирует (подавляет) другой. Менделевские законы справедливы для аутосомных генов с полной пенетрантностью (от лат.penetrans-проникающий, достигающий) и постоянной экспрессивностью (степенью выраженности признака). Если гены локализованы в половых хромосомах (за исключением гомологичного участка в Х- и У-хромосомах), или в одной хромосоме сцеплено, или в ДНК органоидов, то результаты скрещивания не будут следовать законам Менделя. Общие законы наследственности одинаковы для всех эукариот. У человека также имеются менделирующие признаки, и для него характерны все типы их наследования: аутосомно-доминантный, аутосомно-рецессивный, сцепленный с половыми хромосомами (с гомологичным участком Х- и У-хромосом).У человека менделирующими признаками являются, напр., альбинизм (отсутствие пигментации, вызываемое рецессивным геном; встречается у всех человеческих рас с частотой 1 на 20— 30 тыс. новорожденных), цвет глаз, характер волос (курчавые или гладкие), групповые отличия по различным факторам в крови (см. Группы крови) и др. Законам Менделя подчиняются и гены, обусловливающие наследственные болезни человека.
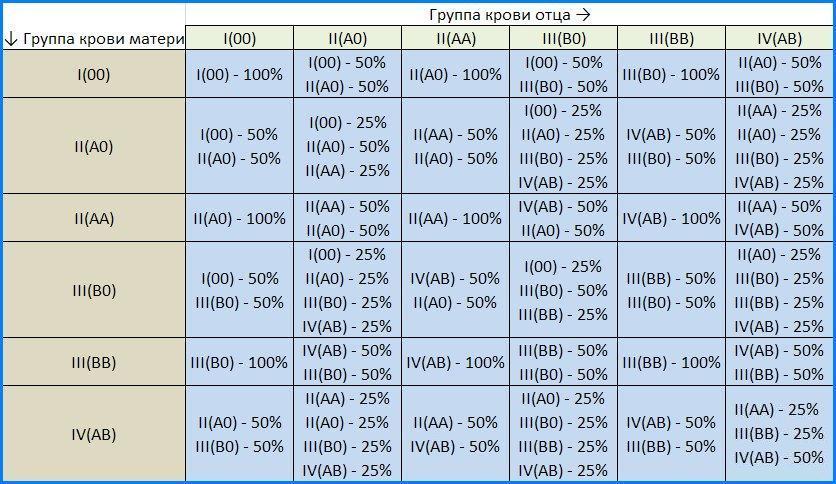
54. Множественные аллели. Наследование групп крови (АВО - системы) у человека.Множественный аллелизм — это существование в популяции более двух аллелей данного гена. В популяции оказываются не два аллельных гена, а несколько. Возникают в результате разных мутаций одного локуса. Гены множественных аллелей взаимодействуют между собой различным образом.В популяциях как гаплоидных, так и диплоидных организмов обычно существует множество аллелей для каждого гена. Это следует из сложной структуры гена — замена любого из нуклеотидов или иные мутации приводят к появлению новых аллелей. Видимо, лишь в очень редких случаях любая мутация столь сильно влияет на работу гена, а ген оказывается столь важным, что все его мутации приводят к гибели носителей. Так, для хорошо изученных у человека глобиновых генов известно несколько сотен аллелей, лишь около десятка из них приводит к серьезным патологиям.Гру́ппа кро́ви — описание индивидуальных антигенных характеристик эритроцитов, определяемое с помощью методов идентификации специфических групп углеводов и белков, включённых в мембраны эритроцитов животных.В наследовании групп крови есть несколько очевидных закономерностей:Если хоть у одного родителя группа крови I(0), в таком браке не может родиться ребёнок с IV(AB) группой крови, вне зависимости от группы второго родителя.1.Если у обоих родителей I группа крови, то у их детей может быть только I группа. 2.Если у обоих родителей II группа крови, то у их детей может быть только II или I группа. 3.Если у обоих родителей III группа крови, то у их детей может быть только III или I группа. 4.Если хоть у одного родителя группа крови IV(AB), в таком браке не может родиться ребёнок с I(0) группой крови, вне зависимости от группы второго родителя. 5.Наиболее непредсказуемо наследование ребенком группы крови при союзе родителей со II и III группами. Их дети могут иметь любую из четырёх групп крови.[30] Остальные варианты рассмотрены в таблице:
 55.
Полигенное наследование - примеры, в
том числе и у человека. это
Н. признака, контролируемого
совместно группой неаллельных
генов.
Широко распространенные болезни человека,
например, артериальная гипертензия,
ишемическая болезнь сердца, бронхиальная
астма, язвенная болезнь желудка,
наследуются полигенно. При этом тяжесть
заболевания зависит не только от
совокупного действия множества генов,
но и от провоцирующих средовых факторов.
При
скрещивании сорта пшеницы,
зерна которой имели темно-красную
окраску, с сортом, имеющим белые зерна,
гибриды первого поколения имели красную
окраску более светлых тонов. Во втором
поколении получилось такое соотношение
по фенотипу: на 63 окрашенных зерна с
различными оттенками красного цвета
приходилась 1 белое зерно (неокрашенное).
Эти результаты были объяснены Нильсоном-Эле
следующим образом. Темно-красная окраска
зерен пшеницы обусловлена действием
трех пар доминантных генов, а белая -
трех пар рецессивных, при этом по мере
увеличения числа доминантных генов
окраска становится более интенсивной.
Обозначим доминантные аллели трех
генов, локализованных в разных хромосомах,
прописными буквами А1 А2 А3 а рецессивные
- строчными а1 а1 а3, тогда генотипы
исходных форм будут: А1А1 А2А2 А3А3 x а1я1
а2а2 a33a.Окраска
зерен у гибридов первого
поколения A1a1 A2a2 A3a3 при наличии трех
доминантных аллелей будет промежуточной
светло-красной. При скрещивании гибридов
первого поколения A1a1 A2a2 A3a3 x A1a1 A2a2 A3a3 у
каждого из гибридов образуется по 8
типов гамет, поэтому во втором поколении
ожидается расщепление в 64-х долях (8 х
8). Среди 63/64 растений с окрашенными
зернами интенсивность окраски усиливается
по мере увеличения числа доминантных
аллелей различных генов в генотипе.
Видимо, каждый доминантный ген способствует
увеличению количества синтезированного
пигмента, и в этом смысле такой признак
можно отнести к количественным
55.
Полигенное наследование - примеры, в
том числе и у человека. это
Н. признака, контролируемого
совместно группой неаллельных
генов.
Широко распространенные болезни человека,
например, артериальная гипертензия,
ишемическая болезнь сердца, бронхиальная
астма, язвенная болезнь желудка,
наследуются полигенно. При этом тяжесть
заболевания зависит не только от
совокупного действия множества генов,
но и от провоцирующих средовых факторов.
При
скрещивании сорта пшеницы,
зерна которой имели темно-красную
окраску, с сортом, имеющим белые зерна,
гибриды первого поколения имели красную
окраску более светлых тонов. Во втором
поколении получилось такое соотношение
по фенотипу: на 63 окрашенных зерна с
различными оттенками красного цвета
приходилась 1 белое зерно (неокрашенное).
Эти результаты были объяснены Нильсоном-Эле
следующим образом. Темно-красная окраска
зерен пшеницы обусловлена действием
трех пар доминантных генов, а белая -
трех пар рецессивных, при этом по мере
увеличения числа доминантных генов
окраска становится более интенсивной.
Обозначим доминантные аллели трех
генов, локализованных в разных хромосомах,
прописными буквами А1 А2 А3 а рецессивные
- строчными а1 а1 а3, тогда генотипы
исходных форм будут: А1А1 А2А2 А3А3 x а1я1
а2а2 a33a.Окраска
зерен у гибридов первого
поколения A1a1 A2a2 A3a3 при наличии трех
доминантных аллелей будет промежуточной
светло-красной. При скрещивании гибридов
первого поколения A1a1 A2a2 A3a3 x A1a1 A2a2 A3a3 у
каждого из гибридов образуется по 8
типов гамет, поэтому во втором поколении
ожидается расщепление в 64-х долях (8 х
8). Среди 63/64 растений с окрашенными
зернами интенсивность окраски усиливается
по мере увеличения числа доминантных
аллелей различных генов в генотипе.
Видимо, каждый доминантный ген способствует
увеличению количества синтезированного
пигмента, и в этом смысле такой признак
можно отнести к количественным
56. Взаимодействие неаллельных генов - комплементарность, эпистаз, плейотропия - примеры.Комплементарное действие генов наиболее четко проявляется, когда скрещиваются две белые формы некоторых животных (кур) или растений (душистого горошка, белого клевера, кукурузы), а в потомстве появляются окрашенные формы. При скрещивании двух рас душистого горошка с белыми цветками (Lathyrus odoratus) в FI формируются растения с пурпурной окраской. При самоопылении этих растений в р2 наблюдается отклонение от менделевского расщепления: 9/ie растений имеют цветки с пурпурной окраской, тогда как 7/ie- с белой. В этом случае сущность взаимодействия генов, так же как при образовании ореховидного гребня у кур, состоит в том, что окрашенные цветки появляются при взаимодействии двух неаллельных доминантных генов А и В.Генотипы растений с неокрашенными цветками могут быть AAbb или ааВВ; если такие растения скрещиваются, то генотип растений FI будет АаВЬ. Это значит, что все растения этого поколения будут иметь пурпурные цветки. У кукурузы установлено, что для возникновения окрашенных семян необходим антоциан, который образуется только в присутствии двух доминантных генов А и В.Эпистатическое действие генов. Эпистаз - подавление действия одного гена другим, не аллельным, геном. Ген-подавитель, или супрессор, действует на подавляемый гипостатический ген по принципу, близкому к доминантности - рецессивности. Разница тут состоит в том, что эпистатический и гипостатический гены не являются ал-лельными, т. е. занимают различные локусы в гомологичных или негомологичных хромосомах. Эпистаз широко распространен в природе, в некоторых случаях изучены биохимические механизмы эпистатических взаимодействий. Так, у домашних птиц, в частности у кур, имеется зпистатическая система из двух генов, влияющих на окраску оперения. Эпистатический ген сам по себе не влияет на окраску пера. В то же время ген С в доминантной форме определяет нормальную продукцию пигмента. Однако белые леггорны с генотипом ССП не имеют пигмента в результате действия эпистатического гена. Есть и другие белые породы кур, окраска которых определяется другим генотипом. Так, оперение белых виандотов обусловлено тем, что они гомозиготны по рецессивным генам ее и ii, тур.-и по гипостатическому, и по эпистатическому генам. Расщепление при эпистазе, которое является результатом дигибридного скрещивания, отличается и от классического менделевского, и от того, которое наблюдается при комп- лементарностиАнализ данных на решетке Пеннета показывает, что только у 3/i6 потомства будет присутствовать ген С в доминантной форме и одновременно эпистатический ген в рецессивной форме i. Соотношение белых и пестрых птиц составит 13:3. Этот тип взаимодействия неаллельных генов называют доминантным эпистазом. В отличие от него при рецессивном эпистазе рецессивная аллель одного гена, будучи в гомозиготном состоянии, подавляет проявление доминантной или рецессивной аллели другого гена, В этом случае вместо ожидаемого при дигибридном скрещивании расщепления 9:3:3:1 получается отношение 9:7.Полимерия. До сих пор рассматривалось действие генов, определяющих контрастирующие, альтернативные признаки. С такого рода генами имел дело Мендель, экспериментируя на горохе. Однако уже с первого десятилетия XX в. известны гены, одинаково влияющие на признаки. Их действие может быть тождественным, но чаще бывает аддитивным, т. е. суммирующимся. В первом случае для развития признака достаточно присутствия одной из аллелей у дигетерозиготы в доминантной форме, тогда как аддитивное действие выражается в степени проявления признаков в зависимости от числа доминантных аллелей. Так, при аддитивном действии фенотип будет более выражен при генотипе ААВВ, чем при АаВЬ.
57. Сцепление генов. Основные положения хромосомной теории наследственности. Полное и неполное сцепление, кроссинговер.Сцепленное наследование — феномен скоррелированного наследования определенных состояний генов, расположенных в одной хромосоме.Полной корреляция не бывает из-за мейотического кроссинговера, так как сцепленные гены могут разойтись по разным гаметам. Кроссинговер наблюдается в виде расцепления у потомства тех аллелей генов и, соответственно, состояний признаков, которые были сцеплены у родителей.Наблюдения, проведенные Томасом Морганом, показали, что вероятность кроссинговера между различными парами генов разная, и появилась идея создать генные карты на основании частот кроссинговера между разными генами. Первая генная карта была построена студентом Моргана, Альфредом Стёртевантом (англ.) в 1913 году на материале Drosophila melanogaster.Расстояние между генами, расположенными в одной хромосоме, определяется по проценту кроссинговера между ними и прямо пропорционально ему. За единицу расстояния принят 1% кроссинговера (1 морганида или 1 сантиморганида). Чем дальше гены находятся друг от друга в хромосоме, тем чаще между ними будет происходить кроссинговер. Максимальное расстояние между генами, расположенными в одной хромосоме, может быть равно 49 сантиморганидам.Основные положения хромосомной теории наследственностиАнализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален. Аллельные гены занимают одинаковые локусы в гомологичных хромосомах. Гены расположены в хромосоме в линейной последовательности. Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола). Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами). Каждый биологический вид характеризуется определенным набором хромосом — кариотипом. Кроссинго́вер (другое название в биологии перекрёст) — явление обмена участками гомологичных хромосом во время конъюгации при мейозе. Помимо мейотического описан также митотический кроссинговер.Во время пахитены (стадия толстых нитей), гомологичные хромосомы находятся в состоянии конъюгации длительный период: у дрозофилы - четверо суток, у человека больше двух недель. Все это время отдельные участки хромосом находятся в очень тесном соприкосновении. Если в таком участке произойдет разрыв цепочек ДНК одновременно в двух хроматидах, принадлежащих разным гомологам, то при восстановлении разрыва может получиться так, что ДНК одного гомолога окажется соединенной с ДНК другой, гомологичной хромосомы. Этот процесс носит -название кроссинговера (англ. crossing-over - перекрест).Кроссинговер - взаимный обмен гомологичными участками хромосом между гомологичными (парными) хромосомами исходных гаплоидных наборов. Таким образом, особи имеют новые, различающиеся между собой генотипы. При этом достигается перекомбинация наследственных свойств родителей, что увеличивает изменчивость и дает более богатый материал для естественного отбора. Как правило, кроссинговер происходит только у женских организмов, однако, встречаются случаи кроссинговера и у гермафродитов.Гены перемешиваются благодаря слиянию гамет двух различных особей, однако генетические изменения осуществляются не только этим путем. Никакие два потомка одних и тех же родителей (если только это не идентичные близнецы) не будут абсолютно одинаковыми. Во время мейоза осуществляются два различных вида пересортировки генов. Один вид пересортировки - это результат случайного распределения разных материнских и отцовских гомологов между дочерними клетками при первом делении мейоза, каждая гамета получает свою, отличную от других выборку материнских и отцовских хромосом. Из этого следует, что клетки любой особи могут в принципе образовать 2 в степени n генетически различающихся гамет, где n - гаплоидное число хромосом. Однако на самом деле число возможных гамет неизмеримо больше из-за кросинговера (перекреста) - процесса, происходящего во время длительной профазы первого деления мейоза , когда гомологичные хромосомы обмениваются участками. У человека в каждой паре гомологичных хромосом кроссинговер происходит в среднем в 2 - 3 точках .При кроссинговере происходит разрыв двойной спирали ДНК в одной материнской и одной отцовской хроматиде, а затем получившиеся отрезки воссоединяются "наперекрест" (процесс генетической рекомбинации). Рекомбинация происходит в профазе первого деления мейоза, когда две сестринские хроматиды так тесно сближены друг с другом, что их невозможно увидеть в отдельности. Гораздо позже в этой растянутой профазе становятся ясно различимы две отдельные хроматиды каждой хромосомы. В это время видно, что они связаны своими центромерами и тесно сближены по всей длине. Два гомолога остаются связанными в тех точках, где произошел кроссинговер между отцовской и материнской хроматидами. В каждой такой точке, которую называют хиазмой , две из четырех хроматид перекрещиваются Таким образом, это морфологический результат произошедшего кроссинговера, который сам по себе недоступен для наблюдения.На этой стадии мейоза гомологи в каждой паре (или бивалент) остаются связанными друг с другом по меньшей мере одной хиазмой. Во многих бивалентах бывает большее число хиазм, так как возможны множественные перекресты между гомологами.Поскольку кроссинговер вносит возмущения в картину сцепленного наследования, его удалось использовать для картирования «групп сцепления» (хромосом). Возможность картирования была основана на предположении о том, что, чем чаще наблюдается кроссинговер между двумя генами, тем дальше друг от друга расположены эти гены в группе сцепления и тем чаще будут наблюдаться отклонения от сцепленного наследования. Первые карты хромосом были построены в 1913 г. для классического экспериментального объекта плодовой мушки Drosophila melanogaster Альфредом Стёртевантом, учеником и сотрудником Томаса Ханта Моргана.
58. Хромосомное определение пола. Наследование признаков, сцепленных с полом (на примере человека).У животных, растений и человека хромосомный механизм является начальным механизмом, определяющим пол. Согласно хромосомной теории, пол организма определяется половыми хромосомами в момент оплодотворения.XY определение полаУ одного пола ядра всех соматических клеток содержат диплоидный набор аутосом 2А и две одинаковые половые хромосомы (XX). Поэтому все гаметы этого пола содержат по одной X-хромосоме. Это гомогаметный пол. У другого пола в каждой соматической клетке, помимо диплоидного набора аутосом 2А, содержатся две разные половые хромосомы Х и Y. Поэтому у него два вида гамет: X- и Y-несущие. Это гетерогаметный пол. У большинства видов животных и растений гомогаметен женский пол, а гетерогаметен мужской. Сюда относятся млекопитающие, большинство насекомых, многие рыбы, растения и др. К видам с мужской гетерогаметностью относятся и плодовые мушки рода Drosophila. Бывает два вида XY-определения пола. Один из них — как у человека: пол зависит от наличия Y-хромосомы (если она есть, генотип самца, если нет — самки). Второй — как у представителей рода Drosophila: пол определяется по соотношению числа X-хромосом и числа аутосом.Наследование, сцепленное с полом — наследование какого-либо гена, находящегося в половых хромосомах. Наследование признаков, проявляющихся только у особей одного пола , но не определяемых генами, находящимися в половых хромосомах, называется наследованием, ограниченным полом.Наследованием, сцепленным с X-хромосомой, называют наследование генов в случае, когда мужской пол гетерогаметен и характеризуется наличием Y-хромосомы (XY), а особи женского пола гомогаметны и имеют две X-хромосомы (XX). Таким типом наследования обладают все млекопитающие (в т.ч. человек), большинство насекомых и пресмыкающихся.
59. Понятие о гене, особенности его строения в клетках прокариот.Ген - структурная и функциональная единица наследственности, контролирующая развитие определенного признака или свойств.Ген — материальный носитель наследственной информации, совокупность которых родители передают потомкам во время размножения. В настоящее время, в молекулярной биологии установлено, что гены — это участки ДНК, несущие какую-либо целостную информацию — о строении одной молекулы белка или одной молекулы РНК. Эти и другие функциональные молекулы определяют рост и функционирование организма.В то же время, каждый ген характеризуется рядом специфических регуляторных последовательностей ДНК, таких как промоторы, которые принимают непосредственное участие в регулировании проявления гена. Регуляторные последовательности могут находиться как в непосредственной близости от открытой рамки считывания, кодирующей белок, или начала последовательности РНК, как в случае с промоторами (так называемые cis-регуляторные элементы, англ. cis-regulatory elements), так и на расстоянии многих миллионов пар оснований (нуклеотидов), как в случае с энхансерами и супрессорами (иногда классифицируемые как trans-регуляторные элементы, англ. trans-regulatory elements). Таким образом, понятие гена не ограничено только кодирующим участком ДНК, а представляет собой более широкую концепцию, включающую в себя и регуляторные последовательности.Изначально термин ген появился как теоретическая единица передачи дискретной наследственной информации. История биологии помнит споры о том, какие молекулы могут являться носителями наследственной информации. Большинство исследователей считали, что такими носителями могут быть только белки, так как их строение (20 аминокислот) позволяет создать больше вариантов, чем строение ДНК, которое составлено всего из четырёх видов нуклеотидов. Позже было экспериментально доказано, что именно ДНК включает в себя наследственную информацию, что было выражено в виде центральной догмы молекулярной биологии.Гены могут подвергаться мутациям — случайным или целенаправленным изменениям последовательности нуклеотидов в цепи ДНК. Мутации могут приводить к изменению последовательности, а следовательно изменению биологических характеристик белка или РНК, которые, в свою очередь, могут иметь результатом общее или локальное изменённое или анормальное функционирование организма. Такие мутации в ряде случаев являются патогенными, так как их результатом является заболевание, или летальными на эмбриональном уровне. Однако, далеко не все изменения последовательности нуклеотидов приводят к изменению последовательности белка (благодаря эффекту вырожденности генетического кода) или к существенному изменению последовательности и не являются патогенными. В частности, геном человека характеризуется однонуклеотидными полиморфизмами и вариациями числа копий (англ. copy number variations), такими как делеции и дупликации, которые составляют около 1 % всей нуклеотидной последовательности человека[1]. Однонуклеотидные полиморфизмы, в частности, определяют различные аллели одного гена.Свойства генов:дискретность — несмешиваемость генов; стабильность — способность сохранять структуру;лабильность — способность многократно мутировать; множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм; аллельность — в генотипе диплоидных организмов только две формы гена; специфичность — каждый ген кодирует свой продукт; плейотропия — множественный эффект гена; экспрессивность — степень выраженности гена в признаке; пенетрантность — частота проявления гена в фенотипе; амплификация — увеличение количества копий гена..Классификация генов: 1) Структурные гены - уникальные компоненты генома, представляющие единственную последовательность, кодирующую определенный белок или некоторые виды РНК(тРНК, РРНК, м/яРНК) 2) Функциональные гены - регулируют работу структурных геновРазличные организмы резко отличаются по количеству ДНК, составляющей их геномы. У вирусов в зависимости от их величины и сложности размер генома колеблется от нескольких тысяч до сотен пар нуклеотидов. Гены в таких просто устроенных геномах расположены один за другим и занимают до 100% длины соответствующей нуклеиновой кислоты(РНК и ДНК). Для многих вирусов становлена полная нуклеотидная последовательность ДНК. У бактерий размер генома значительно больше. У кишечной палочки единственная нить ДНК – бактериальная хромосома состоит из 4,2х106(6 степень) пар нуклеотидов. Более половины этого количества состоит из структурных генов, т.е. генов, кодирующих определенные белки. Остальную часть бактериальной хромосомы составляют неспособные транскрибироваться нуклеотидные последовательности, функция которых не вполне ясна. Подавляющее большинство бактериальных генов уникальны, т.е. представлены в геноме один раз. Исключение составляют гены транспортных и рибосомальных РНК, которые могут повторяться десятки раз. Геном эукариот, особенно высших, резко превышает по размерам геном прокариот и достигает, как отмечалось, сотен миллионов и миллиардов пар нуклеотидов. Количество структурных генов при этом возрастает не очень сильно. Количество ДНК в геноме человека достаточно для образования примерно 2 млн. структурных генов. Реально имеющееся число оценивается как 50-100 тыс. генов, т.е. в 20-40 раз меньше того, что могло бы кодироваться геномом такого размера. Следовательно, приходится констатировать избыточность генома эукариот. Причины избыточности в настоящее время в значительной степени прояснились: во-первых, некоторые гены и последовательности нуклеотидов многократно повторены, во-вторых, в геноме существует много генетических элементов, имеющих регуляторную функцию, в-третьих, часть ДНК вообще не содержит генов
|
Согласно
современным представлениям, ген,
кодирующий синтез определенного
белка, у эукариот состоит из нескольких
обязательных элементов. Прежде всего
это обширная регуляторная зона,
оказывающая сильное влияние на
активность гена в той или иной ткани
организма на определенной стадии его
индивидуального развития. Далее
расположен непосредственно примыкающий
к кодирующим элементам гена промотор
– последовательность ДНК
длиной до 80-100 пар нуклеотидов,
ответственная за связывание
РНК-полимеразы, осуществляющей
транскрипцию
данного гена. Вслед за промотором
лежит структурная часть гена, заключающая
в себе информацию о первичной структуре
соответствующего белка.
Эта область для большинства генов
эукариот существенно короче регуляторной
зоны, однако ее длина может измеряться
тысячами пар нуклеотидов.
Важная
особенность эукариотических генов –
их прерывность. Это значит, что область
гена, кодирующая белок, состоит из
нуклеотидных последовательностей
двух типов. Одни – экзоны – это участки
ДНК,
которые несут информацию и строении
белка и
входят в состав соответствующих РНК
и белка. Другие – интроны – не кодируют
структуру белка
и в состав зрелой молекулы и-РНК не
входят, хотя и транскрибируются.
Процесс вырезания интронов – «ненужных»
участков молекулы РНК
и сращивания экзонов при образовании
и-РНК осуществляется специальными
ферментами и получил название Сплайсинг
(сшивание, сращивание). |
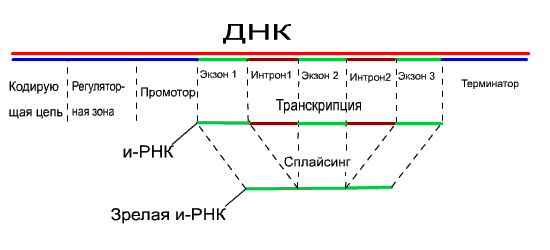 Экзоны обычно соединяются вместе в
том же порядке, в котором они распологаются
в ДНК.
Однако не абсолютно все гены эукариот
прерывисты. Иначе говоря, у некоторых
генов, подобно бактериальным, наблюдается
полное соответствие нуклеотидов
последовательности первичной структуре
кодируемых ими белков.
Таким образом, ген эукариот во многом
похож на оперон прокариот, хотя и
отличается от него более сложной и
протяженной регуляторной зоной, а
также тем, что он кодирует обычно
только один белок,
а не несколько, как оперон у бактерии.
Экзоны обычно соединяются вместе в
том же порядке, в котором они распологаются
в ДНК.
Однако не абсолютно все гены эукариот
прерывисты. Иначе говоря, у некоторых
генов, подобно бактериальным, наблюдается
полное соответствие нуклеотидов
последовательности первичной структуре
кодируемых ими белков.
Таким образом, ген эукариот во многом
похож на оперон прокариот, хотя и
отличается от него более сложной и
протяженной регуляторной зоной, а
также тем, что он кодирует обычно
только один белок,
а не несколько, как оперон у бактерии.60. Генетический код и его свойства. Структурная и функциональная классификация генов. ПримерыГенети́ческий код - это свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.В ДНК используется четыре нуклеотида — аденин (А), гуанин (G), цитозин (С), тимин (T), которые в русскоязычной литературе обозначаются буквами А, Г, Ц и Т. Эти буквы составляют алфавит генетического кода. В РНК используются те же нуклеотиды, за исключением тимина, который заменён похожим нуклеотидом — урацилом, который обозначается буквой U (У в русскоязычной литературе). В молекулах ДНК и РНКнуклеотиды выстраиваются в цепочки и, таким образом, получаются последовательности генетических букв.Для построения белков в природе используется 20 различных аминокислот. Каждый белок представляет собой цепочку или несколько цепочек аминокислот в строго определённой последовательности. Эта последовательность определяет строение белка, а следовательно все его биологические свойства. Набор аминокислот также универсален для почти всех живых организмов.Реализация генетической информации в живых клетках (то есть синтез белка, кодируемого геном) осуществляется при помощи двух матричных процессов: транскрипции (то есть синтеза иРНК на матрице ДНК) и трансляции генетического кода в аминокислотную последовательность (синтез полипептидной цепи на матрице иРНК). Для кодирования 20 аминокислот, а также сигнала «стоп», означающего конец белковой последовательности, достаточно трёх последовательных нуклеотидов. Набор из трёх нуклеотидов называется триплетом. Свойства генетического кодаТриплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон). Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно. Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов. (Не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки). Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте. (Свойство не является универсальным. Кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин)[1] Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов. Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии) (Из этого свойства также есть ряд исключений, см. таблицу в разделе «Вариации стандартного генетического кода» в данной статье). 61. Избыточность ДНК эукариот. Структурная и функциональная классификация генов.Количество ДНК в хромосомах велико и возрастает по мере усложнения организмов. Для эукариот также характерна избыточность генов. Так, у человека геном содержит число нуклеотидных пар, достаточное для образования более 2 млн. структурных генов, в то время как у человека имеется по данным 2000 года 31 тыс. всех генов.Ген — структурная и функциональная единица наследственности, контролирующая развитие определённого признака или свойства. Совокупность генов родители передают потомкам во время размножения. Однако перенос генов от родителей к потомкам не является единственным способом передачи генов.труктурные гены — уникальные компоненты генома, представляющие единственную последовательность, кодирующую определённый белок или некоторые виды РНК. (См. также статью гены домашнего хозяйства).Функциональные гены — регулируют работу структурных генов
62. Типы РКК и их Функциональная характеристика. Сходство и различие с ДНК. Формы существования и-РНК эукариот. Ферментативные Функции PНК.Рибонуклеи́новые кисло́ты (РНК) — нуклеиновые кислоты, полимеры нуклеотидов, в состав которых входят остаток ортофосфорной кислоты, рибоза (в отличие от ДНК, содержащей дезоксирибозу) и азотистые основания — аденин, цитозин, гуанин и урацил (в отличие от ДНК, содержащей вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусах.Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК), принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах.Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы) у отдельных РНК обнаружена собственная энзиматическая активность, способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами.Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.Между ДНК и РНК есть три основных отличия:ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК. Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина. ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные. Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК, мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образующих нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептидил-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК
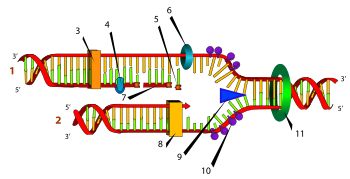
63. Репликация ДНК.Реплика́ция ДНК — это процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты, который происходит в процессе деления клетки на матрице родительской молекулы ДНК. При этом генетический материал, зашифрованный в ДНК, удваивается и делится между дочерними клетками. Репликацию ДНК осуществляет фермент ДНК-полимераза.Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у эукариот осуществляется несколькими разными ДНК-полимеразами. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный.Цепи молекулы ДНК расходятся и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются новые двуспиральные молекулы ДНК, идентичные родительской молекуле.Каждая молекула ДНК состоит из одной цепи исходной родительской молекулы и одной вновь синтезированной цепи. Такой механизм репликации называется полуконсервативным. В настоящее время этот механизм считается доказанным благодаря опытам Мэтью Мезельсона и Франклина Сталя (1958 г.). Ранее существовали и две другие модели: «консервативная» — в результате репликации одна молекула ДНК состоит только из родительских цепей, а другая — только из дочерних цепей; «дисперсионная» — все получившиеся в результате репликации молекулы ДНК состоят из цепей, одни участки которых вновь синтезированы, а другие взяты из родительской молекулы ДНК).Процесс редупликации: раскручивание спирали молекулы - отделение одной цепи от другой на части молекулы ДНК - воздействие фермента ДНК-полимеразы на молекулу - присоединение к каждой цепи ДНК комплиментарных нуклеотидов - образование двух молекул ДНК из одной.
![]()
Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК полимераза (Polα), (4) ДНК лигаза, (5) РНК праймер, (6) ДНК праймаза, (7) фрагмент Оказаки, (8) ДНК полимераза (Polδ), (9) хеликаза, (10) одиночная нить со связанными белками, (11) топоизомераза
64. Общая схема кодирования и реализации генетической информации в клетках про- и эукариот. Биологическая роль белков. Реализация генетической информации в живых клетках (то есть синтез белка, кодируемого геном) осуществляется при помощи двух матричных процессов: транскрипции (то есть синтеза мРНК на матрице ДНК) и трансляциигенетического кода в аминокислотную последовательность (синтез полипептидной цепи на мРНК). Для кодирования 20 аминокислот, а также сигнала «стоп», означающего конец белковой последовательности, достаточно трёх последовательных нуклеотидов. Набор из трёх нуклеотидов называется триплетом. Принятые сокращения, соответствующие аминокислотам и кодонам, изображены на рисунке.Белок, являясь важнейшим компонентом питания, обеспечивающим пластические и энергетические нужды организма, справедливо назван протеином, показывающим первую его роль в питании. Роль белков в питании человека трудно переоценить. Сама жизнь является одним из способов существования белковых тел. Биологическая роль белковБелок можно отнести к жизненно важным пищевым веществам, без которых невозможны жизнь, рост и развитие организма. Достаточность белка в питании и высокое его качество позволяют создать оптимальные условия внутренней среды для нормальной жизнедеятельности организма, его развития и высокой работоспособности. Белок является главной составной частью пищевого рациона, определяющей характер питания. На фоне высокого уровня белка отмечается наиболее полное проявление в организме биологических свойств других компонентов питания. Белки обеспечивают структуру и каталитические функции ферментов и гормонов, выполняют защитные функции, участвуют в образовании многих важных структур белковой природы: иммунных тел, специфических ного иммунитета, участвуют в образовании тканевых белков, таких как миозин и актин, обеспечивающих мышечные сокращения, глобина, входящего в состав гемоглобина эритроцитов крови и выполняющего важнейшую функцию дыхания. Белок, образующий зрительный пурпур (родопсин) сетчатки глаза, обеспечивает нормальное восприятие света, и др.Следует отметить, что белки определяют активность многих биологически активных веществ: витаминов, а также фосфолипидов, отвечающих за холестериновый обмен. Белки определяют активность тех витаминов, эндогенный синтез которых осуществляется из аминокислот. Например, из триптофана – витамина PР (никотиновая кислота), обмен метионина – связан с синтезом витамина U (метилметионин-сульфоний). Установлено, что белковая недостаточность может привести к недостаточности витамина С и биофлаваноидов (витамина Р). Нарушение в печени синтеза холина (группы витаминоподобных веществ) приводит к жировой инфильтрации печени.При больших физических нагрузках, а также при недостаточном поступлении жиров и углеводов белки участвуют в энергетическом обмене организма.Белки рациона определяют такие состояния, как алиментарная дистрофия, маразм, квашиоркор. Квашиоркор означает «отнятый от груди ребенок». Им заболевают дети, отнятые от груди и переведенные на углеводистое питание с резкой недостаточностью животного белка. Квашиоркор вызывает как стойкие необратимые изменения конституционального характера, так и изменения личности.Наиболее тяжелые последствия в состоянии здоровья, нередко на всю жизнь, оставляет такой вид недостаточности питания, как алиментарная дистрофия, чаще всего возникающая при отрицательном энергетическом балансе, когда в энергетические процессы включаются не только пищевые химические вещества, поступающие с пищей, но и собственные, структурные белки организма. В алиментарной дистрофии выделяют отечную и безотечную формы с явлениями или без явлений витаминной недостаточности.Может сложиться впечатление, что заболевания алиментарного характера возникают только при недостаточном поступлении белка в организм. Это не совсем так! При избыточном поступлении белка у детей первых трех месяцев жизни появляются симптомы дегидратации, гипертермии и явления обменного ацидоза, что резко увеличивает нагрузку на почки. Обычно это возникает, когда при искусственном вскармливании используют неадаптированные молочные смеси, негуманизированные типы молока.Обменные нарушения в организме могут появиться и при несбалансированности аминокислотного состава поступающих белков.
65. Экспрессия генов в клетках про- и эукариот.Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок. Экспрессия генов может регулироваться на всех стадиях процесса: и во время транскрипции, и во время трансляции, и на стадии посттрансляционных модификаций белков. Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. Экспрессия генов является субстратом для эволюционных изменений, так как контроль за временем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме.Экспрессия генов эукариот отличатся от прокариот. Уэукариот имеются три типа РНК-полимераз. РНК-полимераза1 катализирует транскрипцию рибосомальных генов. РНК-полимераза2 катализирует транскрипцию всех структурных генов. РНК-полимераза3 катализирует транскрипцию тРНК и 5S-рибосомальной РНК (катализирует образование мяРНК, присутствующие только у эукариот). Промоторный участок (промотор – участок присоединения РНК-полимеразы) у эукариот более длинный. Здесь три последовательности: одна проксимальная и две дистальные. В проксимальной основной мотив ТАТА. У эукариот это бокс Хогнеса-Гольдберга, у прокариот – прибнов бокс (ТАТААТ в проксимальном положении, ТТGАСА – в дистальном положении). Особенность дистальной последовательности заключается в том, что с одной стороны они имеют GC-основания и образуют шпильку посредством инвертированного повтора, и освобождается транскрибируемая нить РНК. Терминирующая последовательность содержит мотив ААУААА. Терминирующая последовательность расположена у конца, представленным поли-А. У эукариот любой ген представляет чередующиеся кодирующими и некодирующими последовательностями. Кодирующие – экзоны, некодирующие – интроны. Иногда внутри экзоны может быть встроен небольшой интрон. У эукариот встречаются усилители, узнаваемыми белками. Они могут быть расположены достаточно далеко от начала транскрипции. Усилитель и связанный с ним белок приближаются к участку связывания РНК-полимеразы с ДНК. Существуют "глушители", подавляющие транскрипцию. Активная транскрипция гена становится возможной после того, как на ДНК соберется крупный белковый комплекс, включающий факторы транскрипции и РНК-полимеразу. Пространственные структуры белковых факторов должны быть хорошо "подогнаны" друг к другу, обеспечивая работу гена, либо его выключение. Изучение регуляции генной активности у прокариот привело французских микробиологов Ф. Жакоба и Ж. Моно к созданию (1961) оперонной модели регуляции транскрипции. Оперон — это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований. Например, это могут быть гены, которые детерминируют синтез ферментов, участвующих в метаболизме какого-либо вещества или в синтезе какого-то компонента клетки. Оперонная модель регуляции экспрессии генов предполагает наличие единой системы регуляции у таких объединенных в один оперон структурных генов, имеющих общий промотор и оператор.Особенностью прокариот является транскрибирование мРНК со всех структурных генов оперона в виде одного полицистронного транскрипта, с которого в дальнейшем синтезируются отдельные пептиды.
66. Основные этапы биосинтеза, протекающие по матричному принципу. Участие т-РНК в синтезе белков.Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков, происходящий на рибосомах клеток живых организмов с участием молекул мРНК и тРНК.Биосинтез белка можно разделить на стадии транскрипции, процессинга и трансляции. Во время транскрипции происходит считывание генетической информации, зашифрованной в молекулах ДНК, и запись этой информации в молекулы иРНК. В ходе ряда последовательных стадий процессинга из иРНК удаляются некоторые фрагменты, ненужные в последующих стадиях, и происходит редактирование нуклеотидных последовательностей. После транспортировки кода из ядра к рибосомам происходит собственно синтез белковых молекул, путем присоединения отдельных аминокислотных остатков к растущей полипептидной цепи.Процессинг.Между транскрипцией и трансляцией молекула иРНК претерпевает ряд последовательных изменений, которые обеспечивают созревание функционирующей матрицы для синтеза полипептидной цепочки. С появлением процессинга в эукариотической клетке стало возможно комбинирование экзонов гена для получения большего разнообразия белков, кодируемым единой последовательностью нуклеотидов ДНК.Основная статья: 5'-кэп Химическая структура кэпа.При кэпировании происходит присоединение к 5'-концу транскрипта 7-метилгуанозина посредством трифосфатного моста, соединяющего их в необычной позиции 5'-5', а также метилирование рибоз двух первых нуклеотидов. Процесс кэпирования начинается ещё до окончания транскрипции молекулы пре-мРНК.Функции кэп-группы:регулирование экспорта мРНК из ядра;защита 5'-конца транскрипта от экзонуклеаз;участие в инициации трансляцииПолиаденилированиеПолиаденилирование заключается в присоединении к 3'-концу транскрипта от 100 до 200 остатков адениловой кислоты, осуществляемом специальным ферментом poly(A)-полимераза.Основная статья: СплайсингПосле полиаденилирования мРНК подвергается удалению интронов. Процесс катализируется сплайсосомой и называется сплайсингом.Трансляция (биология)Трансляция заключается в синтезе полипептидной цепи в соответствии с информацией, закодированной в матричной РНК. Аминокислотная последовательность выстраивается при помощи транспортных РНК (тРНК), которые образуют с аминокислотами комплексы — аминоацил-тРНК. Каждой аминокислоте соответствует своя тРНК, имеющая соответствуюищий антикодон, «подходящий» к кодону мРНК. Во время трансляции рибосома движется вдоль мРНК, по мере этого наращивается полипептидная цепь. Энергией биосинтез белка обеспечивается за счет АТФ.Готовая белковая молекула затем отщепляется от рибосомы и транспортируется в нужное место клетки. Для достижения своего активного состояния некоторые белки требуют дополнительной посттрансляционной модификации.
67. Понятие об опероне. Опероны про- и эукариот. Регуляция деятельности адаптивных оперонов прокариот в реакциях расщепления (на примере лактозного оперона кишечной палочки). Сплайсинг белков.Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны (гены, единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами. Такая функциональная организация позволяет эффективнее регулировать экспрессию (транскрипцию) этих генов.Концепцию оперона для прокариот предложили в 1961 году французские ученые Жакоб и Моно, за что получили Нобелевскую премию в 1965 году.Опероны по количеству цистронов делят на моно-, олиго- и полицистронные, содержащие, соответственно, только один, несколько или много цистронов (генов). Состав оперона прокариот входят структурные гены и регуляторные элементы (не путать с геном-регулятором). Структурные гены кодируют белки, осуществляющие последовательно этапы биосинтеза какого-либо вещества. Этих генов может быть один, два или несколько. Они тесно сцеплены друг с другом и, что самое главное, в ходе транскрипции работают как один единый ген: на них синтезируется одна общая молекула иРНК, которая лишь потом расщепляется на несколько иРНК, соответствующих отдельным генам. Регуляторными элементами являются следующие:— промотор — участок связывания фермента, осуществляющего транскрипцию ДНК - РНК-полимеразы. Является местом начала транскрипции. Представляет собой короткую последовательность из нескольких десятков нуклеотидов ДНК, с которой специфически связывается РНК-полимераза. Кроме того, промотор определяет, какая из двух цепей ДНК будет служить матрицей для синтеза иРНК;- оператор - участок связывания регуляторного белка;- терминатор - участок в конце оперона, сигнализирующий о прекращении транскрипции.На работу оператора данного оперона влияет самостоятельный ген-регулятор, синтезирующий соответствующий регуляторный белок. Этот ген не обязательно располагается рядом с опероном. Кроме того, один регулятор может регулировать транскрипцию нескольких оперонов. Ген-регулятор также имеет собственный промотор и терминатор. Регуляторные белки бывают двух типов: белок-репрессор или белок-активатор. Они присоединяются к специфическим нуклеотидным последовательностям ДНК оператора, что либо препятствует транскрипции генов (негативная, отрицательная регуляция), либо способствует ей (позитивная, положительная регуляция); механизмы их работы противоположны. Кроме того, на работу белков-репрессоров могут влиять вещества — эффекторы: соединяясь с репрессором, они влияют на его взаимодействие с оператором. У эукариот транскрипция осуществляется с участков, подобных оперонам прокариот и также состоящих из регуляторных и структурных генов, однако у оперонов эукариот имеется ряд особенностей.1. В состав оперона эукариот входит лишь один структурный ген (а не несколько — как у прокариот).2. Оперон эукариот почти всегда содержит только структурный ген, а прочие гены разбросаны по хромосоме или даже по разным хромосомам.3. Оперон эукариот состоит из чередующихся друг с другом значащих (экзонов) и незначащих (интронов) участков. При транскрипции вчитываются как экзоны, так и интроны, а затем в ходе процессинга происходит вырезание интронов (сплайсинг). У эукариот механизмы регуляции активности отдельных генов и генома в целом довольно сложны, и рассмотрение этих вопросов выходит далеко за рамки школьного курса биологии.Сплайсинг (от англ. splice — сращивать или склеивать концы чего-либо) — процесс вырезания определенных нуклеотидных последовательностей из молекул РНК и соединения последовательностей, сохраняющихся в «зрелой» молекуле, в ходе процессинга РНК. Наиболее часто этот процесс встречается при созревании информационной РНК (мРНК) у эукариот, при этом путём биохимических реакций с участием РНК и белков из мРНК удаляются участки, не кодирующие белок (интроны) и соединяются друг с другом кодирующие аминокислотную последовательность участки — экзоны. Таким образом незрелая пре-мРНК превращается в зрелую мРНК, с которой считываются (транслируются) белки клетки. Большинство генов прокариот, кодирующих белки, не имеют интронов, поэтому у них сплайсинг пре-мРНК встречается редко. У представителей эукариот, бактерий и архей встречается также сплайсинг транспортных РНК (тРНК) и других некодирующих РНК. 68. Регуляция деятельности адаптивных оперонов в клетках прокариот в реакциях синтеза (на примере аргининового оперона кишечной палочки).Состав оперона прокариот входят структурные гены и регуляторные элементы (не путать с геном-регулятором). Структурные гены кодируют белки, осуществляющие последовательно этапы биосинтеза какого-либо вещества. Этих генов может быть один, два или несколько. Они тесно сцеплены друг с другом и, что самое главное, в ходе транскрипции работают как один единый ген: на них синтезируется одна общая молекула иРНК, которая лишь потом расщепляется на несколько иРНК, соответствующих отдельным генам. Регуляторными элементами являются следующие:— промотор — участок связывания фермента, осуществляющего транскрипцию ДНК - РНК-полимеразы. Является местом начала транскрипции. Представляет собой короткую последовательность из нескольких десятков нуклеотидов ДНК, с которой специфически связывается РНК-полимераза. Кроме того, промотор определяет, какая из двух цепей ДНК будет служить матрицей для синтеза иРНК;- оператор - участок связывания регуляторного белка;- терминатор - участок в конце оперона, сигнализирующий о прекращении транскрипции.
69. Формы изменчивости организмов - модификационная, комбинативная, мутационная. Их значение в онтогенезе и эволюции.Изменчивость — это возникновение индивидуальных различий. На основе изменчивости организмов появляется генетическое разнообразие форм, которые в результате действия естественного отбора преобразуются в новые подвиды и виды. Различают изменчивость модификационную, или фенотипическую, и мутационную, или генотипическую.ТАБЛИЦА Сравнительная характеристика форм изменчивости (Т.Л. Богданова. Биология. Задания и упражнения. Пособие для поступающих в ВУЗы. М.,1991)Формы изменчивости Причины появления Значение ПримерыНеиаследственная модификационная (фенотипическая) Изменение условий среды, в результате чего организм изменяется в пределах нормы реакции, заданной генотипом Адаптация — приспособление к данным условиям среды, выживание, сохранение потомства Белокочанная капуста в условиях жаркого климата не образует кочана. Породы лошадей и коров, завезенных в горы, становятся низкорослымиНаследственная (генотипическая)-Мутационная.Влияние внешних и внутренних мутагенных факторов, в результате чего происходит изменение в генах и хромосомах Материал для естественного и искусственного отбора, так как мутации могут быть полезные, вредные и безразличные, доминантные и рецессивные Появление полиплоидных форм в популяции растений или у некоторых животных (насекомых, рыб) приводит к их репродуктивной изоляции и образованию новых видов, родов — микроэволюции Комбинатнвная.Возникает стихийно в рамках популяции при скрещивании, когда у потомков появляются новые комбинации генов Распространение в популяции новых наследственных изменений, которые служат материалом для отбора Появление розовых цветков при скрещивании белоцветковой и красноцветковой примул. При скрещивании белого и серого кроликов может появиться черное потомствоСоотносительная (коррелятивная) Возникает в результате свойства генов влиять на формирование не одного, а двух и более признаков Постоянство взаимосвязанных признаков, целостность организма как системы Длинноногие животные имеют длинную шею. У столовых сортов свеклы согласованно изменяется окраска корнеплода, черешков и жилок листа.Модификационная изменчивостьМодификационная изменчивость не вызывает изменений генотипа, она связана с реакцией данного, одного и того же генотипа на изменение внешней среды: в оптимальных условиях выявляется максимум возможностей, присущих данному генотипу. Так, продуктивность беспородных животных в условиях улучшенного содержания и ухода повышается (надои молока, нагул мяса). В этом случае все особи с одинаковым генотипом отвечают на внешние условия одинаково (Ч. Дарвин этот тип изменчивости назвал определенной изменчивостью). Однако другой признак — жирность молока — слабо подвержен изменениям условий среды, а масть животного — еще более устойчивый признак. Модификационная изменчивость обычно колеблется в определенных пределах. Степень варьирования признака у организма, т. е. пределы модификационной изменчивости, называется нормой реакции.Широкая норма реакции свойственна таким признакам, как удои молока, размеры листьев, окраска у некоторых бабочек; узкая норма реакции — жирности молока, яйценоскости у кур, интенсивности окраски венчиков у цветков и др.Фенотип формируется в результате взаимодействий генотипа и факторов среды. Фенотипические признаки не передаются от родителей потомкам, наследуется лишь норма реакции, т. е. характер реагирования на изменение окружающих условий. У гетерозиготных организмов при изменении условий среды можно вызвать различные проявления данного признака.Свойства модификаций: 1) ненаследуемость; 2) групповой характер изменений; 3) соотнесение изменений действию определенного фактора среды; 4) обусловленность пределов изменччивоти генотипом.Генотипическая изменчивостьГенотипическая изменчивость подразделяется на мутационную и комбинативную. Мутациями называются скачкообразные и устойчивые изменения единиц наследственности — генов, влекущие за собой изменения наследственных признаков. Термин “мутация” был впервые введен де Фризом. Мутации обязательно вызывают изменения генотипа, которые наследуются потомством и не связаны со скрещиванием и рекомбинацией генов.Классификация мутаций. Мутации можно объединять, в группы—классифицировать по характеру проявления, по месту или, по уровню их возникновения.Мутации по характеру проявления бывают доминантными и рецессивными. Мутации нередко понижают жизнеспособность или плодовитость. Мутации, резко снижающие жизнеспособность, частично или полностью останавливающие развитие, называют полулетальными а несовместимые с жизнью — летальными. Мутации подразделяют по месту их возникновения. Мутация, возникшая в половых клетках, не влияет на признаки данного организма, а проявляется только в следующем поколении. Такие мутации называют генеративными. Если изменяются гены в соматических клетках, такие мутации проявляются у данного организма и не передаются потомству при половом размножении. Но при бесполом размножении, если организм развивается из клетки или группы клеток, имеющих изменившийся — мутировавший — ген, мутации могут передаваться потомству. Такие мутации называют соматическими.Мутации классифицируют по уровню их возникновения. Существуют хромосомные и генные мутации. К мутациям относится также изменение кариотипа (изменение числа хромосом).. Полиплоидия — увеличение числа хромосом, кратное гаплоидному набору. В соответствии с этим у растений различают триплоиды (Зп), тетраплоиды (4п) и т. д. В растениеводстве известно более 500 полиплоидов (сахарная свекла, виноград, гречиха, мята, редис, лук и др.). Все они выделяются большой вегетативной массой и имеют большую хозяйственную ценность.Большое многообразие полиплоидов наблюдается в цветоводстве: если одна исходная форма в гаплоидном наборе имела 9 хромосом, то культивируемые растения этого вида могут иметь 18, 36, 54 и до 198 хромосом. Полиплоиды пблучают в результате воздействия на растения температуры, ионизирующей радиации, химических веществ (колхицин), которые разрушают веретено деления клетки. У таких растений гаметы диплоидны, а при слиянии с гаплоидными половыми клетками партнера в зиготе возникает триплоидный набор хромосом (2п + п = Зп). Такие триплоиды не образуют семян, они бесплодны, но высокоурожайны. Четные полиплоиды образуют семена.Гетероплоидия —изменение числа Хромосом, не кратное гаплоидному набору. При этом набор хромосом в клетке может быть увеличен на одну, две, три хромосомы (2п + 1; 2п + 2; 2п + 3) или уменьшен на одну хромосому (2л-1). Например, у человека с снндромом Дауна оказывается одна лишняя хромосома по 21-й паре и кариотип такого человека составляет 47 хромосом У людей с синдромом Шерешевского —Тернера (2п-1) отсутствует одна Х-хромосома и в кариотипе остается 45 хромосом. Эти и другие подобные отклонения числовых отношений в кариотипе человека сопровождаются расстройством здоровья, нарушением психики и телосложения, снижением жизнеспособности и др.Хромосомные мутации связаны с изменением структуры хромосом.Существуют следующие виды перестроек хромосом: отрыв различных участков хромосомы, удвоение отдельных фрагментов, поворот участка хромосомы на 180° или присоединение отдельного участка хромосомы к другой хромосоме. Подобное изменение влечет за собой нарушение функции генов в хромосоме и наследственных свойств организма, а иногда и его гибель.Генные мутации затрагивают структуру самого гена и влекут за собой изменение свойств организма (гемофилия, дальтонизм, альбинизм, окраска венчиков цветков и т. д.). Генные мутации возникают как в соматических, так и в половых клетках. Они могут быть доминантными и рецессивными. Первые проявляются как у гомозигот, так и. у гетерозигот, вторые — только у гомозигот. У растений возникшие соматические генные мутации сохраняются при вегетативном размножении. Мутации в половых клетках наследуются при семенном размножении растений и при половом размножении животных. Одни мутации оказывают на организм положительное действие, другие безразличны, а третьи вредны, вызывая либо гибель организма, либо ослабление его жизнеспособности (например, серповидноклеточная анемия, гемофилия у человека).При выведении новых сортов растений и штаммов микроорганизмов используют индуцированные мутации, искусственно вызываемые теми или иными мутагенными факторами (рентгеновские или ультрафиолетовые лучи, химические вещества). Затем проводят отбор полученных мутантов, сохраняя наиболее продуктивные. В нашей стране этими методами получено много хозяйственно перспективных сортов растений: неполегающие пшеницы с крупным колосом, устойчивые к заболеваниям; высокоурожайные томаты; хлопчатник с крупными коробочками и др.Свойства мутаций:1. Мутации возникают внезапно, скачкообразно.2. Мутации наследственны, т. е. стойко передаются из поколения в поколение.3. Мутации ненаправденны - мутировать может любой локус, вызывая изменения как незначительных, так и жизненно важных признаков. 4. Одни и те же мутации могут возникать повторно. 5. По своему проявлению мутации могут быть полезными и вредными, доминантными и рецессивными.Способность к мутированию — одно из свойств гена. Каждая отдельная мутация вызывается какой-то причиной, но в большинстве случаев эти причины неизвестны. Мутации связаны с изменениями во внешней среде. Это убедительно доказывается тем, что путем воздействия внешними факторами удается резко повысить их число.Комбинативная изменчивостьКомбинативная наследственная изменчивость возникает в результате обмена гомологичными участками гомоло гичных хромосом в процессе мейоза, а также как следствие независимого расхождения хромосом при мейозе и случайного их сочетания при скрещивании. Изменчивость может быть обусловлена не только мутациями, но и сочетаниями отдельных генов и хромосом, новая комбинация которых при размножении приводит к изменению определенных признаков и свойств организма. Такой тип изменчивости называют комбинативной наследственной изменчивостью. Новые комбинации генов возникают: 1) при кроссинговере, во время профазы первого мейотического деления; 2) во время независимого расхождения гомологичных хромосом в анафазе первого мейотического деления; 3) во время независимого расхождения дочерних хромосом в анафазе второго мейотического деления и 4) при слиянии разных половых клеток. Сочетание в зиготе рекомбинированных генов может привести к объединению признаков разных пород и сортов.В селекции важное значение имеет закон гомблогических рядов наследственной изменчивости, сформулированный советским ученым Н. И. Вавиловым. Он гласит: внутри разных видов и родов, генетически близких (т. е. имеющих единое происхождение), наблюдаются сходные ряды наследственной изменчивости. Такой характер изменчивости выявлен у многих злаков (рис, пшеница, овес, просо и др.), у которых сходно варьируют окраска и консистенция зерна, холодостойкость и иные качества. Зная характер наследственных изменений у одних сортов, можно предвидеть сходные изменения у родственных видов и, воздействуя на них мутагенами, вызывать у них подобные полезные изменения, что значительно облегчает получение хозяйственно ценных форм. Известны многие примеры гомологической изменчивости и у человека; например, альбинизм (дефект синтеза клетками красящего вещества) обнаружен у европейцев, негров и индейцев; среди млекопитающих — у грызунов, хищных, приматов; малорослые темнокожие люди — пигмеи — встречаются в тропических лесах экваториальной Африки, на Филиппинских островах и в джунглях полуострова Малакки; некоторые наследственные дефекты и уродства, присущие человеку, отмечены и у животных. Таких животных используют в качестве модели для изучения аналогичных дефектов у человека. Например, катаракта глаза бывает у мыши, крысы, собаки, лошади; гемофилия — у мыши и кошки, диабет — у крысы; врожденная глухота — у морской свинки, мыши, собаки; заячья губа — у мыши, собаки, свиньи и т. д. Эти наследственные дефекты — убедительное подтверждение закона гомологических рядов наследственной 70. Норма реакции. Адаптивный характер модификаций. Роль наследственности и среды в развитии, обучении и воспитании человека.Но́рма реа́кции — способность генотипа формировать в онтогенезе, в зависимости от условий среды, разные фенотипы. Она характеризует долю участия среды в реализации признака и определяет модификационную изменчивость вида. Чем шире норма реакции, тем больше влияние среды и тем меньше влияние генотипа в онтогенезе. Один и тот же ген в разных условиях среды может реализоваться в несколько проявлений признака (фенов). В каждом конкретном онтогенезе из спектра проявлений признака реализуется только один. Аналогично один и тот же генотип в разных условиях среды может реализоваться в целый спектр потенциально возможных фенотипов, но в каждом конкретном онтогенезе реализуется только один фенотип. Под наследственной нормой реакции понимают максимально возможную ширину этого спектра: чем он шире, тем шире норма реакции.МОДИФИКАЦИЯ АДАПТИВНАЯфенотипические генетические изменения признаков организма (не затрагивающие его генетической структуры), вызванные модифицирующим действием среды в чувствительных периодах его развития. М. а. сохраняются только на протяжении жизни измененного организма и передаются лишь потомству, полученному путем вегетативного размножения родительской формы.
71. Мутационная изменчивость, классификация мутаций по изменению в генотипе. Генные мутации. Роль их в патология человека. Мутационная изменчивость - изменения генов, это мутации, новые состояния генов. Они устойчивы, как и гены из которых они возникли, путем ауторепродукции и они передаются потомкам. Причины появления мутации- действия мутогенов и особенности обмена веществ.Классификация мутаций по генотипу:I.генные (кол-во последовательности нуклеотидов)- почковые мутации (касаются нуклеотидов внутри гена)1)генные ставки (появление новой пары нуклеотидов)2)выпадение пары нуклеотидов (любое выпадение носит название- делеция)3)перестановки нуклеотидов внутри (генная инверсия)4)генные дупликации (удвоение какой-либо пары или нескольких)II.хромосомные- возникают в результате перестройки хромосом. Возможны внутрихромосомные операции.1)возможна делеция (потеря участка хромосомы)2)появление лишнего, дублирующего участка (дубликация)3)инверсия (когда какой-либо участок инвертируется, поворот на 180градусов при разрыве хромосом)III.межхромосомные-возникают при перераспределении генного материала между разными хромосомами1)транслокация (возникает в мейозе, если обмениваются участки, негомологичные хромосомам)2)трансформация (1944г. Эвери описал ее у бактерий, заключается в том, что участок ДНК переносится из одной хромосомы в другую)3)трансдукция (перенос и рекомбинация генов у бактерий одного генотипа, с помощью бактериофага,к бактериям другого генотипа)1952г-работа Ледерберга IV.геномные – связаны с изменением числа хромосом в кл-ке1)полиплоидия (увеличение диплоидного числа хромосом, путем добавления целых геномных n-наборов). Имеет значение, кратность добавления наборов.Причины полиплоидии:а)нарушение мейоза,связанное с нерасхождением хромосо 2n♀ x n♂ = 3nб)удвоение хромосом в S-периоде без деления кл-ки (норма для человека)в)слияние ядер соматических кл-к2)Анеуплоидия (число хромосом изменяется внекратно набору)3)гаплоидия(уменьшение числа хромосом,половинный гаплоидный наборнегомологичных хромосом). Проявляются все рецесивные признаки.Механизмы мутаций:На ур-не ДНК:В результате ультрафиолетового воздействия в момент репликации, появляются таутомерные формы А-Ц; А-Г; Т-Г; Т-Ц.И эти отклонения будут передаваться новым цепям ДНК.На ур-не хромосом:Эти изменения происходят при неравном кроссинговере; А при кроссинговере происходит обмен равными участками. Но разрыв может произойти в несимметричных точках. В одной хромосоме может удвоится ген,а вдругом можетбыть нехвватка( Делеция 5-ой пары,синдром кошачьего мурлыканья).На ур-не генома:Связано с нарушением нормального течения митоза, возникают кл-ки с неравным кареотипом. При овогенезе у женщин старше 35 лет,при нерасхождении хромосом по 21 паре, по 13 паре,и нерасхождения половых хромосом
♀ хх
♂
xy
хх
♂
xy
 x+o
=>xo
(синдром Шаушершера)
x+o
=>xo
(синдром Шаушершера)
хх о y+o =>yo
xxy (синдром Транфельтера)
xxx (трейфомия)
Генные (точковые) мутации - это изменения числа и/или последовательности нуклеотидов в структуре ДНК (вставки, выпадения, перемещения, замещения нуклеотидов) в пределах отдельных генов, приводящие к изменению количества или качества соответствующих белковых продуктов. Эффекты генных мутаций чрезвычайно разнообразны. Большая часть мелких генных мутаций фенотипически не проявляется, поскольку они рецессивны, однако известен ряд случаев, когда изменение всего лишь одного основания в определенном гене оказывает глубокое влияние на фенотип. Одним из примеров служит серповидноклеточная анемия - заболевание, вызываемое у человека заменой основания в одном из генов, ответственных за синтез гемоглобина. Молекула дыхательного пигмента гемоглобина у взрослого человека состоит из четырех полипептидных цепей (двух (- и двух (- цепей), к которым присоединены четыре простетические группы гема.
72. Комбинативная изменчивость. Значение комбинативной изменчивости в обеспечении генотипического разнообразия.Комбинативной называют изменчивость, в основе которой лежит образование рекомбинаций, т. е. таких комбинаций генов, которых не было у родителей.В основе комбинативной изменчивости лежит половое размножение организмов, вследствие которого возникает огромное разнообразие генотипов. Практически неограниченными источниками генетической изменчивости служат три процесса: Независимое расхождение гомологичных хромосом в первом мейотическом делении. Именно независимое комбинирование хромосом при мейозе является основой третьего закона Менделя. Появление зеленых гладких и желтых морщинистых семян гороха во втором поколении от скрещивания растений с желтыми гладкими и зелеными морщинистыми семенами — пример комбинативной изменчивости. Взаимный обмен участками гомологичных хромосом, или кроссинговер. Он создает новые группы сцепления, т. е. служит важным источником генетической рекомбинации аллелей. Рекомбинантные хромосомы, оказавшись в зиготе, способствуют появлению признаков, нетипичных для каждого из родителей. Случайное сочетание гамет при оплодотворении. Эти источники комбинативной изменчивости действуют независимо и одновременно, обеспечивая при этом постоянную «перетасовку» генов, что приводит к появлению организмов с другими генотипом и фенотипом (сами гены при этом не изменяются). Однако новые комбинации генов довольно легко распадаются при передаче из поколения в поколение. Комбинативная изменчивость является важнейшим источником всего колоссального наследственного разнообразия, характерного для живых организмов. Однако перечисленные источники изменчивости не порождают существенных для выживания стабильных изменений в генотипе, которые необходимы, согласно эволюционной теории, для возникновения новых видов. Такие изменения возникают в результате мутаций.
73. Мутации соматические и генеративные, их роль в патологии человека.В настоящее время считается, что многие мутации не оказывают существенного влияния на жизнеспособность особей; такие мутации называются нейтральными. Нейтральность мутаций часто обусловлена тем, что большинство мутантных аллелей рецессивно по отношению к исходному аллелю. Однако существуют мутации, приводящие к гибели организма (летальные) или заметно снижающие его жизнеспособность (полулетальные). В определенных условиях мутации могут повышать жизнеспособность организмов (как в примере с серповидноклеточной анемией).По способности передаваться при половом размножении различают соматические и генеративные мутации. Соматические мутации не затрагивают половые клетки и не передаются потомкам. В результате соматических мутаций возникают генетическиемозаики. Генеративные мутации происходят в половых клетках и могут передаваться потомкам. При участии мутантных половых клеток образуются полностью мутантные организмы.Мутации возникают как в аутосомах, так и в половых хромосомах; соответственно различают аутосомные мутации и мутации, сцепленные с полом. Кроме того, по возможности проявления в фенотипе различают доминантные, полудоминантные и рецессивные мутации (заметим, что подавляющее большинство мутаций является рецессивными).Мутантный аллель может возвращаться в исходное состояние. Тогда первоначальная мутация называется прямой (например, переход А → а), а другая – обратной мутацией, или реверсией (например, обратный переход а → А).Мутация в соматической клетке на ранней стадии развития может вызвать наследуемое или ненаследуемое изменения фенотипа. Организм с такой мутацией называют мозаичным, потому что он содержит как нормальные клетки, так и мутантные. Мутации в соматических клетках, возможно, вызывают процессы старения , рак и другие, менее существенные изменения в организме.Соматическими называют клетки, составляющие тело (сому) многоклеточных организмов и не принимающие участия в половом размножении. Входя в состав разнообразных тканей тела, соматические клетки каждой ткани обладают специфическими структурными, метаболическими и химическими особенностями, которые приобретаются в процессе дифференцировки. Соматические и половые клетки имеют общее происхождение, так как образуются из генетически одинаковых эмбриональных клеток, которые содержат всю генетическую информацию, необходимую для образования клеток различных типов в ходе развития организма.Различные ткани многоклеточных организмов представляют собой популяции клеток, которые могут изменяться с возрастом и в зависимости от состояния организма. Входящие в состав таких популяций клетки в результате дифференцировки приобретают стабильные наследственные свойства соответствующего типа. Эта стабильность сохраняется даже при культивировании in vitro выделенных из организма соматических клеток. Многие генетические процессы, характерные для половых клеток, происходят и в соматических клетках (мутации, хромосомные перестройки, рекомбинации, расщепления, полиплоидизация и др.).Изучение наследственности и изменчивости соматических клеток необходимо для решения многих важных проблем, среди которых особо следует выделить проблему старения, патологии клеток, действия на организм различных физических факторов, а также проблему дифференцировки клеток в онтогенезе и их интеграции в ткани. Генетика соматических клеток является важным разделом цитогенетики, и в этой главе будут кратко рассмотрены ее задачи, методы и основные достижения.. Изменчивость и наследственность соматических клетокизучении наследственности соматических клеток используют обычные генетические методы, методы культуры клеток и тканей, а также пересадку тканей между разными мутантными линиями растений и животных. Проведение генетического анализа и эксперимента на соматических клетках связано с рядом трудностей, и в настоящее время продолжаются поиски наиболее рациональных методов исследования. Кроме того, для изучения изменчивости и наследственности соматических клеток широко используются цитологические методы.генетического анализа соматических клеток будет успешным, когда будут соблюдены следующие основные условия: 1) учет соматических мутаций; 2) получение потомства от одной клетки, что очень важно для любого генетического эксперимента; 3) обеспечение передачи генетической информации от одной клетки к другой. Здесь имеется в виду не только передача генетической информации от материнской клетки к дочерним в ходе деления, но и передача, аналогичная гибридизации при скрещиваниях; 4) учет расщепления в потомстве гетерозиготных соматических клеток.В настоящее время уже получены экспериментальные данные, доказывающие, что в ряде случаев возможно учесть и воспроизвести необходимые условия для проведения генетического анализа соматических клеток.Существуют такие явления, как соматический кроссинговер и соматические мутации. Эти явления показывают, что при комбинировании у гетерозиготного организма различных сцепленных мутаций можно наблюдать рекомбинации по их мозаичному проявлению в фенотипе. При воздействии же мутагенами на ранние стадии развития организма можно вызвать мозаичное проявление мутаций в тех участках тканей, которые развились из мутантных клеток. Дополнительно к сказанному ранее рассмотрим еще несколько примеров соматического кроссинговера и соматических мутаций.Подробное исследование соматического кроссинговера было проведено Понтекорво с сотрудниками (1958) на грибе Aspergillus nidulans. Этот гриб имеет гаплоидные ядра и одноядерные конидии, после прорастания которых образуются гифы, дающие мицелий. Гифы генетически различных штаммов могут сливаться, и их ядра объединяются в общем мицелии. Новый штамм представляет собой гетерокарион, так как клетки его мицелия имеют много генотипически различных ядер, лежащих в общей цитоплазме. Гетерокарион образует конидии, каждая из которых имеет одно ядро того или другого типа, что позволяет опять получать штаммы, сходные с «родительскими». Смешивая два штамма, нуждающихся в разных питательных веществах, можно получить стабильный гетерокарион. В этом случае генотипически различные ядра в общей цитоплазме дополняют друг друга, обеспечивая синтез необходимых веществ и тем самым рост гетерокариона на минимальной среде.Образованный мутантными штаммами у paba (желтый, нуждающийся в п-аминобензойной кислоте) и w ad (белый, нуждающийся в аденине) гетерокарион имеет ядра, которые различаются по гену образования пигмента и по генам, определяющим потребность в факторах роста. Такой гетерокарион образует колонию на минимальной среде, но образующиеся споры не содержат пигмента дикого типа. Лишь иногда появляются споры с зеленой пигментацией дикого типа. Эти споры прорастают на минимальной среде и дают одноцветные зеленые колонии. Они являются диплоидными и гетерозиготными по родительским хромосомам. Имеющиеся у них гены у+ и w+, определяющие синтез пигмента, дополняют друг друга (комплементация), а это позволяет идентифицировать и отобрать диплоидные клетки.Наличие двух пар генов-маркеров позволило точно определить частоту соматических рекомбинаций, происходящих в результате соматического кроссинговера, и частоту соматических мутаций. В гетерокарионах Aspergillus происходит отбор генетически благоприятных конституций, что позволяет организму приспосабливаться к меняющимся условиям. Рассматривать же такие изменения как модель изменчивости соматических клеток можно лишь в тех пределах, в каких вегетативную массу мицелия можно рассматривать как сому. Следует учитывать, что гетерокарионы – особые образования и их нельзя отождествлять с соматическими клетками высших растений и животных., например, еще удобнее, чем грибы, для модельных опытов по изучению мутационного процесса и отбора в популяциях соматических клеток. Однако и здесь при сравнениях необходима известная осторожность. Правда, существующие в настоящее время данные о соматических клетках высших организмов, в общем, не противоречат данным, полученным на одноклеточных организмах. Более того, экспериментирование на соматических клетках высших организмов в той или иной мере имитирует подходы, применяемые в генетике микроорганизмов.При изучении изменчивости и наследственности соматических клеток у высших растений и животных возникает много технических трудностей. Кроме того, диплоидность соматических клеток высших организмов препятствует непосредственному наблюдению рецессивных мутаций.Многие культурные растения подвергаются отбору на мутабильность. Мозаичность почек, например, часто связана с соматическими мутациями, что можно выявить при анализе результатов скрещивания. У вегетативно размножающихся растений соматические мутации удобно изучать на гаплоидных формах. При воздействии различными мутагенами на гаплоидные растения в отдельных соматических клетках индуцируются мутации. Если мутантные клетки дадут начало побегам, то при наличии гаплоидного набора хромосом любая мутация проявится фенотипически, даже если она рецессивна. Такие мутантные побеги можно укоренить, а затем при помощи колхицина перевести на диплоидный уровень, т. е. перевести соматическую мутацию в гомозиготное состояние. Если мутантный побег образует затем генеративные органы, то данную мутацию можно будет воспроизводить через семенное размножение. Получение таким способом гомозиготных диплоидных мутантов на основе гаплоидных форм вполне возможно у томатов, картофеля и у других растений, которые можно размножать вегетативно.Одним из эффективных методов изучения генетики соматических клеток является метод культуры клеток и тканей in vitro, основанный на создании условий, благоприятных для жизнедеятельности и деления клеток вне организма. Этот метод в принципе не отличается от генетических методов исследования микроорганизмов. Основные его преимущества заключаются в том, что он позволяет точно контролировать условия среды и генотипы отдельных клеток.Клетки каждой ткани в связи с постоянно происходящим в них мутационным процессом образуют гетерогенную популяцию. В организме клетки и ткани интегрированы в систему функционально взаимосвязанных органов. При культивировании клеток отдельных тканей in vitro контроль и регулировка со стороны организма снимаются, и популяции клеток оказываются дезинтегрированными, что делает их более пластичными в отношении наследственной изменчивости. В культуре появляются клетки с различной плоидностью, хромосомными перестройками, различными морфологическими и биохимическими мутациями, селективная ценность которых в однородной среде будет различной. Одни клетки начинают делиться быстрее, другие медленнее, а третьи вообще не делятся. В связи с этим при использовании метода культуры клеток и тканей для цитогенетических исследований важным является получение клонов клеток с известными генетическими особенностями и поддержание их в культуре.Клоны соматических клеток (колонии, происходящие от отдельных клеток), получают путем изоляции одиночных клеток при помощи микрокапилляра с последующей их инкубацией в специальных сосудах.В настоящее время существует много способов культивирования клеток, тканей и органов, растительных и животных объектов. Они различаются методами эксплантации, составом питательных сред, условиями культивирования и т. д. С этими методами можно ознакомиться в специальной литературе по культуре клеток и тканей. Наибольшие успехи достигнуты при изучении культуры клеток животных и человека, а цитогенетические исследования культуры клеток растительных тканей пока проводились в значительно меньших масштабах. Рассмотрим основные результаты исследования изменчивости и наследственности соматических клеток при использовании метода культуры клеток и тканей.Возможности получения клонов соматических клеток различных животных и человека позволили цитогенетикам начать изучение у них мутационного процесса. Это имеет большое значение для познания закономерностей возникновения различных соматических мутаций (хромосомных и генных) и для изучения особенностей передачи наследственной информации как в ряду клеточных поколений, так и от одной соматической клетки к другой, неродственной первой.В результате таких исследований было установлено, что в клетках животных и человека возникают различные мутации: морфологические, биохимические и мутации резистентности к различным инфекциям и химическим агентам.Прежде всего, в соматических клетках значительно изменяются числа хромосом, т. е. часто наблюдается полиплоидия и гетероплоидия (анеуплоидия). В ряде случаев в ходе жизни культуры количество полиплоидных клеток значительно увеличивается, т. е. они как бы вытесняют диплоидные клетки.В соматических клетках млекопитающих и человека довольно часто наблюдаются различные хромосомные аберрации (фрагментация, делеции, дупликации. инверсии, транслокации). Это характерно бывает и для культуры клеток, и для организмов. Интересно, что с возрастом организма в популяциях клеток нормальных тканей частота хромосомных аберраций увеличивается. Полиплоидизация и хромосомные перестройки, частота которых бывает значительной, указывают на большую изменчивость соматических клеток.С использованием культуры клеток значительно увеличились масштабы радиационно-генетических исследований. Изучение действия различных излучений на клеточном уровне имеет ряд существенных преимуществ по сравнению с радиационно-генетическими исследованиями на уровне организма. В результате таких исследований было установлено, что у соматических клеток под действием различных излучений возникают все виды мутаций, характерные и для половых клеток. При этом существует такая же зависимость выхода хромосомных и генных мутаций от вида излучения, дозы и мощности облучения. Таким образом, соматические клетки являются хорошей моделью для изучения индуцированного радиацией мутационного процесса.Интересные данные были получены при сравнительном изучении радиочувствительности соматических клеток in vivo и in vitro, согласно которым радиочувствителыюсть соматических клеток в культуре оказалась сходной с радиочувствительностью клеток in vivo. Это обстоятельство очень важно для работ по радиационной генетике соматических клеток.При изучении мутационного процесса в культуре соматических клеток часто используется метод селективных сред, который был разработан при изучении генетики микроорганизмов. Этот метод очень удобен для выявления мутаций резистентности к каким-либо химическим веществам. Общая схема его сводится к следующему. В питательную среду добавляется химическое вещество в такой концентрации, при которой нормальные клетки не могут развиваться и размножаться. Колонии на этой среде будут давать только мутантные клетки, которые и отбираются. Если известно количество высеянных клеток и количество полученных колоний, то можно приблизительно определить темп мутационного процесса. Селективные среды позволили получить много различных мутантных линий соматических клеток, резистентных к различным фармакологическим веществам, антибиотикам, вирусным инфекциям и химическим мутагенам. В ряде случаев удалось определить частоту спонтанного возникновения мутации резистентности в соматических клетках млекопитающих и человека. Установлено, что частоты мутирования в половых и соматических клетках существенно не различаются.В ходе изучения соматических клеток животных и человека накопились данные, позволяющие предположить существование таких способов обмена генетической информацией между соматическими клетками, как трансформация, трансдукция и гибридизация. Явления трансформации и трансдукции были открыты при генетических исследованиях микроорганизмов. Трансформация – это генотипическое изменение какого-либо бактериального штамма в результате поглощения ДНК бактерий другого штамма, а трансдукция – генетические изменения бактериальных клеток при передаче им инфицирующими их бактериофагами отдельных частей хромосом бактерий других штаммов. Эти явления хорошо изучены на микроорганизмах и обнаружены на соматических клетках человека.Например, было взято два штамма клеток костного мозга и культуре, которые различались генотипически обусловленной пищевой потребностью. Клетки одного штамма могли использовать гипоксантин в качестве источника пуринов, а клетки другого не обладали этой способностью. Первые использовались в качестве донора, а вторые являлись реципиентом. Из донорских клеток извлекалась ДНК, и ею обрабатывались клетки реципиента. Обработанные клетки помещались на селективную среду, в которой единственным источником пуринов был гипоксантин. Выжить в этих условиях могли лишь клетки, которые претерпели трансформацию и приобрели способность использовать гипоксантин. Такие клетки были получены, хотя и с небольшой частотой (примерно одна на 105 клеток). Количество жизнеспособных в данных условиях клеток увеличивалось с увеличением концентрации ДНК. Затем была проверена специфичность действия ДНК. Если реципиент обрабатывался ДНК, извлеченной из его же клеток, то трансформация отсутствовала. Следовательно, ДНК для обработки должна быть взята из клеток, которые могли использовать гипоксантин как источник пуринов. Следующим доказательством трансформации в этом случае было то, что при обработке ДНК донора ДНК-азой исчезала ее трансформирующая активность. Все это подтверждает возможность передачи генетической информации от одних соматических клеток человека к другим путем трансформации.Происходит ли подобная трансформация в организме, пока остается неясным. Многие исследователи считают, что это вполне возможно. Например, между эмбриональными клетками и в дифференцированных тканях имеют место индукционные отношения, обеспечивающие взаимовлияние клеток и тканей, но механизмы этих процессов пока неизвестны.Обмен генетической информацией между соматическими клетками может происходить не только путем трансформации, но и посредством гибридизации. Использование цитогенетического и генетического контроля позволило получить прямые доказательства переноса ядерных генов от одной клетки к другой, а также совмещения геномов двух клеток в одной с воспроизведением их в последующих клеточных поколениях. Такие работы в последние годы были проведены многими исследователями. В качестве примера рассмотрим одну из них.Для опыта (Барский и Корнеферт, 1962) использовали две линии клеток мышиной саркомы, которые различались по ряду свойств и признаков. Одна из них была высокоопухолевой, ее клетки имели веретенообразую форму и многочисленные цитоплазматические отростки. Для этой линии модальное (наиболее распространенное) число хромосом равнялось 55…56, причем все хромосомы были телоцентрическими. Другая линия была низкоопухолевой, с клетками округлой формы без цитоплазматических отростков. Модальное число хромосом этой линии составляло 62, и, кроме телоцентрических, здесь имелись метацентрические хромосомы (напомним, что в «стандартных» клетках домовой мыши 2n=40).Была заложена смешанная культура, содержащая примерно по 5 млн. клеток каждой линии, которая инкубировалась в течение 2,5 месяцев. В такой культуре появилась линия новых клеток, которые отличались от двух исходных и были названы клетками М. Эти клетки были крупнее, округлой формы и имели цитоплазматические выросты. У клеток типа М модальное число хромосом оказалось равным 100…115, среди, них были мета- и телоцентрические хромосомы. Все это подтверждало, что клетки М образовались в результате слияния клеток исходных линий, т. е. являются гибридными формами. При этом клетки М имели селективное преимущество перед исходными и вытесняли их, что упрощало их выделение из общей культуры. Полученные от М-клеток клоны культивировались в течение года; затем они утрачивали часть хромосом и видоизменялись. М-клетки совмещали наследственные свойства исходных линий. По показателю злокачественности они были сходны с клетками первой линии, т. е. во всех случаях их введения мышам подкожно вызывали развитие опухоли.Пока нет оснований утверждать, что такая гибридизация происходит внутри целостного организма, но нет оснований и для отрицания этого. Очевидно, что гибридизация соматических клеток при половом воспроизведении не имеет эволюционного значения, но для онтогенетического развития и полиморфизма клеточных популяции в тканях она может иметь исключительно важное значение.Рассмотренные в этом разделе данные свидетельствуют о важности изучения генетических процессов в соматических клетках, которые во многом оказываются сходными с половыми клетками. Соматические клетки обладают и некоторыми специфическими особенностями, которые обусловлены тем, что в них не происходит мейоза и генетические процессы осуществляются на основе митотического цикла.
74. Спонтанные и индуцированные мутации. Мутагены. Мутагенез и канцерогенез.Мутации делятся на спонтанные и индуцированные. Спонтанные мутации возникают самопроизвольно на протяжении всей жизни организма в нормальных для него условиях окружающей среды с частотой около 10 − 9 — 10 − 12 на нуклеотид за клеточную генерацию.Индуцированными мутациями называют наследуемые изменения генома, возникающие в результате тех или иных мутагенных воздействий в искусственных (экспериментальных) условиях или при неблагоприятных воздействиях окружающей среды.Мутации появляются постоянно в ходе процессов, происходящих в живой клетке. Основные процессы, приводящие к возникновению мутаций — репликация ДНК, нарушения репарации ДНК и генетическая рекомбинация.Мутагены (от мутации и греч. -genes-рождающий, рожденный), химические и физические факторы, вызывающие наследственные изменения — мутации. Впервые искусственные мутации получены в 1925 году Г. А. Надсеном и Г. С. Филипповым у дрожжей действием радиоактивного излучения радия; в 1927 году Г. Мёллер получил мутации у дрозофилы действием рентгеновских лучей. Способность химических веществ вызывать мутации (действием иода на дрозофилы) открыта в 1932 году В. В. Сахаровым.Содержание КлассификацияМутагенами могут быть различные факторы, вызывающие изменения в структуре генов, структуре и количестве хромосом. По происхождению мутагены классифицируют на эндогенные, образующиеся в процессе жизнедеятельности организма и экзогенные — все прочие факторы, в том числе и условия окружающей среды.
75. Хромосомные аберрации. Геномные мутации. Хромосомные синдромы человека.Развитие наследственных болезней зависит от хромосомных перестроек (аберраций) и изменений непосредственно в самих генах (мутаций). Наследственные задатки, заключенные в хромосомах и генах, постоянно взаимодействуют со средой и имеется несколько путей изменения наследственного материала. Прежде всего это относительно стабильны спонтанные и менее стабильные индуцированные хромосомные аберрации и геномные мутации. Представление о количественной стороне дают следующие данные: 3 — 4% зрелых половых клеток имеют хромосомные перестройки. Возникновение хромосомных перестроек, геномных мутаций и перемещение генов в наследственном материале обусловлено воздействием самых различных факторов внешней и внутренней среды (ионизирующее излучение, химические вещества, вакцины, лекарства, интоксикации, гормональные расстройства и др.). Чтобы вызвать геномную перестройку — мутацию, на ДНК должна воздействовать энергия в 2,5 — 3 электроновольта. Наиболее благоприятные условия для этого создаются во время репликации — удвоения ДНК, которое происходит под влиянием фермента полимеразы. По-видимому, поломки хотя бы в одном звене ДНК или в структуре полимеразы приводят к нарушениям в обмене веществ и могут вызвать уродства и аномалии или увеличить восприимчивость к разнообразным заболеваниям. Поэтому генетические особенности организма являются важным фактором в развитии большинства известных нам болезней. Возникающие изменения исправляются в ряде случаев особыми ферментами — репаразами (молекулярными корректорами). Лучше исправляются одиночные разрывы, затрагивающие лишь одну из цепочек молекулы ДНК, значительно хуже — двойные разрывы, когда повреждаются сразу обе цепочки ДНК. Установлена избирательная способность генов к образованию мутаций, что в свою очередь определяет относительную стабильность форм геномных заболеваний нервной системы на протяжении ряда поколений.Хромосомные болезни или хромосомные синдромы – это комплексы множественных врожденных пороков развития, вызываемых числовыми или структурными изменениями хромосом, видимыми в световой микроскоп. Нарушения в строении хромосом, изменения их количества, генные мутации могут возникать на разных этапах развития организма. Если они возникают в гаметах родителей, то аномалия будет наблюдаться во всех клетках организма (полный мутант). Если они возникают в процессе эмбрионального развития, хромосомный набор в разных клетках тела будет разным. В процессе развития появляется несколько следующих друг за другом поколений клеток с различными хромосомными наборами. При незначительном количестве аномальных клеток болезни в последующем может и не быть. Причинами хромосомных болезней являются все хромосомные мутации (отклонения в строении и функциях) и некоторые геномные мутации (изменения числа хромосом). У человека обнаружены все виды хромосомных мутаций: делеции (потеря участка хромосомы), дупликации (представление какого-либо участка хромосомы два раза), инверсии (поворот сегмента хромосомы на 180 градусов), транслокации (перенос участка хромосомы в необычное место)..Наиболее специфичные для того или иного синдрома проявления обусловлены изменениями небольших участков хромосом. Проявления хромосомных мутаций зависят от следующих главных факторов:особенностей вовлеченной в аномалию хромосомытипа аномалии (трисомия, моносомия, полная, частичная);размера недостающего (при частичной моносомии) или избыточного (при частичной трисомии) генетического материала;степени мозаичности организма по клеткам, имеющим отклонения в строении и функциях;генотипа организма; условий среды. У человека встречаются только 3 вида геномных мутаций: триплоидия (утроенное число хромосом по сравнения с нормальным числом хромосом в клетках тела), тетраплоидия (учетверенное число хромосом), анеуплоидия (содержание хромосом на одну больше или меньше) Из всех вариантов анеуплоидий встречаются только трисомии (не расхождение хромосом при делении клетки) всех клеток, кроме половых; полисомии (наличие в наборе хромосом лишних) половых клеток (три-, тетра- и пентасомии); моносомия X (отсутствие половой Х-хромосомы). Хромосомные болезни у новорожденных детей встречаются с частотой примерно 2,4 случая на 1 тыс. родившихся. Большинство хромосомных аномалий несовместимы с жизнью и эмбрионы и плоды гибнут в организме матери в основном в ранние сроки беременности. Хромосомные аномалии возникают во всех клетках тела с частотой около 2 %. В норме такие клетки уничтожаются иммунной системой, если проявляют себя чужеродно. Однако в некоторых случаях, например, при активация генов, превращающих нормальные клетки в злокачественные, хромосомные аномалии могут быть причиной злокачественного роста. Общим для всех форм хромосомных болезней является множественность поражения. Это черепно-лицевые поражения, врожденные пороки развития систем органов, замедленные внутриутробные и постнатальные рост и развитие, отставание в психическом развитии, нарушения функций нервной, иммунной и эндокринной систем. Механизм возникновения и развития хромосомных болезней не ясен. Окончательный диагноз хромосомных болезней устанавливается цитогенетическими методамиСиндром Шереше́вского — Тёрнера — хромосомная болезнь, сопровождающаяся характерными аномалиями физического развития, низкорослостью и половым инфантилизмом.Основные сведенияВпервые эта болезнь как наследственная была описана в 1925 г. Н. А. Шерешевским, который считал, что она обусловлена недоразвитием половых желез и передней доли гипофиза и сочетается с врожденными пороками внутреннего развития. В 1938 г. Тернер выделил характерную для этого симптомокомплекса триаду симптомов: половой инфантилизм, кожные крыловидные складки на боковых поверхностях шеи и деформацию локтевых суставов. В России этот синдром принято называть синдромом Шерешевского — Тернера.Четкой связи возникновения синдрома Тернера с возрастом и какими-либо заболеваниями родителей не выявлено. Однако беременности обычно осложняются токсикозом, угрозой выкидыша, а роды часто бывают преждевременными и патологическими. Особенности беременностей и родов, заканчивающихся рождением ребенка с синдромом Тернера, — следствие хромосомной патологии плода. Нарушение формирования половых желез при синдроме Тернера обусловлено отсутствием или структурными дефектами одной половой хромосомы (X-хромосомы).У эмбриона первичные половые клетки закладываются почти в нормальном количестве, но во второй половине беременности происходит их быстрая инволюция (обратное развитие), и к моменту рождения ребенка количество фолликулов в яичнике по сравнению с нормой резко уменьшено или они полностью отсутствуют. Это приводит к выраженной недостаточности женских половых гормонов, половому недоразвитию, у большинства больных — к первичной аменорее (отсутствию менструаций) и бесплодию. Возникшие хромосомные нарушения являются причиной возникновения пороков развития. Возможно также, что сопутствующие аутосомные мутации играют определенную роль в появлении пороков развития, поскольку существуют состояния, сходные с синдромом Тернера, но без видимой хромосомной патологии и полового недоразвития.При синдроме Тернера половые железы обычно представляют собой недифференцированные соединительнотканные тяжи, не содержащие элементов гонад. Реже встречаются рудименты яичников и элементы яичек, а также рудименты семявыносящего протока. Другие патологические данные соответствуют особенностям клинических проявлений. Наиболее важны изменения костно-суставной системы — укорочение пястных и плюсневых костей, аплазия (отсутствие) фаланг пальцев, деформация лучезапястного сустава, остеопороз позвонков. Рентгенологически при синдроме Тернера турецкое седло и кости свода черепа обычно не изменены. Отмечаются пороки сердца и крупных сосудов (коарктация аорты, незаращение боталлова протока, незаращение межжелудочковой перегородки, сужение устья аорты), пороки развития почек. Проявляются рецессивные гены дальтонизма и других заболеваний.Синдром Шерешевского-Тёрнера встречается много реже, чем трисомия Х, синдром Клайнфельтера (ХХУ, ХХХУ), а так же ХУУ, что указывает на наличие сильного отбора против гамет, не содержащих половых хромосом, или против зигот ХО. Это предположение подтверждается достаточно часто наблюдемой моносомией Х среди спонтанно абортированных зародышей. В связи с этим допускается, что выжившие зиготы ХО являются результатом не мейотического, а митотического нерасхождения, или утраты X-хромосомы на ранних стадиях развития. Моносомии УО у человека не обнаружено. Популяционная частота 1:1500.Клиническая картина и диагностикаСкладки кожи в области шеи — характерный признак болезни. На фото: девочка до и после пластической операцииОтставание больных с синдромом Тернера в физическом развитии заметно уже с рождения. Примерно у 15 % больных задержка наблюдается в период полового созревания. Для доношенных новорожденных характерна малая длина (42—48 см) и масса тела (2500—2800 г и менее). Характерными признаками синдрома Тернера при рождении являются избыток кожи на шее и другие пороки развития, особенно костно-суставной и сердечно-сосудистой систем, «лицо сфинкса», лимфостаз (застой лимфы, клинически проявляющийся крупными отеками). Для новорожденного характерны общее беспокойство, нарушение сосательного рефлекса, срыгивание фонтаном, рвота. В раннем возрасте у части больных отмечают задержку психического и речевого развития, что свидетельствует о патологии развития нервной системы. Наиболее характерным признаком является низкорослость. Рост больных не превышает 135—145 см, масса тела часто избыточна. У больных с синдромом Тернера патологические признаки по частоте встречаемости распределяются следующим образом: низкорослость (98 %), общая диспластичность (неправильное телосложение) (92 %), бочкообразная грудная клетка (75 %), укорочение шеи (63 %), низкий рост волос на шее (57 %), высокое «готическое» нёбо (56 %), крыловидные складки кожи в области шеи (46 %), деформация ушных раковин (46 %), укорочение метакарпальных и метатарзальных костей и аплазия фаланг (46 %), деформация локтевых суставов (36 %), множественные пигментные родинки (35 %), лимфостаз (24 %), пороки сердца и крупных сосудов (22 %), повышенное артериальное давление (17 %).Половое недоразвитие при синдроме Тернера отличается определенным своеобразием. Нередкими признаками являются геродермия (патологическая атрофия кожи, напоминающая старческую) и мошонкообразный вид больших половых губ, высокая промежность, недоразвитие малых половых губ, девственной плевы и клитора, воронкообразный вход во влагалище. Молочные железы у большинства больных не развиты, соски низко расположены. Вторичное оволосение появляется спонтанно и бывает скудным. Матка недоразвита. Половые железы не развиты и представлены обычно соединительной тканью. При синдроме Тернера отмечается склонность к повышению артериального давления у лиц молодого возраста и к ожирению с нарушением питания тканей.Интеллект у большинства больных с синдромом Тернера практически сохранен, однако частота олигофрении все же выше. В психическом статусе больных с синдромом Тернера главную роль играет своеобразный психический инфантилизм с эйфорией при хорошей практической приспособляемости и социальной адаптации.Диагноз синдрома Тернера основывается на характерных клинических особенностях, определении полового хроматина (вещества клеточного ядра) и исследовании кариотипа (хромосомного набора). Дифференциальный диагноз проводится с нанизмом (карликовостью), для исключения которого проводится определение содержания гормонов гипофиза в крови, особенно гонадотропинов.ЛечениеНа первом этапе терапия заключается в стимуляции роста тела анаболическими стероидами и другими анаболическими препаратами. Лечение следует проводить минимальными эффективными дозами анаболических стероидов с перерывами при регулярном гинекологическом контроле. Главным видом терапии больных является эстрогенизация (назначение женских половых гормонов), которую следует проводить с 14-16 лет. Лечение приводит к феминизации телосложения, развитию женских вторичных половых признаков, улучшает трофику (питание) половых путей, уменьшает повышенную активность гипоталамо-гипофизарной системы. Лечение следует проводить в течение всего детородного возраста больных. При синдроме Тернера у мужчин применяется заместительная терапия мужскими половыми гормонами.Прогноз для жизни при синдроме Тернера благоприятный, исключение составляют больные с тяжелыми врожденными пороками сердца и крупных сосудов и почечной гипертензией. Лечение женскими половыми гормонами делает больных способными к семейной жизни, однако абсолютное большинство из них остаются бесплодными.Кроме этого в последнее время проводится терапия соматотропином или человеческим гормоном роста
76. Значение генетики для медицины. Цитогенетические методы изучения наследственности человека.Прогресс в развитии медицины и общества приводит к относительному возрастанию доли генетически обусловленной патологии в заболеваемости, смертности, социальной дизадаптации (инвалидизации).Известно более 4000 нозологических форм наследственных болезней. Около 5-5,5% детей рождаются с наследственными или врождёнными болезнями. Генегика человека, или медицинская генетика, изучает явления наследственности и изменчивости в различных популяциях людей, особенности проявления и развития нормальных (физических, творческих, интеллектуальных способностей) и патологических признаков, зависимость заболеваний от генетической предопределенности и условий окружающей среды, в том числе от социальных условий жизни. Формирование медицинской генетики началось в 30-е гг. XX в., когда стали появляться факты, подтверждающие, что наследование признаков у человека подчиняется тем же закономерностям, что и у других живых организмов.Задачей медицинской генетики является выявление, изучение, профилактика и лечение наследственных болезней, а также разработка путей предотвращения вредного воздействия факторов среды на наследственность человека.Цитогенетичесий метод основан на микроскопическом исследовании структуры хромосом у здоровых и больных людей. Цитогенетический контроль применяют при диагностике ряда наследственных заболеваний, связанных с явлениями анеуплои-дии и различными хромосомными перестройками. Он позволяет также изучать старение тканей на основе исследований возрастной динамики структуры клеток, устанавливать мутагенное действие факторов внешней среды на человека и т. д.В последние годы цитогенетический метод приобрел большое значение в связи с возможностями генетического анализа человека, которые открыла гибридизация соматических клеток в культуре. Получение межвидовых гибридов клеток (например, человека и мыши) позволяет в значительной степени приблизиться к решению проблем, связанных с невозможностью направленных скрещиваний, локализовать ген в определенной хромосоме, установить группу сцепления для ряда признаков и т. д. Объединение генеалогического метода с цитогенетическим, а также с новейшими методами генной инженерии значительно ускорило процесс картирования генов у человека.
77. Человек как объект генетического анализа, методы изучения наследственности человека и их возможности.Исследования генетики человека встречают большие трудности, связанные с невозможностью произвольного скрещивания, поздним наступлением половой зрелости, малым числом потомков в каждой семье, невозможностью уравнивания условий жизни для потомков.Все, что известно в настоящее время в области генетики человека получено с помощью генеалогического, цитогенетического близнецового, онтогенетического и популяционного методов исследования.Генеалогический метод – анализ закономерности наследования признаков человека на основе составления родословной – генеалогии.Анализ родословной показывает, что признак пробанда передается из поколения в поколение особям любого пола, т.е. наследуется как доминантный аутосомный.Болезни: кондродистрофическая карликовость, катаракта глаз, хрупкость костей.Применение этого метода показало, что такой признак, как гемофилия, наследуется своеобразно. Волосатые уши – признак, передающийся от отца к сыну.Цитогенетический метод – цитологический анализ кариотипа человека в норме и патологии. Кариотип – набор всех хромосом у человека и отображение его в идеограмме графически.Близнецовый метод. Близнецы – одновременно родившиеся особи у одноплодных животных. Близнецы могут быть однояйцевыми и разнояйцевыми.Дал возможность выяснить наследственную предрасположенность человека к ряду заболеваний (шизофрения, туберкулез)Онтогенетический метод используется для выяснения механизма развития наследственных заболеваний в онтогенезе.Известно, что некоторые наследственные болезни проявляются не только у гомозигот, но и в стертой форме у гетерозигот. В настоящее время усиленно разрабатываются методы выявления таких гетерозигот. Так гетерозиготный носитель Гена фенилкетонурии может быть определен введением в кровь фенилаллонина и последующим определение его уровня в плазме крови. В норме у гомозигот по доминантной аллели уровень фенилаллонина при этом не изменяется. У гетерозигот же по данной аллели, внешне здоровых людей, обнаруживается повышенное содержание в крови фенилаллонина.Популяционный метод позволяет изучать распространение отдельных генов или хромосомных аномалий в человеческих популяциях. Этот метод основывается на данных демографической статистики. Популяционный анализ интересен и тем, что он помогает понять динамику генетической структуры различных популяций и способствует выявлению связей между ними.
78. Кариотип и идиограмма человека в норме и патологии.Кариоти́п — совокупность признаков (число, размеры, форма и т. д.) полного набора хромосом, присущий клеткам данного биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Кариотипом иногда также называют и визуальное представление полного хромосомного набора (кариограммы). Внешний вид хромосом существенно меняется в течение клеточного цикла: в течение интерфазы хромосомы локализованы в ядре, как правило, деспирализованы и труднодоступны для наблюдения, поэтому для определения кариотипа используются клетки в одной из стадий их деления — метафазе митоза.Для процедуры определения кариотипа могут быть использованы любые популяции делящихся клеток, для определения человеческого кариотипа используется либо одноядерные лейкоциты, извлечённые из пробы крови, деление которых провоцируется добавлением митогенов, либо культуры клеток, интенсивно делящихся в норме (фибробласты кожи, клетки костного мозга). Обогащение популяции клеточной культуры производится остановкой деления клеток на стадии метафазы митоза добавлением колхицина — алкалоида, блокирующего образование микротрубочек и «растягивание» хромосом к полюсам деления клетки и препятствующего тем самым завершению митоза.Полученные клетки в стадии метафазы фиксируются, окрашиваются и фотографируются под микроскопом; из набора получившихся фотографий формируются т. н. систематизированный кариотип — нумерованный набор пар гомологичных хромосом (аутосом), изображения хромосом при этом ориентируются вертикально короткими плечами вверх, их нумерация производится в порядке убывания размеров, пара половых хромосом помещается в конец набора (см. Рис. 1).Исторически первые недетализованные кариотипы, позволявшие проводить классификацию по морфологии хромосом получались окраской по Романовскому — Гимзе, однако дальнейшая детализация структуры хромосом в кариотипах стала возможой с появлением методик дифференциального окрашивания хромосом.Для получения классического кариотипа используется окраска хромосом различными красителями или их смесями: в силу различий в связывании красителя с различными участками хромосом окрашивание происходит неравномерно и образуется характерная полосчатая структура (комплекс поперечных меток, англ. banding), отражающая линейную неоднородность хромосомы и специфичная для гомологичных пар хромосом и их участков (за исключением полиморфных районов, локализуются различные аллельные варианты генов). Первый метод окраски хромосом, позволяющий получить такие высокодетализированные изображения, был разработан шведским цитологом Касперссоном (Q-окрашивание) [1] Используются и другие красители, такие методики получили общее название дифференциального окрашивания хромосом:[2]Q-окрашивание — окрашивание по Касперссону акрихин-ипритом с исследованием под флуоресцентным микроскопом. Чаще всего применяется для исследования Y-хромосом (быстрое определения генетического пола, выявление транслокаций между X- и Y-хромосомами или между Y-хромосомой и аутосомами, скрининг мозаицизма с участием Y-хромосом)G-окрашивание — модифицированное окрашивание по Романовскому — Гимзе. Чувствительность выше, чем у Q-окрашивания, поэтому используется как стандартный метод цитогенетического анализа. Применяется при выявлении небольших аберраций и маркерных хромосом (сегментированных иначе, чем нормальные гомологичные хромосомы)R-окрашивание — используется акридиновый оранжевый и подобные красители, при этом окрашиваются участки хромосом, нечувствительные к G-окрашиванию. Используется для выявления деталей гомологичных G- или Q-негативных участков сестринских хроматид или гомологичных хромосом.C-окрашивание — применяется для анализа центромерных районов хромосом, содержащих конститутивный гетерохроматин и вариабельной дистальной части Y-хромосомы.T-окрашивание — применяют для анализа теломерных районов хромосом.В последнее время используется методика т. н. спектрального кариотипирования, состоящая в окрашивании хромосом набором флуоресцентных красителей, связывающихся со специфическими областями хромосом[3]. В результате такого окрашивания гомологичные пары хромосом приобретают идентичные спектральные характеристики, что не только существенно облегчает выявление таких пар, но и облегчает обнаружение межхромосомных транслокаций, то есть перемещений участков между хромосомами — транслоцированные участки имеют спектр, отличающийся от спектра остальной хромосомы.Сравнение комплексов поперечных меток в классической кариотипии или участков со специфичными спектральными характеристиками позволяет идентифицировать как гомологичные хромосомы, так и отдельные их участки, что позволяет детально определять хромосомные аберрации — внутри- и межхромосомные перестройки, сопровождающиеся нарушением порядка фрагментов хромосом (делеции, дупликации, инверсии, транслокации). Такой анализ имеет большое значение в медицинской практике, позволяя диагностировать ряд хромосомных заболеваний, вызванных как грубыми нарушениями кариотипов (нарушение числа хромосом), так и нарушением хромосомной структуры или множественностью клеточных кариотипов в организме (мозаицизмом).В качестве примера можно привести следующие кариотипы:нормальный (видовой) кариотип домашнего кота: 38, XYиндивидуальный кариотип лошади с «лишней» X-хромосомой (трисомия по X-хромосоме): 65, XXXиндивидуальный кариотип домашней свиньи с делецией (потерей участка) длинного плеча (q) 10-й хромосомы: 38, XX, 10q-индивидуальный кариотип мужчины с транслокацией 21-х участков короткого (p) и длинного плеч (q) 1-й и 3-й хромосом и делецией 22-го участка длинного плеча (q) 9-й хромосомы (приведён на Рис. 3): 46, XY, t(1;3)(p21;q21), del(9)(q22)Поскольку нормальные кариотипы являются видоспецифичными, то разрабатываются и поддерживаются стандартные описания кариотипов различных видов животных и растений, в первую очередь домашних и лабораторных животных и растений[6].Аномальные кариотипы и хромосомные болезниНарушения нормального кариотипа у человека возникают на ранних стадиях развития организма: в случае, если такое нарушение возникает при гаметогенезе, в котором продуцируются половые клетки родителей, кариотип зиготы, образовавшейся при их слиянии, также оказывается нарушенным. При дальнейшем делении такой зиготы все клетки эмбриона и развившегося из него организма обладают одинаковым аномальным кариотипом.Однако нарушения кариотипа могут возникнуть и на ранних стадиях дробления зиготы, развившийся из такой зиготы организм содержит несколько линий клеток (клеточных клонов) с различными кариотипами, такая множественность кариотипов всего организма или отдельных его органов именуется мозаицизмом.Как правило, нарушения кариотипа у человека сопровождаются множественными пороками развития; большинство таких аномалий несовместимо с жизнью и приводят к самопроизвольным абортам на ранних стадиях беременности. Однако достаточно большое число плодов (~2.5 %[источник не указан 216 дней]) с аномальными кариотипами донашивается до окончания беременности.
79. Болезни, связанные с нерасхождением половых хромосом и их диагностика. Синдром Клайнфельтера — генетическое заболевание.Клиническая картина синдрома описана в 1942 году в работах Гарри Клайнфельтера и Фуллера Олбрайта[1]. Генетической особенностью этого синдрома является разнообразие цитогенетических вариантов и их сочетаний (мозаицизм). Обнаружено несколько типов полисомии по хромосомам X и Y у лиц мужского пола: 47, XXY; 47, XYY; 48, XXXY; 48, XYYY; 48 XXYY; 49 XXXXY; 49 XXXYY. Наиболее распространен синдром Клайнфельтера (47, XXY). Общая частота его колеблется в пределах 1 на 500—700 новорождённых мальчиков. Синдром Клайнфельтера обычно клинически проявляется лишь после полового созревания и поэтому диагностируется относительно поздно. К началу полового созревания формируются характерные пропорции тела: больные часто оказываются выше сверстников, но в отличие от типичного евнухоидизма размах рук у них редко превышает длину тела, ноги заметно длиннее туловища. Кроме того, некоторые дети с данным синдромом могут испытывать трудности в учёбе и в выражении своих мыслей. В подростковом возрасте синдром чаще всего проявляется увеличением грудных желез, хотя в некоторых случаях этот признак может и отсутствовать. Также необходимо отметить что у 60-75 % подростков пубертатного возраста также отмечается увеличение грудных желез — пубертатная гинекомастия, которая, однако, самостоятельно проходит в течение 2-х лет, в то время как у пациентов с синдромом Клайнфельтера гинекомастия сохраняется на всю жизнь. Гинекомастия у пациентов с синдромом Клайнфельтера двусторонняя и, как правило, безболезненная. Диагностика синдрома Клайнфельтера несложна. Распознать заболевание можно даже по внешнему виду больных и по изменениям со стороны вторичных половых признаков. Евнухоидный тип телосложения обусловлен циркуляцией в крови повышенной концентрации фолликулостимулирующего гормона, являющегося неотъемлемой частью гормональной регуляции менструальной функции женского организма. Кроме того, в биохимическом анализе крови часто отмечается абсолютный дефицит андростерона и тестостерона. Поэтому данные изменения являются крайне ценными в диагностике синдрома Клайнфельтера. Поскольку заболевание в детском возрасте практически никак себя не проявляет, вовремя выявить больных удается очень редко, что, в свою очередь, делает весьма сомнительным прогноз лечения. Наиболее эффективным способом свести к минимуму проявления синдрома Клайнфельтера является курсовая гормональная терапия в виде инъекций метилтестостерона - синтетического аналога тестостерона. Наиболее видимые результаты от такого лечения достигаются, если синдром Клайнфельтера был выявлен в возрасте до 10 лет.Синдро́м Шереше́вского — Те́рнера — хромосомная болезнь, сопровождающаяся характерными аномалиями физического развития, низкорослостью и половым инфантилизмом. Моносомия по Х-хромосоме (ХО). Отставание больных с синдромом Тернера в физическом развитии заметно уже с рождения. Примерно у 15 % больных задержка наблюдается в период полового созревания. Для доношенных новорожденных характерна малая длина (42—48 см) и масса тела (2500—2800 г и менее). Характерными признаками синдрома Тернера при рождении являются избыток кожи на шее и другие пороки развития, особенно костно-суставной и сердечно-сосудистой систем, «лицо сфинкса», лимфостаз (застой лимфы, клинически проявляющийся крупными отеками). Для новорожденного характерны общее беспокойство, нарушение сосательного рефлекса, срыгиваниефонтаном, рвота. В раннем возрасте у части больных отмечают задержку психического и речевого развития, что свидетельствует о патологии развития нервной системы. Наиболее характерным признаком является низкорослость. Рост больных не превышает 135—145 см, масса тела часто избыточная.При синдроме Тернера патологические признаки по частоте встречаемости распределяются следующим образом: низкорослость (98%), общая диспластичность (неправильное телосложение) (92%), бочкообразная грудная клетка (75%), укорочение шеи (63%), низкий рост волос на шее (57%), высокое «готическое» нёбо (56%), крыловидные складки кожи в области шеи (46%), деформация ушных раковин (46%), укорочение метакарпальных и метатарзальных костей и аплазия фаланг (46%), деформация локтевых суставов (36%), множественные пигментные родинки (35%), лимфостаз (24%), пороки сердца и крупных сосудов (22%), повышенное артериальное давление (17%).Половое недоразвитие при синдроме Тернера отличается определённым своеобразием. Нередкими признаками являются геродермия (патологическая атрофия кожи, напоминающая старческую) и мошонкообразный видбольших половых губ, высокая промежность, недоразвитие малых половых губ, девственной плевы и клитора, воронкообразный вход во влагалище. Молочные железы у большинства больных не развиты, соски низко расположены. Вторичное оволосение появляется спонтанно и бывает скудным. Матка недоразвита. Половые железы не развиты и представлены обычно соединительной тканью. При синдроме Тернера отмечается склонность к повышению артериального давления у лиц молодого возраста и к ожирению с нарушением питания тканей.Интеллект у большинства больных с синдромом Тернера практически сохранен, однако частота олигофрении все же выше. В психическом статусе больных с синдромом Тернера главную роль играет своеобразный психический инфантилизм с эйфорией при хорошей практической приспособляемости и социальной адаптации.Диагноз синдрома Тернера основывается на характерных клинических особенностях, определении полового хроматина (вещества клеточного ядра) и исследовании кариотипа (хромосомного набора). Дифференциальный диагноз проводится с нанизмом (карликовостью), для исключения которого проводится определение содержания гормонов гипофиза в крови, особенно гонадотропинов.
80. Хромосомные синдромы человека, связанные с нерасхождением аутосом и аберрацией хромосом.Синдром Дауна (трисомия по 21 хромасоме),Синдром патау (трисомия по 13 хромосоме),синдром Лекена(делеция короткого плеча 5 хромосомы),син-м Клайнфельтера(синдром ХХУ),синдром тернера(моносомия хромосомы х) 81. Понятие об энзимопатиях (на примере нарушения обмена фенилаланина). Примеры энзимопатий.Энзимопатии (ферментопатии) в широком смысле слова — патологические изменения активности ферментов. В более узком смысле этим термином обозначают наследственные заболевания, при которых вследствие изменения активности ферментов нарушается течение соответствующих биохимических реакций в организме и развиваются болезни обмена веществ. Приобретенные энзимопатии чаще всего отмечаются у детей после тяжелых, хронически протекающих заболеваний (например, при дизентерии может развиться недостаточность дисахаридаз кишечника). Состояние новорожденных при энзимопатии часто ошибочно расценивается как последствия тяжелой родовой травмы.О наследственных энзимопатиях следует думать при отставании ребенка в психофизическом развитии и при упорной дистрофии. Энзимопатии могут сопровождаться явлениями токсического симптомокомплекса, характеризоваться признаками упорного рахита, остеодистрофий, поражения почек. Причиной энзимопатий служат мутации генов, ответственных за синтез белковой молекулы энзима, которые выражаются в выпадении матрицы нуклеиновых кислот или в изменении их структуры. Возможны следующие варианты нарушений синтеза ферментов: 1) фермент не синтезируется совсем, так как отсутствует нуклеиновая кислота — матрица; 2) нарушена последовательность аминокислот в молекуле энзима; 3) отсутствует или неправильно синтезируется кофермент; 4) активность энзима изменена в связи с аномалиями в других ферментных системах; 5) блокада может быть обусловлена генетически предопределенным синтезом веществ, инактивирующих энзим. Отсутствие или сниженная активность фермента создает предпосылки для накопления в организме компонентов, предшествующих блокированной реакции или образующихся в результате ее извращения и нередко оказывающих токсическое влияние. Такое положение создается при наследственных болезнях обмена аминокислот, например фенилпировиноградная олигофрения (см.). Нарушение процессов гликогенолиза сопровождается накоплением гликогена в клетках, развитием гликогенной болезни (см.). Патологические проявления нарушенного обмена могут возникнуть в связи с недостатком продуктов, образующихся при нормальном течении реакции. В качестве примера можно привести недостаток глюкозы при одном из типов гликогенной болезни, причем степень гипогликемии во многом определяет тяжесть клинической картины
82. Генеалогический метод изучения наследственности человека и его возможности. Примеры болезней, сцепленных с Х-хромосомой. Генеалогический метод относится к наиболее универсальным методам в медицинской генетике. Этот метод помог установить закономерности наследования очень большого числа самых различных признаков у человека, как нормальных, подобных цвету глаз, цвету и форме волос и т.п., так и сопутствующих наследственным болезням. Генеалогический метод - метод родословных. Он основан на составлении и анализе родословных, при помощи чего возможно прослеживание болезни (или признака) в семье или роду. В медицинской генетике этот метод называется клинико-генеалогическим. Суть метода сводится к выявлению родословных связей и прослеживанию патологического признака среди дальних и близких прямых и непрямых родственников. С его помощью может быть установлена наследственная обусловленность изучаемого признака, а также тип его наследования. Этот метод позволяет изучить интенсивность мутационного процесса, оценить экспрессивность и пенетрантность аллеля. Он складывается из двух этапов: составления родословной и генеалогического анализа. При составлении родословной исходным является человек - пробанд, родословную которого изучают. Как правило, это и есть больной, или носитель признака, наследование которого необходимо изучить. Родословная может собираться по одному или нескольким признакам. В последнем случае может быть выявлен сцепленный характер их наследования, что используется при составлении хромосомных карт. В зависимости от цели исследования родословная может быть полной или ограниченной. Необходимо все же стремится к наиболее полному составлению родословных, а для этого необходимы сведения не менее чем о 3-4 поколениях семьи пробанда. Составление родословной сопровождается краткой записью о каждом члене родословной с точной характеристикой его родства по отношению к пробанду (легенда родословной). Необходимо также отмечать обследованных и необследованных на наличие исследуемого признака. Получение сведений о родственниках - непростая задача. Пациенты зачастую не знают о болезнях родственников или предоставляют неверные сведения. Для получения более точных сведений применяют анкетирование, а иногда и полное клиническое и лабораторно-генетическое обследование родственников. При анализе родословных в первую очередь необходимо установление наследственного характера признака. Если в родословной встречается один и тот же патологический признак несколько раз (на протяжении нескольких поколений), то, вероятно, он имеет наследственную природу. Далее необходимо установить тип наследования (аутосомно-доминантный, аутосомно-рецессивный, Х-сцепленный доминантный или рецессивный, Y-сцепленный). Определение типа наследования в конкретной родословной является серьезной генетической задачей, для ее решения врач должен иметь специальную подготовку.Болезни, связ с х-хромасомой. Тяжесть заболевания зависит от пола. Полные формы болезни проявляются преимущественно у мужчин, поскольку они гемизиготны по генам, локализованным на Х-хромосоме. Если мутация затрагивает рецессивный сцепленный с Х-хромосомой ген ( XR-болезнь ), то гетерозиготные женщины здоровы, но являются носительницами гена (а гомозиготы в большинстве случаев летальны). Если мутация затрагивает доминантный сцепленный с Х-хромосомой ген ( XD-болезнь ), то у гетерозиготных женщин болезнь проявляется в легкой форме (а гомозиготы летальны). Важнейшее свойство болезней, сцепленных с Х-хромосомой , - невозможность их передачи от отца к сыну (поскольку сын наследует Y-, а не Х-хромосому отца). К Х-сцепленным рецессивным болезням относятся, например, синдром Леша-Найхана , недостаточность Г-6-ФД , тестикулярная феминизация , синдром Гунтера ( мукополисахаридоз типа II ). Распространенная Х-сцепленная рецессивная болезнь - дальтонизм . Им страдают 8% белых мужчин, поэтому и гомозиготные женщины с этим заболеванием не редкость.
83. Понятие о близнецах. Близнецовый метод и его возможности.Близнецы́ — дети одной матери, развившиеся в течение одной беременности и появившиеся на свет в результате одних родов практически одновременно. Выделяют два основных типа близнецов. Первый тип: монозиготные (гомозиготные). Второй тип: дизиготные (гетерозиготные). Монозиготные (однояйцевые) идентичные близнецы образуются из одной зиготы (одной яйцеклетки, оплодотворенной одним сперматозоидом), разделившейся на стадии дробления на две (или более) части. Они обладают одинаковыми генотипами. Монозиготные идентичные близнецы всегда одного пола и обладают очень большим портретным сходством. Дизиготные близнецы развиваются в том случае, если две яйцеклетки оплодотворены двумя сперматозоидами. Естественно, дизиготные близнецы имеют различные генотипы. Они сходны между собой не более, чем братья и сестры, так как имеют около 50 % идентичных генов. Общая частота рождения близнецов составляет примерно 1 %, из них около 1/3 приходится на монозиготных близнецов [источник не указан 221 день]. Дизиготные близнецы не обязательно зачаты во время одного полового акта, разница может составлять несколько дней. Интересно что в редких случаях могут родится дизиготные близнецы от разных отцов. Это явление называется суперфекундация. Иногда дизиготные близнецы имеют общую сросшуюся плаценту.Как дизиготные, так и монозиготные близнецы бывают не только двойняшками, но и тройняшками, четверней и так далее вплоть до 9 детей. Полярные близнецы крайне мало изучены, но, вероятно, могут быть только двойней. Также зафиксированы случаи, когда в тройне рождались к примеру 2 идентичных близнеца и один не идентичный.Близнецовый метод. Этот метод заключается в изучении закономерностей наследования признаков в парах одно- и двуяйцевых близнецов. Он предложен в 1875 г. Гальтоном первоначально для оценки роли наследственности и среды в развитии психических свойств человека. В настоящее время этот метод широко применяют в изучении наследственности и изменчивости у человека для определения соотносительной роли наследственности и среды в формировании различных признаков, как нормальных, так и патологических. Он позволяет выявить наследственный характер признака, определить пенетрантность аллеля, оценить эффективность действия на организм некоторых внешних факторов (лекарственных препаратов, обучения, воспитания).Суть метода заключается в сравнении проявления признака в разных группах близнецов при учете сходства или различия их генотипов. Монозиготные близнецы, развивающиеся из одной оплодотворенной яйцеклетки, генетически идентичны, так как имеют 100% общих генов. Поэтому среди монозиготных близнецов наблюдается высокий процент конкордантных пар, в которых признак развивается у обоих близнецов. Сравнение монозиготных близнецов, воспитывающихся в разных условиях постэмбрионального периода, позволяет выявить признаки, в формировании которых существенная роль принадлежит факторам среды. По этим признакам между близнецами наблюдается дискордантность, т.е. различия. Напротив, сохранение сходства между близнецами, несмотря на различия условий их существования, свидетельствует о наследственной обусловленности признака.
84. Принципы профилактики наследственных болезней.Почти все заболевания зависят от наследственной предрасположенности человека. Иными словами, в зависимости от того, какие признаки унаследовал человек от родителей, его шансы на заболевание отдельными болезнями могут различаться. Среди прочих болезней выделяют такие, которые целиком (или почти полностью) зависят от наследственных факторов. Эти болезни называют наследственными. Их можно предотвратить или уменьшить вероятность их проявления, если принимать специальные меры.По степени угрозы (риска) повторного проявления в семье наследственных заболеваний их подразделяют на 3 группы:1. заболевания с высокой степенью генетического риска (1 : 4), к которым относятся болезни с аутосомно-доминантным, аутосомно-рецессивным и сцепленным с полом типом наследования;2. заболевания с умеренной степенью генетического риска (менее 1 : 10); к ним относятся наследственные заболевания, вызванные свежими мутациями, а также хромосомные болезни и заболевания с полигенным типом наследования, то есть значительная часть врождённых уродств и наследственных заболеваний, развивающихся на генетически неблагоприятном фоне;3. болезни, характеризующиеся незначительным риском повторного проявления или полным отсутствием риска.Одним из направлений профилактики наследственных заболеваний является их ранняя диагностика, определение риска наследственных заболеваний. 85. Принцип составления и анализа родословныхГенеалогический метод относится к наиболее универсальным методам в медицинской генетике. Этот метод помог установить закономерности наследования очень большого числа самых различных признаков у человека, как нормальных, подобных цвету глаз, цвету и форме волос и т.п., так и сопутствующих наследственным болезням. Генеалогический метод - метод родословных. Он основан на составлении и анализе родословных, при помощи чего возможно прослеживание болезни (или признака) в семье или роду. В медицинской генетике этот метод называется клинико-генеалогическим. Суть метода сводится к выявлению родословных связей и прослеживанию патологического признака среди дальних и близких прямых и непрямых родственников. С его помощью может быть установлена наследственная обусловленность изучаемого признака, а также тип его наследования. Этот метод позволяет изучить интенсивность мутационного процесса, оценить экспрессивность и пенетрантность аллеля. Он складывается из двух этапов: составления родословной и генеалогического анализа. При составлении родословной исходным является человек - пробанд, родословную которого изучают. Как правило, это и есть больной, или носитель признака, наследование которого необходимо изучить. Родословная может собираться по одному или нескольким признакам. В последнем случае может быть выявлен сцепленный характер их наследования, что используется при составлении хромосомных карт. В зависимости от цели исследования родословная может быть полной или ограниченной. Необходимо все же стремится к наиболее полному составлению родословных, а для этого необходимы сведения не менее чем о 3-4 поколениях семьи пробанда. Составление родословной сопровождается краткой записью о каждом члене родословной с точной характеристикой его родства по отношению к пробанду (легенда родословной). Необходимо также отмечать обследованных и необследованных на наличие исследуемого признака. Получение сведений о родственниках - непростая задача. Пациенты зачастую не знают о болезнях родственников или предоставляют неверные сведения. Для получения более точных сведений применяют анкетирование, а иногда и полное клиническое и лабораторно-генетическое обследование родственников. При анализе родословных в первую очередь необходимо установление наследственного характера признака. Если в родословной встречается один и тот же патологический признак несколько раз (на протяжении нескольких поколений), то, вероятно, он имеет наследственную природу. Далее необходимо установить тип наследования (аутосомно-доминантный, аутосомно-рецессивный, Х-сцепленный доминантный или рецессивный, Y-сцепленный). Определение типа наследования в конкретной родословной является серьезной генетической задачей, для ее решения врач должен иметь специальную подготовку.
86. Положение человека в системе животного мира. Качественное своеобразие человека.Еще в античное время человек признавался «родственником» животных. К. Линней в своей «Системе природы» поместил его вместе с высшими и низшими обезьянами в один отряд приматов. Ч. Дарвин на многочисленных примерах в специальном труде «Происхождение человека и половой отбор» показал близкое родство человека с высшими антропоидными обезьянами.Человек разумный (Homo sapiens) относится к типу Хордовые, подтипу Позвоночные, классу Млекопитающие, подклассу плацентарные, отряду приматы, семейству гоминиды.С хордовыми человека роднит: наличие хорды на ранних эмбриональных стадиях, нервной трубки, лежащей над хордой, жаберных щелей в стенках глотки, сердца на брюшной стороне под пищеварительным фактом.Принадлежность человека к подтипу позвоночных определяется замещением хорды позвоночником, развитым черепом и челюстным аппаратом, двумя парами конечностей, головным мозгом, состоящим из пяти отделов.Наличие волос на поверхности тела, пяти отделов позвоночника, сальных, потовых и млечных желез, диафрагмы, четырехка-мерного сердца, сильно развитая кора головного мозга и теплокровность свидетельствуют о принадлежности человека к классу млекопитающих.Развитие плода в теле матери и питание его через плаценту — особенности, характерные для подкласса плацентарных.Наличие передних конечностей хватательного типа (первый палец противопоставлен остальным), хорошо развитых ключиц, ногтей на пальцах, одной пары сосков млечных желез, замена в онтогенезе молочных зубов на постоянные, рождение, как правило, одного детеныша позволяют отнести человека к приматам.Более частные признаки, такие как сходная структура мозгового и лицевого отделов черепа, хорошо развитые лобные доли головного мозга, большое число извилин на полушариях головного мозга, наличие аппендикса, исчезновение хвостового отдела позвоночника, развитие мимической мускулатуры, четыре основные группы крови, сходные резус-факторы и другие признаки, сближают человека с человекообразными обезьянами. Антропоиды также болеют многими инфекционными болезнями, присущими человеку (туберкулез, брюшной тиф, детский паралич, дизентерия, СПИД и др.). У шимпанзе встречается болезнь Дауна, возникновение которой, как и у человека, связано с присутствием в кариотипе животного третьей хромосомы по 21 -й паре. Близость человека к антропоидам прослеживается и по другим признакам.В то же время между человеком и животными, в том числе и человекообразными обезьянами, существуют коренные отличия. Только человек имеет истинное прямохождение. В силу вертикального положения скелет человека имеет четыре резких изгиба позвоночника, опорную сводчатую стопу с сильно развитым большим пальцем, плоскую грудную клетку.Гибкая кисть руки — органа труда — способна выполнять самые разнообразные и высокоточные движения. Мозговой отдел черепа значительно преобладает над лицевым. Площадь коры больших полушарий и объем головного мозга значительно выше, чем у человекообразных обезьян. Человеку присуще сознание и образное мышление, с чем связана такая деятельность, как конструирование, живопись, литература, наука. Наконец, только люди могут общаться друг с другом при помощи речи. Эти особенности строения, жизнедеятельности и поведения человека —результат эволюции его животных предков.Качественное своеобразие эволюции человека заключается в том, что ее движущими силами были не только биологические, но и социальные факторы, причем именно последние имели решающее значение в процессе становления человека и продолжают играть ведущую роль в развитии современного человеческого общества. 87. Доказательства животного происхождения человека (анатомо- физиологические, биохимические, генетические).Еще в античное время человек признавался «родственником» животных. К. Линней в своей «Системе природы» поместил его вместе с высшими и низшими обезьянами в один отряд приматов. Ч. Дарвин на многочисленных примерах в специальном труде «Происхождение человека и половой отбор» показал близкое родство человека с высшими антропоидными обезьянами. Человек разумный (Homo sapiens) относится к типу Хордовые, подтипу Позвоночные, классу Млекопитающие, подклассу плацентарные, отряду приматы, семейству гоминиды. С хордовыми человека роднит: наличие хорды на ранних эмбриональных стадиях, нервной трубки, лежащей над хордой, жаберных щелей в стенках глотки, сердца на брюшной стороне под пищеварительным фактом. Принадлежность человека к подтипу позвоночных определяется замещением хорды позвоночником, развитым черепом и челюстным аппаратом, двумя парами конечностей, головным мозгом, состоящим из пяти отделов. Наличие волос на поверхности тела, пяти отделов позвоночника, сальных, потовых и млечных желез, диафрагмы, четырехка-мерного сердца, сильно развитая кора головного мозга и теплокровность свидетельствуют о принадлежности человека к классу млекопитающих. Развитие плода в теле матери и питание его через плаценту — особенности, характерные для подкласса плацентарных. Наличие передних конечностей хватательного типа (первый палец противопоставлен остальным), хорошо развитых ключиц, ногтей на пальцах, одной пары сосков млечных желез, замена в онтогенезе молочных зубов на постоянные, рождение, как правило, одного детеныша позволяют отнести человека к приматам. Более частные признаки, такие как сходная структура мозгового и лицевого отделов черепа, хорошо развитые лобные доли головного мозга, большое число извилин на полушариях головного мозга, наличие аппендикса, исчезновение хвостового отдела позвоночника, развитие мимической мускулатуры, четыре основные группы крови, сходные резус-факторы и другие признаки, сближают человека с человекообразными обезьянами. Антропоиды также болеют многими инфекционными болезнями, присущими человеку (туберкулез, брюшной тиф, детский паралич, дизентерия, СПИД и др.). У шимпанзе встречается болезнь Дауна, возникновение которой, как и у человека, связано с присутствием в кариотипе животного третьей хромосомы по 21 -й паре. Близость человека к антропоидам прослеживается и по другим признакам. В то же время между человеком и животными, в том числе и человекообразными обезьянами, существуют коренные отличия. Только человек имеет истинное прямохождение. В силу вертикального положения скелет человека имеет четыре резких изгиба позвоночника, опорную сводчатую стопу с сильно развитым большим пальцем, плоскую грудную клетку. Гибкая кисть руки — органа труда — способна выполнять самые разнообразные и высокоточные движения. Мозговой отдел черепа значительно преобладает над лицевым. Площадь коры больших полушарий и объем головного мозга значительно выше, чем у человекообразных обезьян. Человеку присуще сознание и образное мышление, с чем связана такая деятельность, как конструирование, живопись, литература, наука. Наконец, только люди могут общаться друг с другом при помощи речи. Эти особенности строения, жизнедеятельности и поведения человека —результат эволюции его животных предков.Эволюция приматов до человекаИзменения средней температуры Земли за последние 500 млн летПриматы — одна из старейших групп современных плацентарных млекопитающих. Эволюционная история приматов может быть прослежена примерно на 90 млн лет назад, когда Приматоморфы разделились на Приматов и Шерстокрылов. 85 млн. лет назад Сухоносые обезьяны отделились от Мокроносых. Около 80 млн. л.н. разошлись линии Долгопятообразных и Обезьянообразных, а Лемуры отделились от Лориобразных. Останки древнейших приматов известны лишь из палеоценовых и эоценовых отложений Северной Америки, Евразии и Африки (Плезиадаписы, Антрасимия, Нотарктус, Дарвиниус (Ида) и др). Только для Пургаториуса предполагается верхнемеловое время.После глобального похолодания, когда около 30 млн лет назад, в раннем олигоцене, Антарктида начала покрываться льдом, приматы вымерли повсеместно кроме Африки, Америки и юга Азии. Одним из выживших был Грифопитек — ископаемая обезьяна, жившая на территории современной Германии и Турции около 16,5 млн лет назад, на 1,5 млн лет раньше, чем подобные виды появились в Африке. Возможно, первые человекообразные обезьяны также появились не в Африке, а в Евразии. С другой стороны, высказываются предположения, что предки гоминид мигрировали в Евразию из Африки около 17 млн лет назад, когда эти континенты некоторое время были соединены, прежде чем вновь разделились при расширении Средиземного моря. В начале миоцена климат снова стал тёплым, и они могли процветать в Евразии, после чего ареал обитания одного из них, дриопитека, распространился из Европы или западной части Азии в Африку[3].Возможно, что уцелевшая после климатических изменений, тропическая популяция обезьян, хорошо представленная в слоях верхнего эоцена и нижнего олигоцена Файюмского оазиса в Египте (биретия, каранисия, сахарогалаго, протеопитек, египтопитек, катопитек и др.), дала начало всем ныне существующим приматам — лемурам Мадагаскара, лори Юго-Восточной Азии, африканским галаго, широконосым обезьянам Нового Света и узконосым обезьянам Старого Света (человекообразные обезьяны и мартышкообразные).В раннем миоцене примитивные узконосые обезьяны Восточной Африки пережили длительный период видообразования. Среди выделяемых ныне видов и родов обезьян этой эпохи известны, в частности, камойяпитек, моротопитек, лимнопитек, проконсул, афропитек, кениапитек, чорорапитек, экваториус, отавипитек, ньянцапитек, викториапитек, дендропитек, все из Восточной Африки, а также ореопитек, который обитал в Италии около 9 млн лет назад и европейские — пиеролапитек, анойяпитек, дриопитек. При сравнении ДНК современных обезьян было показано, что гиббоны отделились от общего ствола гоминид ~ 18 млн лет назад, а орангутаны — около 14 млн лет назад. За исключением дендропитека останки ископаемых гиббонов науке неизвестны, и их происхождение остается неясным. Ископаемым прото-орангутаном считается сивапитек, обитавший в Азии около 12—10 млн лет назад и хорапитек из Таиланда.Предполагается, что видами, близкими к общему предку горилл, шимпанзе и людей, были накалипитек из Кении и уранопитек с Балканского полуострова. По данным молекулярной биологии, около 7 — 8 млн лет назад сначала гориллы, а потом и шимпанзе отделились от предков людей. ДНК шимпанзе на 98,4 % идентична человеческой. Из-за влажного климата тропических лесов, в кислых почвах которого кости плохо сохраняются, а также отчасти из-за невнимания исследователей, сосредоточенных преимущественно на поиске предков человека, ископаемых горилл и шимпанзе до сих пор практически не найдено.В середине миоцена климат снова стал холоднее и засушливее, что спровоцировало новое массовое вымирание фауны. Однако гоминины, как и многие другие виды (антилопы, гиены, собаки, свиньи, слоны, лошади) пережили климатические изменения и сумели к ним адаптироваться. Их дальнейшая эволюция привела к образованию множества новых видов, из которых самыми древними сейчас считаются Sahelanthropus tchadensis (7 млн лет назад) и Orrorin tugenensis (6 млн лет назад). За ними, в частности, последовали:Ardipithecus (5.8-4.4 млн лет назад), с видами Ar. kadabba и Ar. ramidus;Australopithecus (4-2 млн лет назад), с видами Au. anamensis, Au. afarensis, Au. africanus, Au. bahrelghazali и Au. garhi;Paranthropus (3-1.2 млн лет назад), с видами P. aethiopicus, P. boisei и P. robustus;Kenyanthropus (3 млн лет назад), вид Kenyanthropus platyopsHomo (от 2 млн лет назад), с видами Homo habilis (или Australopithecus habilis), Homo rudolfensis, Homo ergaster, Homo georgicus, Homo antecessor, Homo cepranensis, Homo erectus, Homo heidelbergensis, Homo rhodesiensis, Homo neanderthalensis, Homo sapiens idaltu, Homo sapiens sapiens, Homo floresiensis.
88. Основные этапы антропогенеза. Ископаемые предки человека. Соотносительная роль биологических и социальных факторов в эволюции человека.Огромное значение в развитии науки о происхождении человека имеют вопросы, подробно рассмотренные Энгельсом в его известной работе „Роль труда в процессе превращения обезьяны в человека" (1876). В ней Энгельс убедительно раскрыл картину формирования человека под влиянием труда и наметил определенную последовательность в развитии существенных органов нашего тела.Начальным этапом процесса выделения человека из животного мира Энгельс считает усвоение какой-то необычайно развитой породой человекообразных (антропоморфных) обезьян прямой походки (прямохождения). „Этим был сделан решающий шаг для перехода от обезьяны к человеку" .При прямой походке рука освободилась для совершения трудовых операций.Однако только у человека рука стала вполне свободной и могла совершенствоваться в ловкости, гибкости и мастерстве. С данного момента и началась ее бурная эволюция по пути все большего приспособления к трудовым операциям. Приобретенные человеком навыки и свойства передавались по наследству и закреплялись в последующих поколениях.Таким образом, рука не только орган, но и продукт труда. При этом имеются в виду больше функциональные, чем анатомические изменения органа. Энгельс подчеркивает, что общее расположение костей и мышц на руке одинаково у человека и у обезьяны, но человеческая рука может производить сотни работ, совершенно недоступных обезьяне. Он также отмечает, что под влиянием развития рук и их деятельности произошли изменения и в организме человека.Труд расширял кругозор людей, сплачивал их в более тесные коллективы, общение в которых привело к появлению и развитию речи. Это вполне понятно: у формировавшихся людей в трудовом процессе постоянно появлялась „потребность что-то сказать друг другу. Потребность создала себе свой орган: неразвитая гортань обезьяны медленно, но неуклонно преобразовывалась путем модуляции для все более развитой модуляции, а органы рта постепенно научались произносить один членораздельный звук за другим".Под влиянием труда и языка постепенно совершенствовался мозг человека. Он, в свою очередь, тоже оказывал соответствующее влияние на язык и труд. Вместе с человеком рождалось человеческое общество, резко отличавшееся от стада обезьян, в первую очередь трудом и общением.Трудовая деятельность человека как охотника и рыболова привела его к постоянному потреблению мяса - более питательного продукта, чем растительная пища. Это способствовало необычайному сокращению процесса пищеварения, что позволило увеличить расход энергии на другие жизненные функции. Кроме того, мозг получил гораздо больше необходимых для своего развития веществ. Огромное значение в процессе эволюции человека имели использование огня и приручение животных. Необычайно важным следствием трудовой деятельности явилось расширение области расселения человека: он приспособился жить в разных, нередко очень трудных, природных и климатических условиях, а с течением времени научился даже изменять природу в своих интересах.Свою трудовую теорию происхождения человека Энгельс сформулировал в тот период, когда историческая наука располагала еще небольшим количеством достоверных фактов. Все последующие находки ископаемых людей, орудий труда и предметов быта подтверждают и дополняют эти выводы одного из гениальных основоположников марксизма.Современная наука о человеке (антропология) располагает большим фактическим материалом, накопленным в последарвиновский период. Уже на основе вполне реальных находок наука продолжает изучение процесса становления человека. Как же сегодня выглядит в общем виде схема эволюции от животных к человеку? Примерно около 60 миллионов лет назад на Земле из насекомоядных млекопитающих в результате влияния природной среды и естественного отбора развились полуобезьяны, которые затем довольно быстро разделились на две ветви. Одна из них вела к широконосым обезьянам, а другая к узконосым. Вторая ветвь и привела в конце концов к человеку.Прежде чем рассматривать ее эволюцию, отметим, что современные человекообразные обезьяны отнюдь не предки человека, хотя они и появились на Земле раньше человека (более 30 миллионов лет назад). Человекообразные тоже произошли от полуобезьян в середине третичного периода (начало его отстоит от нашего времени приблизительно на 70 миллионов лет). За несколько десятков миллионов лет полуобезьяны постепенно превратились в древесных обезьян, из которых в ходе эволюции возникли виды высших человекоподобных (антропоморфных) обезьян. Одни из древнейших антропоморфных обезьян стали предками современных шимпанзе и горилл, а другие положили начало линии, которая привела к человеку.По многим признакам древнейшие человекоподобные были сходны с современными человекообразными обезьянами, в особенности с шимпанзе и гориллой. Вместе с тем у них, однако, имелись и существенные отличия. Так, руки человекоподобных существ были короче, а ноги длиннее, чем у современных обезьян; отличались они и размерами мозга, и положением клыков.Важное значение имеют находки, сделанные в городе ЭльФайюма, расположенном в 20 километрах южнее Каира (Египет). Из них особенно интересно для ученых животное парапитек (предобезьяна) Фраасов. От этого (или от очень близкого ему) существа и ведет начало ветвь к узконосым обезьянам. Следующий этап появление проплиопитеков (предков более развитых обезьян). По сравнению со своими ближайшими предшественниками они представляют собой определенный шаг вперед и являются прародителями рода плиопитековых (более развитых обезьян). Плиопитеки в свою очередь дали начало роду древесных обезьян дриопитеков. Под этим названием объединяют большую группу животных, которые, в отличие от своих предшественников, имели одинаковую основную схему строения нижних коренных зубов. Ученые считают, что дриопитеки уже весьма близки к той форме, от которой впоследствии развились некоторые современные человекообразные обезьяны и человек.Когда мы говорим о происхождении дриопитеков и их дальнейшей эволюции, то имеем в виду, что развитие человекообразных обезьян на нашей планете шло во многих местах. Причем дриопитеки далеко не единственная известная ученым сегодня ископаемая форма человекообразных обезьян. Есть и другие подобные находки, например сивапитек в Индии, удабнопитек в Грузии. А раньше в тропических лесах, возможно, жили и другие высшие и низшие обезьяны, костные останки которых наверняка лежат гдето в третичных отложениях Евразиатского материка. Однако для эволюции человека и удабнопитек, и сивапитек не имеют такого важного значения, как дриопитек. Мы упомянули их только для того, чтобы показать, как трудно было ученым разобраться во всех этих ответвлениях и выделить основную линию развития человека. И все же, несмотря на все препятствия, наука уже выявила основные вехи формирования человека из мира животных. Как это происходило?Одним из важнейших факторов развития всего живого на Земле служили изменения природной среды. Особенно сильно воздействие их сказывалось на сравнительно недавно возникших организмах. Приспособление к новым условиям на первых порах не вызывает какихлибо ощутимых физиологических изменений, но со временем оно приводит к явно заметным сдвигам в строении тела. В конечном счете такие факторы оказали свое влияние на эволюцию обезьяны, превращение ее в человека.Свыше 20 миллионов лет назад дриопитеки разделились на две группы. Первые почти без изменений дожили до современности и навсегда остались в царстве животных (гориллы, шимпанзе). Судьба вторых иная: они сначала обитали на окраине лесов, а позже оказались в лесостепных районах с очень небольшим количеством леса. Перемещение их было вызвано тем, что в третичном периоде произошло некоторое похолодание климата. Оно явилось предвестником приближающейся эпохи оледенения четвертичного периода. В результате похолодания площади, занятые джунглями, сокращались и отступали к югу. Часть дриопитеков переселялась в эти леса. А оставшиеся на старых местах вынуждены были приспосабливаться к жизни в травянистой степи с отдельными группами деревьев. Перемены природных условий вызывали, таким образом, изменения и в образе жизни дриопитеков.Так, предки этих человекообразных обезьян издавна привыкли питаться плодами и ягодами, молодыми побегами, корнями и клубнями. Иногда они прибавляли к своему столу насекомых, птичьи яйца, червей, личинок, мелких млекопитающих. Причем большую часть жизни они проводили на деревьях, где сооружали себе и своим детенышам убежища, спасались от хищников. Теперь же дриопитеки вынуждены были, во-первых, постепенно перейти от лазания по деревьям и хождения на четвереньках к прямохождению по ровной местности, во-вторых, изменить ассортимент добываемых продуктов питания.Сегодня многие ученые склоняются к мысли, что наиболее вероятным предком существ, развивавшихся в направлении человека, является рамапитек. Останки его были найдены в Индии. (Находку назвали „рамапитек" в честь индийского бога Рамы: Рама и "питекос" греческое слово означает ..обезьяна".) Это, по сути, точка, в которой пути эволюции человека и высших обезьян разошлись. Рамапитек, по-видимому, жил в редком лесу и некоторую часть времени еще проводил на деревьях.Прямохождение через многие миллионы лет привело к специализации конечностей человекоподобных обезьян. Задние конечности распрямились в коленном суставе, кости удлинились, окрепли, пальцы укоротились, выработалась упругая сводчатая стопа. Став упругой, стопа изменила походку: уменьшились толчки при хождении. Менялась форма и положение пяточной кости, большой палец ноги стал толще и примкнул к остальным; развились сильные икроножные мышцы. А передние конечности оказались свободными. Ими можно было хватать и удерживать добычу, брать камень или палку и т. п. Фигура обезьяны все более выпрямлялась; руки стали укорачиваться, плечи расправляться. В результате этого процесса происходили изменения и в строении кистей рук: развивался большой палец; движения руки стали более пластичными. В конце концов рука оказалась органом удивительно хорошо приспособленным к труду.Прямохождение вызывало и многие другие изменения в организме наших далеких предков. Все процессы подобного рода совершались постепенно на протяжении большого периода времени. И не исключено, что при этом возникали свои сложности и затруднения. Сегодня тут ясно главное: большинство особенностей в строении тела человека связано с его прямохождением. И это, кстати, одно из самых веских доказательств, что нашими предками были крупные человекообразные обезьяны.Огромная роль в совершавшихся изменениях, очевидно, принадлежит и образу питания. Недостаток растительной пищи в степных или полустепных зонах нужно было както компенсировать. Выход был найден в употреблении все большего количества мяса, в результате чего возникла необходимость в охоте на животных. Охота, особенно на крупных животных, в свою очередь, требовала смекалки, хитрости, а главное, объединения усилий отдельных особей. Так начали складываться сперва стада, затем сообщества человекоподобных обезьян.Наши предки не отличались ни особой силой, ни острыми когтями и клыками. Скорость бега их тоже была невелика. Защищаться и охотиться человекоподобным обезьянам помогала их стадная жизнь. Причем с освобождением передних конечностей они умножали свою силу, используя палки, большие кости и камни. А это был уже, по существу, процесс перехода к зачаточным формам труда. Он возникал и развивался во многих стадах и постоянно упрочивался, становился жизненно необходимым, закреплялся в потомстве. Основное здесь заключалось в том, что эти животные постепенно переходили от употребления случайно найденных палок или камней к выбору более удобных, а затем и изготовляемых, хотя и примитивных, орудий.Коллективная охота и использование орудий труда требовали новых, более совершенных способов передачи информации друг другу. Поначалу это, очевидно, были нечленораздельные выкрики, рычание и т. п. Потом появились уже членораздельные сигналы, обозначавшие вполне определенные предметы или действия.И наконец, коллективная жизнь и труд, общение в стаде обусловили еще одно важнейшее качество развитой мозг, что в дальнейшем привело к возникновению и развитию человеческого мышления. Зарождение и развитие этого свойства оказывало огромное влияние на эволюцию разных видов человекоподобных существ (антропоидов): оно позволяло антропоидам успешно приспосабливаться к изменяющимся условиям.Так в общих чертах выглядит схема развития гоминидов представителей группы человекообразных обезьян, у которых уже появились признаки, свойственные человеку.На нашей Земле примерно от двух до пяти миллионов лет назад обитало несколько видов человекообразных обезьян. Ученым сегодня известны многие останки одного из представителей сравнительно развитых обезьян австралопитека (южная обезьяна). Впервые его костные останки были обнаружены в 1924 году в Африке.Австралопитеки по величине равны примерно современному павиану. В строении тела австралопитеков проглядывали уже некоторые признаки будущего человека: тело их было выпрямлено, они постоянно передвигались на двух ногах, руки их были свободными. Мозг достаточно велик около 600 кубических сантиметров.К прямым предкам современного человека австралопитеков относят не только из-за телесного сходства с ним. Эти существа уже могли изготавливать орудия труда, хотя еще чрезвычайно примитивные. Изготовление орудий первый важнейший поворотный пункт в истории человечества. Ведь палку или камень, как известно, нередко используют и обезьяны. А вот для того, чтобы целенаправленно изготовлять орудия труда, требуется уже определенный уровень сознания. При этом, однако, надо иметь в виду, что биологические факторы в эволюции человека по-прежнему играли главную роль.До недавнего времени считалось, что австралопитеки жили на Земле около миллиона лет назад. Но в 1960 году научный мир облетела весть об открытии английского археолога Л. Лики. При раскопках в Олдовайском ущелье (Восточная Африка) он обнаружил останки древнего существа, которое назвал зинджантропом (восточно африканский человек). Размеры мозга зинджантропа не превышали размеров мозга австралопитеков. Но некоторые особенности в строении тела показывали, что он более древняя форма на пути эволюции человека. Древнее были и те геологические отложения, в которых обнаружены останки ископаемого существа.Вскоре Лики нашел еще одно существо. Его останки залегали на большей глубине, чем останки ранее открытых существ. В строении конечностей, ключицы и кистей презинджантроп так ученый назвал новую находку тоже приближался к человеку. Особенно поразил исследователей объем мозга. Он равнялся 670680 кубическим сантиметрам, то есть был больше, чем у австралопитека. А ведь мозг как никакой другой орган характеризует положение существ на эволюционной лестнице. Удивил ученых и возраст находки около 2 миллионов лет!На территории Африки теперь известно уже около 100 местонахождений древнейших предков человека. Наиболее древнее из них лежит юго-западнее озера Рудольф (Кения). Ученые относят его к эпохе, отстоящей от нас почти на 5 500 000 лет.В слоях почвы, где находились останки презинджантропа, были найдены также грубые орудия труда из расколотой речной гальки, заостренной при помощи нескольких сколов. Установлено, что они, несомненно, использовались презинджантропом. А труд, как уже говорилось, главный показатель очеловечивания.В науке много раз обсуждался вопрос о времени возникновения труда у предков современного человека. Некоторые ученые не считали труд отличительной чертой человеческого существа. Они полагали, что коренное отличие человека от животных определяется более высоким уровнем психического развития. Никто, конечно, не отрицает такого отличительного признака, но при этом не следует забывать, что высокий уровень человеческого сознания прямой результат развития трудовой деятельности.В то же время часть ученых признает иногда орудиями труда использовавшиеся простые камни и кости, никогда не подвергавшиеся даже самой примитивной обработке. Но ведь главное тут, как уже говорилось, не в употреблении, а в изготовлении орудий труда. Накопленные археологами материалы позволяют сегодня уже достаточно четко определять признаки орудий труда, созданных человеком, и более правильно решать вопрос о времени появления труда. Пока оно датируется эпохой, когда жил презинджантроп.Еще Геккель в своей книге „Естественная история и сотворение мира" (1868) указывал на то, что в ходе эволюции между обезьяной и человеком должно было существовать какое-то переходное звено, обладавшее как человеческими, так и обезьяньими признаками. Это недостающее звено он назвал питекантропом (обезьяночеловеком). В конце прошлого века многие ученые стали искать это “недостающее звено". Голландский антрополог Э. Дюбуа в 1891 году нашел на острове Ява на глубине 15 метров коренной зуб и черепную крышку, а спустя год еще два фрагмента скелета человекоподобного существа. В 1894 году Дюбуа опубликовал описание своего открытия, которое назвал питекантропом с острова Ява". Через несколько десятков лет (с 1936 по 1939 год) на той же Яве были обнаружены еще несколько останков обезьяночеловека, а рядом с ними грубые каменные орудия, в том числе одно, напоминающее ручное рубило. Установлено, что питекантроп был значительно крупнее австралопитека: его рост не менее 170 сантиметров, объем мозга 850900 кубических сантиметров. Вспомним для сравнения, что объем мозга современных человекообразных обезьян равен 600, а человека примерно 14001600 кубическим сантиметрам. Таким образом, питекантропа можно считать переходным звеном от обезьяны к человеку. Он жил на Земле 500800 тысяч лет тому назад.Прекрасным подтверждением этого вывода оказалась находка в 20х годах в Китае останков древнего человека, названного синантропом (китайский человек). Он жил приблизительно в то же время, что и питекантроп, а объем его мозга был несколько больше.Рядом с останками синантропов обнаружены разнообразные орудия труда, изготовленные из кварца, кварцита, кремниевой гальки, из оленьего рога и трубчатых костей. В пищу синантропы употребляли мясо около 70 видов млекопитающих. Раскопки свидетельствуют, что мясо синантропы уже поджаривали на огне. Некоторые черепа животных оказались обработанными и напоминают сосуды.Огромным достижением синантропов явилось систематическое использование огня для обогрева и приготовления пищи. Это доказывается тем, что в одной из пещер слой золы от кострища достигал толщины 7 метров. Добывать огонь, вероятнее всего, эти предки человека еще не умели. Костер в пещере горел, по-видимому, беспрерывно, а для разжигания его синантропы могли использовать, например, огонь лесных пожаров, возникших от удара молнии. Речи синантропы, как и питекантропы, еще не знали. Они, наверное, способны были издавать различные звуки, а может быть, могли различать и звуковые интонации. Уровень организации их общества был стадный. Это, по существу, самая древняя форма общественной жизни.Изготовление орудий труда, пользование огнем и совместная охота, конечно, сыграли главенствующую роль в организации этих древнейших людей (объединительное название их архантропы) в общество еще очень примитивное, но уже спаянное родством и жизненной необходимостью быть вместе.Следующий важнейший этап в эволюции человека неандерталец, названный так по месту первой находки останков этого предка человека. История находки его такова.В Западной Германии, близ впадения в Рейн реки Дюссель, есть долина Неандерталь. В 1856 году здесь начали разрабатывать известковый карьер и при этом наткнулись на пещеру. В ней рабочие обнаружили 14 частей скелета человека, но не обратили на них внимания и выбросили все кости в долину. Местный учитель И. К. Фульрот собрал их и в следующем году выступил на съезде естествоиспытателей и врачей в Бонне с предположением о том, что это останки вымершего типа человека. Его сообщение произвело тогда потрясающее впечатление, так как по самой своей сути ставило под сомнение религиозную догму о божественном творении человека, то есть о том, что бог создал его по своему образу и подобию и с тех пор вид людей не мог измениться. Смущение ученых было тем более глубоким, что в то время они еще не знали о дарвиновском учении.Шли годы. Учение великого английского естествоиспытателя получало признание все большего числа ученых. К этому времени подоспели и новые находки останков человекоподобных. В 1887 году в Бельгии, в пещере БекоРош Пюид, были найдены останки двух скелетов, схожих с человеческим, а вместе с ними много кремниевых орудий и костей мамонта, пещерного медведя, шерстистого носорога и других животных. Кости скелетов, особенно черепа, были очень похожи на кости человека из Неандерталя. Так ученым ^тало ясно, что неандерталец и в самом деле один из древних предков современного человека.К настоящему времени останки неандертальцев найдены во многих частях Старого Света в Германии, Бельгии, Франции, Хорватии, Англии, Чехословакии, Испании, СССР, а также в Африке, на Яве и в других местах.Изучение останков неандертальцев и их орудий труда показывает, что по уровню организации они стояли значительно ниже современных людей. Самые заметные различия обнаруживаются в строении и форме черепа. У неандертальца лоб, например, был очень покатый, надбровные дуги сильно развиты, черепная коробка низкая, подбородок лишен выступа. Зато отличие от питекантропа разительное. Мозг неандертальца по объему уже почти приближался к мозгу человека разумного. Левая лобная доля у него несколько больше правой. Отмеченная деталь говорит о том, что неандерталец был правшой, как и современный человек. Вообще лобные доли черепа у неандертальцев гораздо больше, чем у других предшественников человека. Это как раз и свидетельствует об относительно более развитых умственных способностях, правда в сравнении с человеком еще довольно ограниченных. Но они компенсировались хорошо развитыми анализаторами органов чувств, о чем свидетельствует расширение затылочных, теменных и височных долей.Неандерталец еще не обладал развитой членораздельной речью. Вероятнее всего, он только-только начинал осваивать ее. Неандертальцы создали свою своеобразную культуру так называемую мустьерскую культуру каменного века (15040 тысяч лет назад).В период великого оледенения неандертальцы, чтобы спастись от холода и свирепых хищников, часто селились в пещерах. Они умели изготовлять уже довольно хорошо отделанные орудия из кремня, кости и дерева, охотились на таких больших и сильных животных, как мамонты и пещерные медведи. Вполне возможно, что они уже научились добывать огонь и использовать шкуры животных как одежду.Чрезвычайно заинтересовали ученых неандертальские погребения. Многие исследователи сегодня считают, что такие погребения доказательство существованияу неандертальцев каких-то нравственных норм. Один из крупнейших советских археологов, академик А. П. Окладников, открывший на территории нашей страны первое неандертальское погребение, пишет: „В середине и в конце мустьерского времени, к которым относятся перечисленные погребения (неандертальские), впервые появляется определенное и совершенно новое отношение к умершим, выраженное в намеренных и уже достаточно сложных по характеру действиях в захоронениях трупов. В основе этого отношения лежала, несомненно, забота о сочлене своего коллектива, вытекающая из всего жизненного строя первобытной общины, из всех неписаных законов и норм поведения того времени. Это было бесспорное выражение того чувства неразрывной кровной связи между сородичами, которое проходит красной нитью через всю первобытную эпоху истории человечества". К концу мустьерского времени неандертальцы уже расселились на обширных пространствах Европы, Азии и Африки. В некоторых же местах они достигли даже Полярного круга. Очевидно, суровый климат не пугал их. Они могли к нему приспосабливаться.Неандертальцы и созданная ими культура явились, по сути, предшественниками современного человека и культур верхнего палеолита (позднего каменного века). Об этом свидетельствуют находки ученых, которые показывают, что многие черты мустьерских памятников характерны и для более поздних культур. Постепенно неандертальцы исчезали с лица Земли. Некоторые из их видов вымерли. Процесс становления современного человека, очевидно, в большой степени связан с приспособлением к разнообразным изменениям климата, происходившим в те эпохи на Земле. Выживали те виды существ, которые могли лучше приспособиться к жизни в суровых условиях планеты, те, кто был в состоянии формировать подлинное человеческое общество, преодолевать дикие инстинкты, совершенствовать орудия труда. Вероятно, к этому времени в объединениях человекоподобных существ естественный отбор уже терял свое былое значение.Прошли многие и многие десятки тысяч лет, прежде чем на нашей планете окончательно воцарился современный человек. Homo sapiens (человек разумный), как его именуют в науке. По месту первой находки его назвали кроманьонцем (1868 год, пещера КроМаньон во Франции).Это были крупные люди высотой до 180 сантиметров, с объемом черепа до 1600 кубических сантиметров. Жили они примерно от 50 до 15 тысяч лет назад. По своему внешнему виду кроманьонцы заметно отличались от неандертальцев: лоб прямой, вместо надглазного валика надбровные дуги, явно выраженный подбородочный выступ. Это уже облик современного человека. Они изготовляли разные орудия из камня, кости и рога, в том числе составные орудия, что свидетельствует о значительном прогрессе в этой области.Находок кроманьонцев и других родственных им типов людей в настоящее время известно много. Есть они и на территории нашей страны. Эти люди уже составляли человеческое общество со сложными общественными взаимоотношениями. Дальнейшая их история уже особая тема, выходящая за рамки ших бесед.При обсуждении проблемы происхождения человека и его древнейших предков возникает еще один вопрос: как произошли человеческие расы? Как известно, христианская религия на это отвечала довольно просто: расы и народы, мол, произошли от трех сыновей Ноя Сима, Хама и Иафета. „От них распространились народы по земле после потопа" (Быт. 10:32).Многочисленные исследования различных ученых показали, что этапы, которые человек проходил в своем развитии, едины для любой части нашей планеты. Расовые различия в том виде, в каком мы их видим сейчас, образовались сравнительно поздно. Они появились лишь у человека современного вида, в эпоху верхнего палеолита.Многие считают, что появление различных рас объясняется своеобразными условиями природной среды. Как уже говорилось, условия обитания формировавшегося человека весьма значительно отличались друг от друга. Позднее, когда сложился тип современного человека, умевшего лучше защищаться от неблагоприятных воздействий внешней среды, влияние ее на человека ослабло, однако все еще продолжало сохраняться. Ведь и сейчас, например, если человек постоянно живет на юге, пигментация кожи у него отличается от пигментации кожи людей той же расы, но живущих на севере.На заре человеческой истории удаленные друг от друга на большие расстояния отдельные группы людей почти не общались друг с другом. В результате у изолированных групп людей признаки, сложившиеся под влиянием природных условий (например, более темный цвет кожи под жарким солнцем), начинали закрепляться в потомстве, становились все более явными в данной группе. Процесс этот был очень длительным.В ходе дальнейшей человеческой истории люди с различными внешними признаками много раз смешивались. И сегодня можно уверенно сказать: на Земле нет чистых рас. Деление на них крайне условно, человечество едино по своему происхождению и по своей природе.Поэтому совершенно лишены каких-либо научных оснований разного рода расистские измышления о якобы существующем биологическом неравенстве различных народов. Сравнительно-анатомические исследования убедительно доказали полное сходство всех человеческих рас. Мало общего с наукой имеют и рассуждения о происхождении различных рас от разных групп древних людей или различных подвидов более ранних предшественников человека: ведь останки древних людей были обнаружены не только в Азии, но и в Африке, а неандертальцы почти на всех материках Старого Света.Биологическое и социальное в историческом развитии человеческого рода Первый вопрос, который в связи с этим возникает: продолжается ли биологическая эволюция человека после возникновения 30—40 тыс. лет назад Homo sapiens? Другими словами, как культурная организация жизни людей, включая способы производства, развитие трудовой деятельности, образ жизни и т. д., могла влиять на биологическую эволюцию человека и влияла ли она на нее вообще?Строго говоря, эволюция человека происходит на всем протяжении его существования. Но она относится к социальной стороне его жизни. Что же касается биологической эволюции человека, то с тех пор как он выделился из животного мира, она перестала иметь по крайней мере решающее значение. Даже люди с ослабленным здоровьем благодаря успехам медицины могут принимать активное участие в жизни общества. Сила естественного отбора в социальном мире все более ослабевает, так как социальные институты, здравоохранение постоянно сглаживают влияние индивидуальной биологической изменчивости. Так, снижение в Европе уровня смертности от туберкулеза с 4000 человек на миллион в 1840 г. до 13 человек на тот же миллион в настоящее время свидетельствует о том, что естественный отбор по сопротивляемости туберкулезу по существу прекратился. Это же можно сказать и о многих других болезнях.Сегодня наблюдается очень медленный темп генетических изменений, производимых отбором, и вместе с тем большое генетическое сходство между различными человеческими группами. С другой стороны, имеется огромное разнообразие человеческих культур и образов жизней, очень быстрый рост социальных изменений, свидетельствующих о происходящей культурной эволюции человечества. Поэтому можно с уверенностью говорить о ведущей роли культуры в эволюции Homo sapiens. Политические, экономические и социальные изменения во многих странах, связанные с улучшением жизни людей, прямо влияют на состояние их здоровья и, следовательно, на уменьшение зависимости человека от естественного отбора. У человека роль естественного отбора заключается в сохранении генофонда, в сдерживании мутаций, отрицательно влияющих на его здоровье.Но меняется ли вместе с социальным обликом человека и его биологическая природа, его физический облик, его умственные способности? Становятся ли новые поколения людей более развитыми в физическом и умственном плане?Прежде всего о физическом здоровье. Вполне очевидно, что его состояние за историю Homo sapiens существенно улучшилось. Комплексным показателем здесь может служить увеличение средней продолжительности жизни населения. Под влиянием социальных условий она возросла с 20—22 лет в древности до 30 лет в XVIII в. К началу XX в. в странах Западной Европы она составляла примерно 56 лет. Сегодня в этих странах средняя продолжительность достигла 75—78 лет, т.е. приближается, по данным современной науки, к средней «нормальной» продолжительности жизни Homo sapiens — 80—90 лет.Теперь обратимся к вопросу о развитии умственных способностей. Один из создателей евгеники (теории о наследственном здоровье человека и путях его улучшения) английский психолог и антрополог Фрэнсис Гальтон (1822—1911) был убежден в том, что интеллект современного человека снижается. По его мнению, представители низших классов обладают более низким коэффициентом интеллектуальности — IQ. В то же самое время именно эти слои имеют и больше детей. Причем статистические данные, говорившие о более высоком репродуктивном уровне людей с более низким IQ, были широко распространены до сравнительно недавнего времени. На основании этих предпосылок Гальтоном и некоторыми другими учеными делался вывод о том, что человеческий вид будет все более наполняться «худшими породами» людей, все более уменьшать свой IQ. Однако в начале 60-х гг. была показана ошибочность того, что существует прямая зависимость между социальным положением, количеством детей и IQ.К тому же у человека нет прямой зависимости между величиной мозга и индивидуальной одаренностью. Так, по имеющимся данным, довольно маленький мозг среди одаренных людей имел известный французский писатель Анатоль Франс. У него размер мозга не превышал 1000 куб. см. Большой объем мозга имел И.С. Тургенев — 2012 куб. см. Но вряд ли кто возьмется сравнивать их способности и тем более ставить последние в зависимость от объема мозга.Современные биологи и антропологи полагают, что процесс биологической эволюции человека как вида, т.е. процесс видообразования человека, прекратился со времени образования Homo sapiens. Прежде всего об этом свидетельствует тот факт, что в течение этого периода мозг человека не изменился, морфологическое изменение его завершилось. Противоположная точка зрения не имеет достаточного основания.С возникновением человека и общества генетическая информация утрачивает свое главенствующее значение в жизнедеятельности вида человек. Она заменяется социальной информацией, развитие которой «определяется уже не столько естественным отбором наиболее умелых и одаренных, сколько социальными факторами, которым подчиняется и общебиологический процесс».При ответе на вопрос, какое будущее ожидает человечество с точки зрения развития вида, иногда высказывается мнение, что все виды животных и растений постепенно вымрут вследствие деградации генома (генетической программы развития). По мнению большинства ученых, самая главная опасность состоит не в старении вида, а в опасности все большего загрязнения биосферы различного рода отходами, повышении уровня радиации в связи с авариями, в увеличении мутационной опасности химических загрязнителей и т. п. Достаточно отметить, что в России в конце прошлого столетия уровень рождаемости детей с генетическими отклонениями достиг 17 процентов. Все более увеличивается число людей, страдающих слабоумием (олигофренией), имеющим генетическое происхождение. Все это свидетельствует о том, что главная опасность и угроза дальнейшему существованию человека — в нашей собственной культуре, а не в мифическом старении вида или каких-либо других биологических факторах.Биологическое и социальное в онтогенезе человека.Рассмотрим соотношение биологических и социальных факторов в индивидуальном развитии человека, или в его онтогенезе. В современной литературе по вопросу о роли социальных и биологических факторов в индивидуальном развитии человека существуют два различных подхода к решению проблемы. Одни авторы утверждают, что развитие человека целиком обусловлено генами, абсолютизируя, таким образом, биологический фактор. Это направление называется панбиологизм. Вторая точка зрения состоит в том, что все люди рождаются с одинаковыми генетическими задатками, а главную роль в развитии их способностей играют воспитание и образование. Эта концепция получила название пансоциологизм.Рассматривая данную проблему, следует иметь в виду, что в ин¬дивидуальном развитии человека различаются два периода — эмбриональный и постэмбриональный. Первый охватывает период внутриутробного развития человеческого эмбриона (зародыша).Уже на самой ранней стадии развития эмбриона начинается реализация генетической программы, полученной от родителей и закрепленной в хромосомах ДНК. Каждый человек является носи¬телем специфического индивидуального набора генов, вследствие чего он генетически уникален. Свойства человека, как и других живых существ, во многом детерминированы генотипом, а их передача от поколения к поколению происходит на основе законов наследственности. Индивид наследует от родителей такие свойства, как телосложение, рост, массу, особенности скелета, цвет кожи, глаз и волос, химическую активность клеток. Многие также говорят о наследовании способности к вычислению в уме, склонности к тем или иным наукам и т. д.На сегодняшний день господствующей точкой зрения можно считать ту, которая утверждает, что наследуются не сами способности как таковые, а лишь их задатки, которые в большей или меньшей степени могут проявляться в условиях среды. Генетическим материалом у человека, как и у других млекопитающих, является ДНК, которая находится в хромосомах. Хромосомы каждой клетки человека несут в себе несколько миллионов генов. При этом генетические возможности, задатки реализуются только при том условии, что ребенок с раннего детства находится в общении с людьми, в соответствующей социальной среде. Если, например, у человека нет возможности заниматься музыкой, то его врожденные музыкальные задатки так и останутся неразвитыми.Генетический потенциал человека ограничен во времени, причем достаточно жестко. Если пропустить срок ранней социализации, этот потенциал не успевает реализоваться и угасает. Ярким примером этого могут служить многочисленные случаи, когда младенцы, силой обстоятельств попавшие в джунгли, проводили среди зверей несколько лет. После возвращения их в человеческое сообщество они не могли уже в полной мере наверстать упущенное, не смогли овладеть речью, приобрести достаточно сложные навыки человеческой деятельности, у них плохо развивались человеческие психические функции. Это свидетельствует о том, что характерные черты человеческого поведения и деятельности приобретаются только через социальное наследование, через передачу социальной программы в процессе воспитания и обучения.Для понимания роли наследственности и среды в онтогенезе человека важное значение имеют такие понятия, как генотип и фенотип. Генотип — это наследственная основа организма, совокупность генов, локализованных в его хромосомах; это та генетическая конституция, которую организм получает от своих родителей. Фенотип — это совокупность всех свойств и признаков организма, сформировавшихся в процессе его индивидуального развития.Фенотип определяется взаимодействием организма с условиями среды, в которых протекает его развитие. В отличие от генотипа фенотип изменяется на протяжении жизни организма. Таким образом, фенотип зависит от генотипа и среды. Одинаковые генотипы (у однояйцовых близнецов), которые оказываются в различных средах, могут давать различные фенотипы. С учетом сказанного можно выделить следующие факторы, воздействующие на человека:- биологические задатки, кодируемые в генах;- среда (социальная и природная);- деятельность индивида;- ум (сознание, мышление).Взаимодействие наследственности и среды в развитии человека имеет место в течение всей его жизни. Но особую важность оно приобретает в периоды формирования организма: эмбрионального, грудного, детского, подросткового и юношеского. Именно в эти периоды наблюдается интенсивный процесс развития организма и формирования личности.Наследственность определяет то, каким может стать организм, но формируется он под одновременным влиянием обоих факторов как наследственности, так и среды. Сегодня становится все более общепризнанным, что адаптация человека осуществляется под влиянием двух программ наследования — биологической и социальной. Все признаки и свойства любого индивида являются, таким образом, результатом взаимодействия его генотипа и среды. Поэтому каждый человек является и частью природы, и продуктом общественного развития.С такой позицией сегодня согласно большинство ученых. Разногласие возникает тогда, когда речь заходит о роли наследственности и среды в детерминации умственных способностей человека.Следует отметить, что точное определение самих умственных способностей представляет собой довольно трудную задачу. Дело в том, что, прежде всего, сами интеллектуальные способности весьма разнообразны и своеобразны. Человек может быть гениальным шахматистом и плохим артистом (поэтом, математиком и т. д.), и (наоборот. Кроме того, сама процедура применения тестов на определение IQ имеет недостатки, которые отмечают многие ученые. В частности, при определении IQ многое зависит от социальной среды, уровня и характера воспитания и образования испытуемых, их организованности, внимательности, собранности и даже темперамента. Более того, результаты тестирования зависят не только от испытуемых, но и от тестирующих: какие вопросы задаются, с какой целью, из какой области знания или деятельности и т. д. Так, если детям с улицы задавать вопросы, как вести себя среди почтенной публики, а детей аристократов спрашивать, как вести кулачный бой, то, по всей вероятности, IQ тех и других будет невелик и достаточно одинаков.
89. Популяционная структура человечества. Демы. Изоляты. Люди как объект действия эволюционных Факторов.В антропогенетике популяцией называют группу людей, занимающих общую территорию и свободно вступающих в брак. Изоляционные барьеры, препятствующие заключению брачных союзов, нередко несет выраженный социальный характер (например, вероисповедание). Размер, уровень рождаемости и смертности, возрастной состав, экономическое состояние, уклад жизни являются демографическими показателями популяций людей. Генетически они характеризуются генофондами. Большое значение в определении структуры браков имеет размер группы. Популяции из 1500-4000 человек называют демами, популяции численностью до1500 человек — изолятами. Для демов и изолятов типичен относительно низкий естественный прирост населения — соответственно порядка 20% и не более 25% за поколение. В силу частоты внутригрупповых браков члены изолятов, просуществовавших 4 поколения и более, являются не менее чем троюродными братьями и сестрами. В настоящее время усилились миграции населения в связи с ростом численности людей, совершенствованием средств транспорта, неравномерным развитием экономики. Популяционные волны — периодические колебания численности людей на обширных или ограниченных территориях, изменение плотности населения (приросты совпадают с важнейшими достижениями человечества, упадок — чума, болезни, войны). Природа изоляционных барьеров между популяциями людей разнообразна. Специфическими для человеческого общества являются формы изоляции, зависящие от разнообразия культур, экономических укладов, религиозных и морально-этических установок. Фактор изоляции оказывал влияние на генофонды популяций людей.
90. Понятие о расах и видовое единство человечества. Современная классификация и распространения человеческих рас.Ра́са — система человеческих популяций, характеризующихся сходством по комплексу определённых наследственных биологических признаков, имеющим внешнее фенотипическое проявление и сформировавшимся в определенном географическом регионе[1]. Черты, характеризующие разные расы, зачастую появляются как результат адаптации к различным условиям среды, происходившей в течение многих поколений.Критерием отличия расы от вида является отсутствие существенных препятствий для создания плодовитого потомства, что приводит к образованию множества переходных форм в области смешения рас.Содержание понятия «раса» более или менее однозначно воспринимается разными исследователями[источник не указан 428 дней], однако в деталях имеются существенные расхождения; разные школы антропологов выделяли от четырёх до семи основных рас и десятки малых антропологических типов. В истории науки было несколько попыток доказать, что все расы людей являются абсолютно различными видами и даже родами. Все же существует мнение, причем его придерживаются большая часть исследователей, что все человечество происходит от единого вида.Все типы людей могут, вступая в брак, рождать способное к размножению потомство, соответственно, они происходят от одного общего ствола. Это главное умозаключение унитаристской школы. Учение о видовом единстве человечества носит название моногенетизмаВсе расы, если они перемешиваются, производят плодовитое потомство. Отсюда понятны огромные цифры рождаемости среди людей Южной Америки, Южной Азии и других областей, где проживают много рас. К примеру, в Бразилии число «чистых» индейцев, которые до сих пор проживают в лесах достигает едва 2% от всего количества населения, а вот количество метисов куда больше – целых 33%.Во всех районах соприкосновения различных расовых типов неизбежен процесс смешения. Это стихийный процесс смешения рас не в силах остановить на какие-либо социальные преграды, ни любого вида террористические меры, которые призваны сохранить «чистоту» расы.Не смотря на то, что некоторые расы по внешности разительно отличаются друг от друга, например: пигмеи Африки, жители Северной страницы и Южные китайцы, все они взаимосвязаны между собой несколькими промежуточными типами.
91. Биосфера как естественно-историческая система. Вклад русских и советских ученых в развитие учения о биосфере (В.В.Докучаев, В.И. Вернадский).Термин «биосфера» введен австрийским геологом Зюссом в 1875 году для обозначения особой оболочки Земли, образованной совокупностью живых организмов, что соответствует биологической концепции биосферы. Вернадский развил это направление и разработал учение о биосфере как глобальной системы нашей планеты, в которой основной ход геохимических и энергетических превращений определяется живым веществом. Он распространил понятие биосферы не только на сами организмы, но и на среду их обитания, чем придал концепции биосферы биогеохимический смысл. Большинство явлений, меняющих в масштабе геологического времени облик Земли, рассматривали ранее как чисто физические, химические или физико-химические. С именем Вернадского связано также формирование социально- экономической концепции биосферы, отражающей ее превращение на определенном этапе эволюции в ноосферу вследствие деятельности человека, которая приобретает роль самостоятельной геологической силы. Учитывая системный принцип организации биосферы, а также то, что в основе ее функционирования лежат круговороты веществ и энергии, современной наукой сформулированы биохимическая, термодинамическая, биогеоценотическая, кибернетическая концепции биосферы. Биосферой называют оболочку Земли, которая населена и активно преобразуется живыми существами. Биосфера — это такая оболочка, в которой существует, и существовала в прошлом жизнь и которая подвергалась и подвергается воздействию живых организмов. Она включает: живое вещество, образованное совокупностью организмов; биогенное вещество, которое создается и перерабатывается в процессе жизнедеятельности организмов; косное вещество, которое образуется без участия живых организмов; биокосное вещество, представляющее собой совместный результат жизнедеятельности организмов и абиогенных процессов. 92. Функции биосферы в развитии природы Земли и поддержания в ней динамического равновесия.Биосфера, по Вернадскому, – земная оболочка, область существования живого вещества. Она включает в себя не только живые организмы, но и изменённую ими среду обитания (кислород в атмосфере, горные породы органического происхождения и т.п.).Биосфера является одной из геологических оболочек Земли или геосфер. На Земле также различают литосферу – твёрдую наружную оболочку Земли, состоящую из осадочных пород и расположенных под ними гранитов и базальтов, гидросферу, включающую в себя все океаны, моря, озёра и реки, и атмосферу – газовую оболочку Земли. В состав биосферы входят верхние слои литосферы, нижний слой атмосферы (тропосфера) и вся гидросфера, связанные между собой сложными круговоротами веществ и энергии. Нижний предел жизни на Земле (до глубины 3 км) ограничен высокой температурой земных недр, верхний предел (20 км) – жёстким излучением ультрафиолетовых лучей (всё, что находится на высоте ниже 20 км, защищено от губительного излучения двадцатикилометровым озоновым слоем). Тем не менее, на границах биосферы можно найти, в основном, лишь микроорганизмы (обычно в виде спор); наибольшая же концентрация биомассы наблюдается у поверхности суши и океана, в местах соприкосновения оболочек. Организмы, составляющие биосферу, обладают поразительной способностью к размножению и распространению по планете.Биосфера играет важную роль в распределении энергетических потоков на Земле. В год до Земли доходит около 1024 Дж солнечной энергии; 42 % из неё отражается обратно в космос, а остальное поглощается. Другим источником энергии является тепло земных недр. 20 % энергии переизлучается в мировое пространство в виде тепла, 10 % расходуется на испарение воды с поверхности Мирового океана. Зелёные растения преобразуют в процессе фотосинтеза около 1022 Дж в год, поглощают 1,7 ∙ 108 т CO2, выделяют около 11,5 ∙ 107 т кислорода и испаряют 1,6 ∙ 1013 т воды. Исчезновение растений привело бы к катастрофическому накоплению углекислоты в атмосфере, и через сотню лет жизнь на Земле в её нынешних проявлениях погибла бы. Наряду с фотосинтезом в биосфере происходит почти такое же по масштабам окисление органических веществ в процессах дыхания и разложения.В организмах содержатся все известные сегодня химические элементы. Если некоторые из них (водород, кислород, углерод, азот, фосфор и другие) являются основой жизни, то другие (рубидий, платина, уран) имеются в организмах в очень малых количествах. Организмы участвуют в миграции химических элементов как прямо (выделение кислорода в атмосферу, окисление и восстановление различных веществ в почвах и гидросфере), так и косвенно (восстановление сульфатов, окисление соединений железа, марганца и других элементов). Биогенная миграция атомов вызвана тремя основными процессами: обменом веществ, ростом и размножением организмов. Огромную роль в биогеохимической активности играет человек, извлекая ежедневно в ходе добычи полезных ископаемых миллиарды тонн горной породы. Влияние человека на глобальные геохимические процессы с каждым годом только растёт.
93. Биосфера как естественно-историческая система. Современные концепции биосферы: биохимическая; биогеоценологическая, термодинамическая, геофизическая, социально-экологическая.Биосфера - оболочка Земли, состав, структура и энергетика которой определяются совокупной деятельностью живых организмов. Термин «биосфера» ввёл австралийский геолог Э. Зюсс в 1875 году, который понимал её как тонкую плёнку жизни на земной поверхности, в значительной мере определяющую «лик Земли». Центральное звено в концепции В.И. Вернадского о биосфере - представление о живом веществе: «Живые организмы, - писал он, - являются функцией биосферы и теснейшим образом материально и энергетически с ней связаны, являются огромной геологической силой, её определяющей». С именем В.И. Вернадского связано также формирование социально-экономической концепции биосферы, суть которой - в превращении биосферы в ноосферу вследствие деятельности новой геологической силы. Исходя из того, что биосфера организована по системному принципу, а также того, что в основе её функционирования лежат круговороты веществ и энергии, сформулированы несколько современных концепций биосферы: биохимическая, биогеоценотическая, кибернетическая, термодинамическая. Биогеоценотическая концепция, например, рассматривает биосферу как сложноорганизованную систему биогеоценозов (динамическую целостную систему организмов разных видов, тесно связанных со средой их обитания), биохимическая концепция - как систему моделей геохимических циклов биогенных элементов. 94. Человек и биосфера. Ноосфера - высший этап эволюции биосферы.Человек, как было отмечено выше, является частью биосферы. Все необходимое для жизни — воду, пищу, значительную часть энергии и строительного материала своих органов — он получает из биосферы. В биосферу человек сбрасывает отходы своей жизнедеятельности. Долгое время природа перерабатывала эти отходы и сохраняла свое равновесие. Однако в последнее столетие вмешательство человека в природные процессы стало не только слишком сильным, но чрезмерным по своим масштабам. По представлениям биологов, в природе действует «правило 10%», согласно которому она в экстремальных ситуациях способна выдержать десятикратную нагрузку по сравнению с обычной. Человек своим воздействием на природу вплотную подошел к этому рубежу, и поэтому сегодня среди прочих глобальных проблем человечества возникла щобальная экологическая проблема сохранения жизни на Земле.Ноосфе́ра (греч. νόος — «разум» и σφαῖρα — «шар») — сфера разума; сфера взаимодействия общества и природы, в границах которой разумная человеческая деятельность становится определяющим фактором развития (эта сфера обозначается также терминами «антропосфера», «биосфера», «биотехносфера»)[1].Ноосфера — предположительно новая, высшая стадия эволюции биосферы, становление которой связано с развитием общества, оказывающего глубокое воздействие на природные процессы. Согласно В. И. Вернадскому, «в биосфере существует великая геологическая, быть может, космическая сила, планетное действие которой обычно не принимается во внимание в представлениях о космосе… Эта сила есть разум человека, устремленная и организованная воля его как существа общественного»[1].
95. Определение науки экологии. Среда как экологическое понятие.Факторы среды. Экосистема, биогеоценоз, антропобиоценоз.Экология — познание экономики природы, одновременное исследование всех взаимоотношений живого с органическими и неорганическими компонентами среды… Одним словом, экология — это наука, изучающая все сложные взаимосвязи в природе, рассматриваемые Дарвином как условия борьбы за существование. (Это определение Э. Геккеля написано в те времена, когда экология была ещё исключительно биологической наукой. Нынешнее понимание экологии шире.)Экология — биологическая наука, которая исследует структуру и функционирование систем надорганизменного уровня (популяции, сообщества, экосистемы) в пространстве и времени в естественных и измененных человеком условиях. Это определение дано на 5-м Международном экологическом конгрессе (1990) с целью противодействия размыванию понятия экологии, наблюдаемому в настоящее время.Среда обитания — это часть природы, окружающая живые организмы и оказывающая на них прямое или косвенное воздействие. Из среды организмы получают все необходимое для жизни и в неё же выделяют продукты обмена веществ. Среда каждого организма слагается из множества элементов неорганической и органической природы и элементов, привносимых человеком и его производственной деятельностью. При этом одни элементы могут быть частично или полностью безразличны организму, другие необходимы, а третьи оказывают отрицательное воздействие. Есть четыре среды обитания Вода, Наземно-воздушная, Почва и Организм.Не тронутая человеком среда обитания многих растений и животныхОтдельные свойства и элементы среды, воздействующие на организмы, называют экологическими факторами. Все экологические факторы можно разделить на две большие группы:Абиотические факторы — это комплекс условий неорганической среды, влияющих на организм. (Свет, температура, ветер, воздух, давление, и т. д.)Биотические факторы — это совокупность влияний жизнедеятельности одних организмов на другие. (Влияние растений и животных на других членов биогеоценоза)Антропогенные (антропические) факторы — это все формы деятельности человеческого общества, изменяющие природу как среду обитания живых организмов или непосредственно влияющие на их жизнь. Выделение антропогенных факторов в отдельную группу обусловлено тем, что в настоящее время судьба растительного покрова Земли и всех ныне существующих видов организмов практически находится в руках человеческого общества.Возможно также выделить следующие компоненты среды обитания: естественные тела среды обитания, гидросреду, воздушное пространство среды, атропогенные тела, поле излучений и тяготения среды.В ООН создана специальная организация — Программа ООН по окружающей среде (ЮНЕП). В целях привлечения внимания к проблемам охраны окружающей среды ООН установила Всемирный день окружающей среды.Экосистема или экологическая система (от греч. óikos — жилище, местопребывание и система), природный комплекс (биокосная система), образованный живыми организмами (биоценоз) и средой их обитания (косной, например атмосфера, или биокосной — почва, водоём и т. п.), связанными между собой обменом веществ и энергии. Одно из основных понятий экологии, приложимое к объектам разной сложности и размеров. Примеры Экосистем — пруд с обитающими в нём растениями, рыбами, беспозвоночными животными, микроорганизмами, донными отложениями, с характерными для него изменениями температуры, количества растворённого в воде кислорода, состава воды и т. п., с определённой биологической продуктивностью; лес с лесной подстилкой, почвой, микроорганизмами, с населяющими его птицами, травоядными и хищными млекопитающими, с характерным для него распределением температуры и влажности воздуха, света, почвенных вод и др. факторов среды, с присущим ему обменом веществ и энергии. Гниющий пень в лесу, с живущими на нём и в нём организмами и условиями обитания, тоже можно рассматривать как ЭкосистемуБиогеоценоз (от био, гео и греч. koinós — общий), сообщество живых существ, тесно связанных между собой и окружающей средойкруговоротом веществ и потоком энергииСвойства биогеоценозаестественная, исторически сложившаяся системасистема, способная к саморегуляции и поддержанию своего состава на определенном постоянном уровнехарактерен круговорот веществоткрытая система для поступления и выхода энергии, основной источник которой — СолнцеАНТРОПОБИОЦЕНОЗ (от антропо... и биоценоз) , биоценоз, находящийся под воздействием хозяйственной деятельности человека.
96. Человек как творческий экологический фактор. Роль медицинских работников в охране окружающей среды.Известно, что к 2000 году население Земли достигло шести миллиардовчеловек. Для их жизнеобеспечения необходимо ежегодно добывать миллиардытонн сырьевых ресурсов, вырабатывать гигантское количество энергии,получать громадные объемы сельскохозяйственной продукции. И любой видпроизводственной деятельности человека – будь то промышленность,энергетика, транспорт или выращивание урожая – обязательно влечет за собойзагрязнение природной среды. Научные прогнозы говорят о том, что ксередине ХХI столетия численность землян достигнет, как минимум, 10-тимиллиардов человек. Хочется надеяться, что качество их жизни будет выше,чем сегодня у жителей слаборазвитых стран: будет потребляться больше пищи,энергии, товаров на душу населения.Значит, если не принять действенных своевременных мер, антропогеннаятоксикация планеты через полвека может достичь уровня, с которым природа несможет справиться, и экологический кризис превратится в глобальнуюкатастрофу. Слово «катастрофа» ассоциируется с понятием неожиданности,внезапности. Природные экологические катастрофы (землетрясения, смерчи,тайфуны, лавины) дополняются антропогенными (Чернобыль, заводские взрывы,аварийные выбросы ядовитых веществ). Существуют, однако, и медленноразвивающиеся экологические катастрофы, приобретающие со временемглобальный характер. Медленное развитие придает им особую опасность,пожалуй, не меньшую, чем внезапность. Общественное сознание не поражают, ачеловечество не мобилизуют на немедленные действия факты постепенного ростанекоторых заболеваний, повышение кислотности дождей, снижение урожайностипочв, уменьшения фертильности мужской спермы и другие подобные явления,признанные экологическим следствием производственной деятельностичеловечества. Одной из таких медленно развивающихся катастроф являетсятоксикация нашей планеты.Некоторые исследователи характеризуют современную нам эпоху как этапперехода к постиндустриальной (информационной) цивилизации, подразумеваяпод этим, что уже сегодня фактически осуществляется переход к главенствупроизводства информации, знаний и гармонизации на этой основевзаимоотношений человека и природы.Еще Ф. Бэкон обращал внимание на особую роль, которую призвано игратьнаучное знание в развитии взаимоотношений общества и природы. Он считал,что знание законов природы позволит людям удовлетворить свои насущныепотребности и интересы.Эти представления были конкретизированы и развиты в получившей в ХХв. широкое распространение и научное признание концепции ноосферы. Понятиеноосферы было введено в науку в 1927 г. французским философом, математикоми антропологом Э. Леруа (1870 – 1954), предложившим назвать ноосферойоболочку Земли, включающую человеческое общество с его индустрией, языком ипрочими атрибутами разумной деятельности. Но главным творцом ноосфернойконцепции по праву считается русский естествоиспытатель и мыслитель В.И.Вернадский (1863 – 1945), развивший в своих трудах идею ноосферы как«биосферизированного общества». Он одним из первых осознал, чточеловечество стало мощной геологической и, возможно, космической силой,способной преобразовывать природу в больших масштабах. Биосфера, с еготочки зрения, постепенно преобразуется в ноосферу – сферу разума.В.И. Вернадский был убежден, что ноосферное человечество найдет путь квосстановлению и сохранению экологического равновесия на планете,разработает и осуществит на практике стратегию бескризисного развитияприроды и общества. При этом полагал, что человек вполне способен принятьна себя функции управления экологическим развитием планеты в целом.По мнению многих современных специалистов-экологов, человечеству ещепредстоит доказать его способность осуществить все то, о чем шла речь выше.Обеспокоенные существующим положением многие видные экологи, социологи,политики и др. начиная со второй половины 70-х гг. ХХ в. объединили своиусилия с целью выработки нового подхода к построению взаимоотношений междучеловеком и средой его обитания. Результатом проделанной работы сталаформулировка концепции устойчивого развития. Согласно определениюМеждународной комиссии по окружающей среде и развитию под устойчивым должнопонимать такое развитие, при котором удовлетворение потребностейсовременного человечества не ставит под угрозу благополучие последующихпоколений и их способность удовлетворять собственные насущные потребности.Важнейшей задачей в этой связи становится охрана окружающей среды, целькоторой, в конечном счете, сводится к тому, чтобы, с одной стороны,обеспечить сохранность таких качеств окружающей среды, которые не должныбыть подвергнуты изменениям, а с другой стороны – обеспечить непрерывныйурожай полезных растений, животных и других необходимых человеку ресурсовпутем сбалансированных циклов изъятия и обновления.Концепция устойчивого развития получила мощную поддержку не только состороны специалистов в области социальной экологии и экологии человека, нои правительств и руководителей государств большинства стран мира, что нашлосвое выражение в решениях Конференции ООН по окружающей среде и развитию,состоявшейся в 1992 г. в Рио-де-Жанейро. Конференция приняла решение обобразовании организации Международный Зеленый Крест, главными задачамикоторой были объявлены экологическое образование и воспитание как основаустойчивого развития и изменения системы ценностей, а также ликвидацияпоследствий «холодной войны» для окружающей среды.В 1996 г. в соответствии с рекомендациями Конференции ООН поокружающей среде и развитию 1992 г. была разработана и утверждена концепцияперехода Российской Федерации к устойчивому развитию. Концепция должнастать основой для выработки стратегии перехода России к устойчивомуразвитию в ХХI в.Охра́на приро́ды — комплекс мер по сохранению, рациональному использованию и восстановлению природных ресурсов Земли, в том числе видового разнообразия флоры и фауны, богатства недр, чистоты вод и атмосферы.Опасность неконтролируемого изменения окружающей среды и вследствие этого угроза существованию на Земле живых организмов, в том числе человека, потребовали решительных практических мер по защите и охране природы, правового регулирования использования природных ресурсов. К таким мерам относятся создание безотходных технологий, очистных сооружений, упорядочение использования пестицидов, прекращение производства ядохимикатов, способных накапливаться в организме, рекультивация земель и пр., а также создание охраняемых территорий (заповедники, национальные парки и др.), центров по разведению редких и исчезающих животных и растений (в том числе для сохранения генофонда Земли), составление мировой и национальных Красных книг.Рациональное природопользование - планомерное, научно обоснованное преобразование окружающей среды по мере совершенствования материального производства на основе комплексного использования невозобновляемых ресурсов в цикле «производство - потребление - вторичные ресурсы» при условии сохранения и воспроизводства возобновляемых природных ресурсов.В XX веке человечество в результате научно-технической революции пришло к техническому круговороту веществ, изображенному на рис. 12.Изучение процессов, протекающих в биосфере, и влияние на них хозяйственной деятельности человека показывает, что только создание экологически безотходных и малоотходных производств может предотвратить оскудение ресурсов и деградацию окружающей среды. Хозяйственная деятельность людей должна строиться по принципу природных экосистем, которые экономно расходуют вещество и энергию и в которых отходы одних организмов служат средой обитания для других, т.е. осуществляется круговорот веществ.В ноябре 1979 года в Женеве было созвано совещание по сотрудничеству в области охраны окружающей среды и была принята декларация, которая гласит: «Важнейшими условиями малоотходной и безотходной технологии и использования отходов являются охрана окружающей среды и рациональное использование ресурсов».
97. Предмет экологии человека. Биологической и социальный аспекты адаптации населения к условиям жизнедеятельности.Экология человека (или социальная экология) - это область экологии, изучающая взаимодействие человеческого общества и окружающей среды. Экология человека изучает закономерности возникновения, существования и развития антропоэкологических систем. Размеры таких систем различны в зависимости от численности и характера организации человеческих популяций. Это могут быть изоляты, демы, нации, наднациональные ассоциации (различающиеся по способу производства, укладу жизни) и, наконец, человечество в целом.Адаптации человека к условиям обитания имеют частично экологическую, но главным образом социальную природу. Адаптации формируются по отношению к факторам как природной, так и искусственной среды, поэтому они носят не только экологический, но и социально-экономический характер. Экологические и социально-экономические адаптации дополняются психологическими адоптациями, поскольку каждый человек индивидуален. Индивидуальные и групповые адаптации человека, в отличие от биологических адаптации растений и животных, обеспечивают наряду с выживанием и воспроизведением потомства выполнение им социальных функций, важнейшей из которых является общественно - полезный труд. Индивидуальные и групповые адаптации человека включают оптимизацию условий жизни и производственной деятельности (устройство жилищ и других помещений, конструкцию одежды, организацию питания и водоснабжения, рациональный режим труда и отдыха, сознательную тренировку организма и др.).Хотя в основе адаптации человека лежат социально-экономические механизмы, важная роль принадлежит также естественным приспособительным и защитным механизмам, которые составляют биологическое наследство человека.Адаптации человека к факторам окружающей среды носят опосредованный характер. Действие экологических факторов всегда опосредовано результатами производственной деятельности людей. В процессе труда человек своей собственной деятельностью опосредует, регулирует и контролирует обмен веществ между своим организмом и природой. К тому же естественные экосистемы в настоящее время всё больше вытесняются антропогенными экосистемами, в которых человек является абсолютно доминирующим экологическим фактором.
98. Антропогенные экосистемы как результат индустриализации, химизации, урбанизации, развитие транспорта, выход в космос.Отличительная черта антропогенных экосистем состоит в том, что доминирующий экологический фактор в них представлен сообществом людей и продуктами его производственной и общественной деятельности. Благодаря целенаправленной преобразующей активности человека в отношении природы антропогенным экосистемам свойственно преобладание искусственной среды над естественной. Преобразующая деятельность людей в современных условиях строится, как правило, на основе предварительного планирования. Однако предполагаемый ход развития антропогенных экосистем нередко искажается благодаря действию стихийных, а также неучтенных человеком заранее сил. В результате возникают состояния «экологических стрессов», которые могут в перспективе привести к экологическому кризису. Важнейшие современные антропогенные системы – города, сельские поселения, транспортные коммуникации – характеризуются определенным сочетанием биоприродных и хозяйственно-культурных условий. 99. Эволюция биосферы. Учение акад. В.И. Вернадского.(от греч. βιος — жизнь и σφαῖρα — сфера, шар) — оболочка Земли, заселённая живыми организмами, находящаяся под их воздействием и занятая продуктами их жизнедеятельности; «пленка жизни»; глобальная экосистема Земли.Термин «биосфера» был введён в биологии Жаном-Батистом Ламарком в начале XIX в., а в геологии предложен австрийским геологом Эдуардом Зюссом в 1875 году[1].Целостное учение о биосфере создал русский биогеохимик и философ В. И. Вернадский. Он впервые отвёл живым организмам роль главнейшей преобразующей силы планеты Земля, учитывая их деятельность не только в настоящее время, но и в прошлом.Существует и другое, более широкое определение: Биосфера — область распространения жизни на космическом теле. При том что существование жизни на других космических объектах, помимо Земли пока неизвестно, считается что биосфера может распространяться на них в более скрытых областях, например, в литосферных полостях или в подлёдных океанах. Так, например, рассматривается возможность существования жизни в океане спутника Юпитера Европы.Развитие наблюдается лишь в живом веществе и связанным с ним биокосном. В косном веществе нашей планеты эволюционный процесс не проявляется[1].Зарождение жизниЖизнь на Земле зародилась ещё в архее — примерно 3,5 млрд лет назад. Такой возраст имеют найденные палеонтологами древнейшие органические остатки. Возраст Земли как самостоятельной планеты Солнечной системы оценивается в 4,5 млрд лет. Таким образом, можно считать, что жизнь зародилась ещё в юношескую стадию жизни планеты. В архее появляются первые эукариоты — одноклеточные водоросли и простейшие. Начался процесс почвообразования на суше. В конце архея появился половой процесс и многоклеточность у животных организмов.Будущее биосферыС течением времени биосфера становится всё более неустойчивой. Существует несколько трагичных для человечества преждевременных изменений состояния биосферы, некоторые из них связаны с деятельностью человечества. Некоторые философы, например, Дэвид Пирс выступают за модификацию биосферы с целью избавления от страданий всех живых существ и создание в буквальном смысле рая на земле (см. одно из значений слова аболиционизм).
100. Основные формы биологических связей в антропобиогеоценозах. Паразитизм как биологический феномен. Всякий организм живет в определенной среде, в которой кроме него обитают и другие существа. Любое живое существо находится в тесной зависимости от окружающей среды, к обитанию в которой оно приспособилось в процессе эволюционного формирования соответствующего вида В отношении факторов питания всех животных можно разделить на три группы: - плотоядных, или животных, которые пожирают другие организмы или даже себе подобных (хищные); - животных, питающихся падалью, разлагающимися органическими веществами (личинки трупной мухи, комнатной мухи, некоторые млекопитающие и птицы); - животных-паразитов, которые питаются за счет соков тканей тела или переваренной пищей живых хозяев. Хищники, нападая на добычу, убивают и пожирают ее. Паразиты же пользуются своими хозяевами многократно в течение всей жизни или жизни своих хозяев. Поэтому паразиты или вовсе не губят своего хозяина, или же не приводят к его смерти в течение продолжительного срока вследствие вызываемого ими заболевания. Паразитизм - это форма сожительства, при которой один организм - паразит живет за счет организма другого вида - хозяина. Паразитизм - это форма сожительства двух генетически разнородных организмов разных видов, при которой один организм (паразит) использует другого (хозяина) как источник питания и среду существования, причиняя ему вред, но, как правило, не уничтожая его. Формы паразитизма чрезвычайно разнообразны. Различают облигатный (постоянно встречающийся, обязательный) и факультативный (возможный, необязательный), временный и постоянный, истинный и ложный, экто- и эндопаразитизм.Паразитизм широко распространен в природе. Он встречается почти во всех типах животного царства. Особенно много паразитов имеется в типах простейших, плоских и круглых червей, членистоногих. Согласно данным В. А. Догеля (1947 г.) в природе около 60-65 тыс. видов являются животными-паразитами. Широкое распространение и разнообразие видов паразитизма позволяет считать, что эта форма сожительства возникла в природе в различных систематических группах животных независимо одна от другой.
101. Принципы взаимодействия паразита и хозяина на уровне особей. Жизненные циклы паразитов. Чередование поколений и феномен смены хозяев. Понятие о био- и геогельминтах. Гельминтам со сменой хозяев в жизненном цикле для его реализации необходимо пройти через строго обязательный круг промежуточных и окончательного хозяев. Это минимально необходимая, и в некоторых случаях достаточная основа паразитарной системы (Догель, 1962, Беклемишев, 1956, 1970). Однако у многих эндопаразитических гельминтов при широкой специфичности личиночных и ювинильных стадий количество особей хозяев на данной стадии личиночного развития может быть достаточно велико (от 2-3 и в условиях океанской пелагиали до 5-6 и более). Особенно характерно это явление для океанических гельминтов, реализующих свои жизненные циклы по трофическим каналам биотической структуры сообществ (Нигматуллин, 1987; Marcogliese, 1995). Эти хозяева могут быть как разно-размерными представителями одного вида, так и представителями разных типов, классов и т.д. Это явление связано с последовательным рядом актов поедания хищниками зараженных жертв (принцип "эстафетной передачи"). В одних случаях за счет тех или иных причин (высокий иммунный барьер и т.п.) гельминты, попав в хозяина живыми и активными, погибают (Шигин, 1981). В других случаях в первом подходящем хищнике (хозяине) у личинок происходят морфо-физиологические изменения, специфичные для данной стадии онтогенеза (промежуточный хозяин), а в последующих хозяевах они могут жить, не претерпевая дальнейшего развития (дополнительные паратенические = транспортные хозяева). Дальнейшее онтогенетическое развитие возможно лишь при попадании личинки гельминта в промежуточных хозяев следующего уровня, которые необходимы для прохождения следующей стадии онтогенеза. При этом, благодаря трофическим, пространственным или временным особенностям экологической ниши данного хозяина, личинки гельминтов могут или оставаться в нем неопределенно долго (в соответствии с продолжительностью онтогенеза хозяина), или попадать к хищникам (хозяевам) не имеющих трофических контактов с необходимыми для дальнейшего развития хозяевами. Последовательный ряд особей дополнительных хозяев для данной личиночной стадии развития гельминта с точки зрения необходимости прохождения через строго генетически детерминированный круг хозяев (морфогенез и половое созревание) не обязателен. Это явление в историческом аспекте вторичное (но возможны и варианты параллельного его формирования наряду с морфо-физиологически обязательными хозяевами), и обусловлено значительной сложностью и удлинением трофических сетей сообществ, особенно в океанической пелагиали. Личинки гельминтов приживаются у этих новых хозяев, и далее ими осваиваются при наличии преадаптации в виде широкой специфичности личиночных стадий гельминтов, или же ее "направленного" расширения (канализированный отбор при увеличении вероятности достижения окончательных хозяев при помощи нового дополнительного хозяина), а также отсутствия у этих новых хозяев непреодолимого иммунного барьера и существенных предпосылок для формирования антигенного паритета паразита и хозяина (обзор: Сергеева, Беэр, 2000). Далее, если новый хозяин имеет "подходящие" биоценотические связи, он "используется" паразитом для оптимизации путей достижения окончательного хозяина в соответствии с особенностями данной экосистемы и положения в ее пространственной и трофической структуре промежуточных и окончательных хозяев с одной стороны и "нового" хозяина - с другой. Наиболее далеко зашедшее взаимоотношение с такими хозяевами - использование их в качестве паратенических (транспортных) хозяев. Это явление приобретает особенно важное значение в случае значительного накопления личинок гельминтов в теле хозяина. Последних в отечественной литературе обычно называют резервуарными хозяевами, и они являются источником массового заражения следующих хозяев (Скрябин, Шульц, 1937, 1940; Рыжиков, 1954; Шульц, Давтян, 1955; Догель, 1962; Гинецинская, 1968; Шульц, Гвоздев, 1972). В этих резервуарных хозяевах иногда сосредоточена основная численность личинок данной стадии жизненного цикла популяции гельминта. В конечном варианте развития этой тенденции "освоения" новых хозяев, некоторые из них могут стать с экологической точки зрения необходимыми (облигатны!) для замыкания жизненного цикла данной популяции гельминта, и далее попасть в категорию промежуточных и или реже окончательных хозяев [3].Промежуточные и транспортные хозяеваКонцепция промежуточных хозяев сформировалась в 1840-1890-х годах благодаря основополагающим работам К. Эшрихта, Я. Стеенструпа, К. Зибольда, П. Бенедена, Ф. Кюхенмейстера, А. Лооса и, особенно, Р. Лейкарта (Мордвилко, 1908; Чеснова, 1978). При этом окончательными хозяевами называли тех животных, которые дают приют половозрелым формам, а промежуточными хозяевами - молодым стадиям паразитов (Мордвилко, 1908; Графф, Линстов, 1910; Caullery, 1952). В самом общем виде это деление на две основные группы хозяев паразитов (т.е. дефинитивных и всех остальных - промежуточных) сохраняется до настоящего времени; по крайней мере, без обозначения других групп хозяев личиночных форм паразитов, присутствует во множестве паразитологических публикаций 1920-1970-х годов (Caullery, 1952; Чеснова, 1978). В течение 20 века для промежуточных хозяев были введены уточняющие определения, в основном, связанные со стадийным морфо-физиологическим развитием, бесполым или партеногенетическим размножением или какими либо подготовительными физиологическими процессами, необходимыми для дальнейшего развития (Догель, 1941, 1947, 1962; Чеботарев, Ратнер, 1962; Шульц, Гвоздев, 1972; Odening, 1976). В то же время, начиная с 1920-1930-х годов постепенно стало увеличиваться количество наименований для хозяев личиночных форм паразитов, выделяемых с самых разных позиций (Odening, 1976). При этом главной характеристикой промежуточного хозяина остается обязательность его участия в жизненном цикле паразита независимо от того происходит в нем морфогенез или нет (Догель, 1962). Эта обязательность выражается как в необходимости данного хозяина для прохождения морфогенеза или физиологических преобразований, личиночного размножения (т.е. физиологическая обусловленность), так и для передачи дефинитивных хозяевам (экологическая обусловленность). Эта двойственность и определяет «рыхлость» понятия промежуточный хозяин. Все это вместе взятое во многом обусловило современную ситуацию «хаоса наименований» хозяев паразитов. Промежуточные хозяева - весьма гетерогенная группа хозяев личинок и неполовозрелой молоди гельминтов с самым разным значением в жизненных циклах последних. Во избежание существующей путаницы термин «промежуточные хозяева» предлагается оставить только за хозяевами в которых происходит морфогенез личинок и ювенильных форм, характерный для смены стадий онтогенеза; физиологическая подготовка для полового созревания в дефинитивном хозяине; личиночное бесполое или партеногенетическое размножение т.е. они являются физиологически обязательными хозяевами. При этом экологическая обусловленность этих промежуточных хозяев, в тех случаях, когда они непосредственно поедаются дефинитивным хозяевам и «заражают» их, является естественным «фоном». Такое четкое деление "промежуточных хозяев" по наличию - отсутствию в них морфогенеза личинок гельминтов на собственно промежуточных (actual intemediate hosts) и вспомогательных (auxiliary hosts) хозяев было сделано еще в 1932 г. в известном "Руководстве по гельминтологии" (Sprehn, 1932; цит. по: Odening, 1976, p. 44).Это сужение понятия промежуточный хозяин вполне соответствует и первоначальному определению В.А. Догеля (1941, с. 145): «Тот же организм, в котором паразит живет в молодом, личиночном состоянии, называется промежуточным хозяином. Последнее определение нуждается, однако, в некотором уточнении. Дело в том, что не всякое животное, в котором молодой паразит проводит часть своего развития, может с одинаковым основанием быть названо промежуточным хозяином. В полном смысле слова промежуточным хозяином можно считать только то животное, прохождение через организм которого, либо является необходимым и обязательным условием для дальнейшего развития паразита в организме основного хозяина, либо в организме которого паразит проходит ту или иную существенную часть своего развития (часть метаморфоза, партеногенетическое размножение и др.). В тех случаях, когда такого развития паразита в промежуточном хозяине не происходит, правильнее говорить о механической передаче и называть эту промежуточную инстанцию не промежуточным хозяином, а механическим переносчиком». Это также соответствует определению (правда, довольно «расплывчатому») промежуточных хозяев Р.С. Шульца и Е.В. Гвоздева (1972, с. 11): «… это такие хозяева, в которых совершается развитие начальных и промежуточных стадий метаксенных паразитов». Для собственно экологически и эпидемиологически обязательных хозяев неполовозрелых гельминтов (Беклемишев, 1956) существует широкий спектр перекрывающихся названий: резервуарные, паратенические, транспортные, дополнительные и вставочные хозяева. На самом деле количество этих терминов значительно больше (Odening, 1976), и здесь приведены лишь наиболее широко используемые в отечественной и англоязычной литературе.Особая сложность существует с термином резервуарные хозяева. Это связано с тем, что в отечественной и западной литературе понятие о резервуарном паразитизме развивалось независимо и в разных областях паразитологии. В советской гельминтологической литературе этот термин начал использоваться с конца 1930-х годов (Скрябин, Щульц, 1937, 1940) и особенно широко с 1950-х годов (Рыжиков, 1954; Шульц, Давтян, 1955; Догель, 1962; Чеботарев, Ратнер, 1962; Гинецинская, 1968; Odening, 1976) для обозначения хозяев, в которых отсутствует развитие личинок гельминтов, но происходит их накопление, переживание в течение длительного времени в инвазионном состоянии, и в той или иной степени повышение вероятности передачи их окончательным хозяевам по трофическим каналам (Догель, 1962; Шульц, Гвоздев, 1972) или даже являющимся экологически обязательным звеном жизненного цикла гельминтов (Гаевская. Нигматуллин, 1976, 1981)). В западной литературе этот термин (reservoir host) широко распространен в эпидемиологической литературе с иным наполнением. Резервуарный хозяин - это инфицированный дефинитивный хозяин, который служит источником постоянного или периодического заражения других животных или человека. В качестве классического примера обычно приводится случай, когда в качестве резервуарного хозяина выступает антилопа зараженная трипаносомой, вызывающей сонную болезнь у человека (Noble et al., 1989). В связи с этими разночтениями данного термина и обычны ситуации непонимания отечественных работ зарубежными коллегами, и наоборот. По-видимому, в сложившейся ситуации при все более возрастающей интернационализации паразитологии, желательно избегать использования этого термина в понимании К.И. Скрябина и Р.С. Шульца (1937, 1940) и К.М. Рыжикова (1954). К этому побуждают также, с одной стороны, и значительные разночтения в трактовке этого понятия в русскоязычной литературе, и с другой, широкое использование в нашей литературе в последнее два десятилетия терминов паратенические и транспортные хозяева. Термин паратенические хозяева был введен в обиход Дж. Баэром (Baer, 1951), он претерпел определенную эволюцию быстро войдя в широкий обиход международного сообщества паразитологов и означает в последнюю четверть века то же, что и отечественный термин “резервуарные хозяева” без акцента на количественном накоплении личинок в теле данного хозяина (Шульц, Гвоздев, 1972; Odening, 1976; К.Г. Галактионов, личное сообщение). Паратенический хозяин - потенциальный промежуточный хозяин, в котором не происходит развития незрелого гельминта, он способствует попаданию личинки в окончательного хозяина (Baer, 1951; Odening, 1976; Noble et al., 1989). Правда К. Оденинг (Odening, 1976, p. 51-53) категорически не согласен с определением паратенических хозяев как потенциальных промежуточных хозяев, однако если принять это определение именно как потенциальную возможность в эволюции жизненных циклов гельминтов оно вполне корректно. Здесь же К. Оденинг подчеркивает, что по ясности определения и благодаря международному признанию термин паратенические хозяева для обозначения экологически обусловленных хозяев наиболее предпочтителен. Термин транспортные хозяева был впервые использован Ф. Фуллеборном (Fulleborn, 1923: цит. по: Odenenig, 1976, p. 44) и означал «промежуточных хозяев», которые «транспортируют» личинок гельминтов следующим хозяевам при их поедании последними. Ныне они определяются как организмы, которые «переносят» неразвивающихся паразитов к следующим хозяевам (Noble et al., 1989). В настоящее время термины паратенические и транспортные хозяева являются во многом синонимичными и соответственно могут быть использоваться как равноценные (Schmidt, Roberts, 1989). Что касается термина вставочные хозяева (Скрябин, Шульц, 1937, 1940), он применим при описании конкретных ситуаций связанных с появлением в жизненном цикле гельминта нового, важного для реализации цикла, паратенического хозяина, и имеет узкое применение.
В целом же все экологически обусловленные паратенические хозяева (наряду с факультативными собственно промежуточными и дефинитивными) с точки зрения их роли в жизненных циклах гельминтов могут быть в целом охарактеризованы как дополнительные хозяева по отношению к обязательным собственно промежуточным и дефинитивным хозяевам. В историческом плане дополнительные хозяева, как правило, вторичны, связи с ними гельминтов более молоды и в конечном счете экологически обусловлены возможностью оптимизации достижения дефинитивного хозяина [3].5. Классификация хозяевВ соответствии с вышеизложенным, для изучения реальной структуры жизненных циклов и паразитарных систем гельминтов имеет смысл выделить четыре главные группы хозяев, и соответственно четыре уровня структур хозяев в жизненном цикле их гельминтов. 1. Первичная структура хозяев. Генетически и соответственно физиологически жестко детерминированный круг обязательных хозяев, в которых происходят обязательные подготовительные морфо-физиологические стадийные процессы онтогенетического развития и достижение взрослого половозрелого состояния гельминтов. Эти обязательные (Догель, 1962) (=специфичные: Галактионов, Добровольский, 1998) хозяева необходимы для завершения жизненного цикла, через них «проходит» в процессе жизненного цикла основная часть популяции гельминта. По месту в жизненных циклах гельминтов выделяются две основные подгруппы хозяев: 1.1. промежуточные обязательные (I-е, II-е, III-и) и 1.2. окончательные обязательные хозяева, последовательно сменяемые гельминтом в процессе онтогенеза. 2. Вторичная структура хозяев. Факультативные (= малоспецифичные Галактионов, Добровольский, 1998) хозяева в которых также происходит полный или частичный морфогенез и достижение половой зрелости, но они не являются обязательными хозяевами и их заражение данным гельминтом носит случайный характер. Это обычно близкородственные виды по отношению к хозяевам первичной структуры с отличающимися от них экологическими, и в первую очередь, трофическими характеристиками. Они не являются полноценными и необходимыми членами паразито-хозяинных систем, но могут быть использованы частью гельминтов данной популяции для замыкания онтогенеза. В реальных природных ситуациях их выделение и дифференцировка от представителей первой группы обычно представляет большую трудность и возможна лишь при детализированных многолетних эколого-паразитологических исследованиях ориентируясь на низкие показатели экстенсивности, и в меньшей степени, интенсивности инвазии. Они так же, как и представители первой группы, по месту в жизненном цикле гельминтов разделяются на две подгруппы: 2.1. промежуточные факультативные (I-е, II-е, III-и) и 2.2. окончательные факультативные хозяева. 3. Третичная структура хозяев. Экологически обусловленный круг дополнительных (вставочных) паратенических хозяев, в которых паразитируют личинки или ювенильные особи гельминтов. С точки зрения прохождения морфо-генетических процессов онтогенетического развития гельминтов они не обязательны, но экологически в той или иной степени способствуют реализации их жизненных циклов. Дальнейшее заражение также возможно и после смерти дополнительных хозяев при поедании их трупов. По роли в реализации жизненных циклов гельминтов выделяются две подгруппы. 3.1. Дополнительные транспортные хозяева с аггломерацией личинок паразитов - накопительные (= резервуарные по: Рыжиков, 1954) хозяева. В них происходит накопление личинок или ювенильных стадий гельминтов и они необходимы для поддержания репродуктивных частей их популяций на оптимальном уровне (Догель, 1962; Шульц, Гвоздев, 1972). В крайнем варианте, при удлинении пищевой цепи, по которой циркулирует популяции данного гельминта, без участия накопительных хозяев замыкание жизненного цикла популяции гельминта в полной мере невозможно (то есть поддержание численности популяции на существующем уровне). В океанических экосистемах цестоды, нематоды и в меньшей степени дициемидные трематоды на пути к дефинитивным хозяевам - хищникам, стоящим на вершине пищевой сети (акулы, дельфины, скомброидные и ксифоидные рыбы), нектонные кальмары как транспортные накопительные хозяева просто обязательны: прямые трофические контакты между I-II промежуточными хозяевами и дефинитивными отсутствуют. Кальмары, являясь обычной пищей этих хищников, как раз и осуществляют эту роль промежуточного звена, и без них реализация жизненных циклов этих гельминтов было бы невозможно (Гаевская, Нигматуллин, 1976, 1981). 3.2. Дополнительные транспортные хозяева без накопления личинок гельминтов. Они осуществляют передачу личинок гельминтов следующим хозяевам без их значительной аккумуляции в своем теле. Хозяева этих двух подгрупп являются паратеническими или транспортными т.е. способствующими циркуляции и передаче личинок гельминтов последующим хозяевам по трофической сети. Эти паратенические (от paratenio = продлеваю) хозяева третичной структуры находятся в биотической структуре сообществ на магистральных, или же второ- и третьестепенных "путях" к следующему дополнительному, промежуточному или же окончательному хозяину. В реальных природных ситуациях при начальных описательных паразитологических исследованиях разделение хозяев на собственно промежуточных (особь хозяина, в которой происходит морфогенез) и дополнительных хозяев крайне затруднено. Однако оно возможно при вовлечении паразитологических исследований в общий контекст синэкологических (и в первую очередь трофологических) работ. 4. Четвертичная структура хозяев. Экологически обусловленные латерально-терминальные хозяева, своего рода экологические тупики ("ловушки") в «потоке» жизненного цикла гельминтов. Это своего рода «фиктивные» (Шульц, Давтян, 1955) или случайные (по Marcogliese, 1995, p. 337: Accidental hosts are those which may acquire infection, but are not involved in transmission and reproduction of the parasite) хозяева. Однако, последний термин вряд ли следует использовать для этой группы хозяев, так как в литературе устоялась более общая его трактовка, включая и редко заражаемых хозяев, но участвующих в реализации жизненного цикла паразита (Odening, 1976). В этих хозяевах можно обнаружить гельминтов, но они не участвуют в дальнейшей передаче паразитов следующим хозяевам, подходящим для реализации их жизненных циклов. Среди них по времени жизни в хозяине выделяются следующие две подгруппы. 4.1. Каптивные или тупиковые хозяева (хозяева - экологические тупики). Гельминты в них живут достаточно долго (крайний вариант - до конца онтогенеза хозяина), но вероятность заражения следующих хозяев (или размножения взрослых форм паразита) близка к нулю. Эти хозяева находятся за пределами функциональных частей трофической структуры сообществ, в которых в норме реализуется жизненные циклы гельминтов. Заражаются они или случайно, или закономерно поедая жертв - обычных промежуточных и дополнительных хозяев. При закономерном заражении для популяций гельминтов каптивные хозяева являются и своеобразной "стерильной зоной выселения", и неизбежной платой за использование трофической структуры сообществ сетевого типа для реализации своих жизненных циклов. В этом случае популяции массовых животных низших трофических уровней используют в пищу самые разнообразные хищники, входящие в различные функциональные подсистемы сообществ. 4.2. Хозяева-убийцы. Это псевдохозяева (=элиминативные хозяева по: Галактионов, Добровольский, 1998, с. 327). После поедания зараженных жертв хозяевами-убийцами, гельминты какое то время находятся в теле хозяина (обычно в пищеварительном тракте) в живом или постмортальном состоянии с нормальным внешним обликом. В результате эти животные могут быть представлены в списках дополнительных, промежуточных или окончательных хозяев тех или иных гельминтов, и эти данные могут использоваться при реконструкции их жизненных циклов и некорректном описании паразитарных систем. Однако эти гельминты вскоре с неизбежностью погибают или за счет действия иммунной системы хозяина, или реже во внешней среде после прохождения транзитом через пищеварительную систему хозяина [3].Биогельминты (био- + гельминты) — общее название гельминтов, для завершения жизненного цикла которых необходима смена хозяев. Вызываемые ими заболевания — биогельминтозы. Биогельминты могут развиваться со сменой двух или трех хозяев, принадлежащих к разным видам организмов, даже порой далеко отстоящих друг от друга в систематическом положении. Например, лентец широкий завершает полностью свой жизненный цикл, или эпидемиологическую цепь, последовательно в организме трех хозяев: веслоногих рачков, рыбы, человека или некоторых плотоядных. К биогельминтам относятся трематоды, цестоды и некоторые нематоды.Биогельминты (от био... и гельминты), группа паразитических червей, которые во взрослом состоянии паразитируют в т. н. дефинитивном (окончательном) хозяине (человек, животное), а на стадии личинки — в промежуточном хозяине (разные животные), в отличие от геогельминтов, развивающихся без промежуточного хозяина. Например, взрослый бычий цепень (см. Цепни) паразитирует в кишечнике человека, а его личинки — финки — развиваются в мышцах крупного рогатого скота. Заражение человека происходит при употреблении в пищу полусырого финнозного мяса. К Б. относятся также лентец широкий, печеночная двуустка, трихина, эхинококк и многие другие гельминты.Геогельминты (от гео... и гельминты), группа паразитических червей человека и животных, развивающихся (в отличие от биогельминтов) без промежуточных хозяев. Яйца Г. с фекалиями попадают в почву, где развиваются в тёплое время года до стадии личинок. Заражение человека происходит либо через немытые овощи, фрукты, руки, на которых находятся инвазионные яйца (например, аскариды, власоглава человеческого, острицы), либо при непосредственном контакте с землёй, где живут личинки (например, анкилостомид).
102. Трансмиссивные и природно-очаговые заболевания. Учение академика Е.Н.Павловского о природной очаговости паразитарных болезней. Природно-очаговыми инфекционными болезнями (ИБ) называют заболевания, при которых источник инфекции находится в природе. Чаще всего это теплокровные дикие животные, для некоторых болезней — кровососущие насекомые, главным образом клещи. Ареал каждой инфекции ограничен определенной эколого-географической местностью. Передача возбудителя болезни человеку происходит через укусы инфицированных насекомых (клещей, блох, комаров, москитов и др.); при употреблении загрязненной больными животными воды, пищи; через предметы домашнего обихода; при прямом контакте — соприкосновении с возбудителем.1Трансмиссивные болезни (лат. transmissio — перенесение на других) — заразные болезни человека, возбудители которых передаются кровососущими членистоногими (насекомыми и клещами).Трансмиссивные болезни включают более 200 нозологических форм, вызываемых вирусами, бактериями, риккетсиями, простейшими и гельминтами. Часть из них передаётся только с помощью кровососущих переносчиков (облигатные трансмиссивные болезни, например сыпной тиф, малярия и др.), часть различными способами, в том числе и трансмиссивно (например, туляремия, заражение которой происходит при укусах комаров и клещей, а также при снятии шкурок с больных животных).[править]ПереносчикиВ передаче возбудителей трансмиссивных болезней участвуют специфические и механические переносчики. В специфических переносчиках возбудитель либо размножается и накапливается (например, вирусы в теле клещей), либо созревает до инвазионной стадии (например, личинки филярий в комарах, мошках, слепнях, малярийные плазмодии в теле комаров). Как размножение, так и развитие возбудителей в теле специфических переносчиков может происходить только при достаточно высокой, строго определённой температуре для каждой пары паразит — переносчик. Так, половое размножение, или спорогония, возбудителя трёхдневной малярии продолжается 19 дней при 20 °C и 6½ дней при температуре 30 °C, и только после этого в слюнных железах переносчика появляется инвазионная стадия паразита — спорозоит. Передача возбудителя переносчиком осуществляется различными способами. Наиболее часто возбудитель проникает со слюной, которую переносчик вводит в место укола перед началом кровососания. Так вводятся спорозоиты малярийного паразита. Перенос риккетсий вшами (см. Сыпной тиф эпидемический) и трипаносом триатомовыми клопами происходит во время дефекации переносчика; возбудитель из фекалий проникает затем в ранку на месте укола. Возбудитель вшиного возвратного тифа (Borrelia recurrentis) скапливается в теле вши и может проникнуть в организм человека лишь при раздавливании вши руками и втирании ее гемолифмы в кожу при расчёсах. Многие переносчики передают лишь одного, специфического для них паразита. Так, мухи цеце передают лишь трипаносому — возбудителя сонной болезни человека, мошки — возбудителя онхоцеркоза, триатомовые клопы — возбудителя американского трипаносомоза (болезни Шагаса). Москиты могут переносить возбудителей лейшманиозов и лихорадки паппатачи. Наиболее разнообразные связи с паразитами установились у комаров и клещей. Комары семейства Culicidae передают многие виды вирусов, в том числе возбудителя жёлтой лихорадки, лихорадки денге, вирусных энцефалитов (см. Энцефалиты комариные). Они являются переносчиками также малярии и филяриатозов. Иксодовые клещи передают вирусы, в том числе возбудителей клещевого энцефалита и геморрагических лихорадок, риккетсий — возбудителей пятнистых лихорадок, североазиатского сыпного тифа (см. Североазиатский клещевой риккетсиоз) и др., а также спирохет. В переносе возбудителей болезни у многих групп насекомых участвуют только самки. У мухи цеце, вшей и у всех групп клещей кровососами и переносчиками являются особи обоего пола.В ряде случаев возбудитель болезни передается переносчиком своему потомству. Такая трансовариальная передача выявлена у комаров, инфицированных вирусами, у клещей, инфицированных вирусами и спирохетами и у москитов, инфицированных флебовирусами.В организме механических переносчиков возбудители не развиваются и не размножаются. Попавший на хоботок, в кишечник или на поверхность тела механического переносчика возбудитель передается непосредственно (при укусе) либо путем контаминации ран, слизистых оболочек хозяина или пищевых продуктов. Самыми распространенными механическими переносчиками являются мухи семейства Muscidae, которые известны как переносчики вирусов, бактерий, простейших, гельминтов.
103. Тип Простейшие. Классификация. Характерные черты организации. К типу Простейшие относят организмы, тело которых состоит из одной клетки, функционирующей, однако, как целый организм. Клетки простейших способны к самостоятельному питанию, передвижению, защите от врагов и к переживанию неблагоприятных условий. Питание простейших происходит с помощью пищеварительных вакуолей, осуществляется за счет фаго - или пиноцитоза. Остатки непереваренной пищи выбрасываются наружу. Некоторые простейшие содержат хлоропласты и способны питаться за счет фотосинтеза.Большинство простейших имеют органеллы передвижения: жгутики, реснички и псевдоподии (временные подвижные выросты цитоплазмы). Формы органелл движения лежат в основе систематики простейших.Размножение простейших осуществляется обычно разными формами деления — разновидностями митоза. Характерен также половой процесс: в виде слияния клеток — копуляция—или обмен частью наследственного материала — конъюгация.Большинство простейших имеют одно ядро, но встречаются и многоядерные формы. В жизненном цикле большинства простейших выделяют стадию трофозоита — активно питающуюся и перемещающуюся форму — и стадию цисты. Циста — неподвижная форма жизненного цикла простейших, покрытая плотной оболочкой и характеризующаяся резко замедленным обменом веществ. Паразитические простейшие инцистируются, попадая во внешнюю среду. При попадании цисты в благоприятные условия происходит эксцистирование и простейшее начинает активно функционировать в состоянии трофозоита. В настоящее время известно около 10 000 видов простейших. Болезни, вызываемые простейшими, называют протозойными. Большинство простейших имеют время генерации от 6 до 24 ч. Один паразитический организм способен, размножившись, привести к гибели своего хозяина. В этом плане простейшие—возбудители заболеваний — сходны с возбудителями инфекционных болезней, например с патогенными бактериями и вирусами.Медицинское значение имеют простейшие, относящиеся к классам Саркодовые, Жгутиковые, Инфузории и Споровики. 104. Дизентерийная амеба. Систематическое положение, морфология, цикл развития, обоснование лабораторной диагностики, профилактика.
Дизентерийная амеба (лат. Entamoeba histolytica) — простейшее, паразитирующее в толстой кишке человека и вызывающее тяжелое заболевание — амебную дизентерию (амебиаз). Распространено заболевание почти во всех частях света, особенно в странах с тропическим климатом. Жизненный цикл дизентерийной амебы проходит в несколько стадий. Различают 3 формы дизентерийной амебы: вегетативная мелкая (просветная) форма; вегетативная крупная (тканевая) форма; циста. Мелкая вегетативная форма питается бактериями, остатками непереваренной пищи кишки и обнаруживается у больных в период ремиссии или у носителей; интенсивно размножается делением надвое, образует цисты. В ослабленном организме человека (при инфекции, переохлаждении и пр.) мелкая вегетативная форма может переходить в крупную патогенную вегетативную форму, которая живет в стенке кишки, выделяет протеолитическийфермент, разъедающий стенку кишки с образованием язв. Питается эритроцитами, может питаться осмотически. Эту форму находят в остром периоде заболевания. С током крови дизентерийная амеба может попадать в различные органы из толстой кишки и обусловливать амебиаз печени и др. органов. Цисты дизентерийной амебы округлой формы с двойной тонкой оболочкой. Молодая циста имеет одно ядро, которое вскоре делится дважды, в результате чего циста становится четырехъядерной. Человек заражается, проглатывая цисты вместе с загрязненной водой и пищей. Диагноз ставится на основании выявления вегетативных форм и цист при микроскопическом исследовании фекальных масс больных или гноя из абсцессов пораженных органов, мокроты и др. выделений. Профилактические мероприятия: соблюдение правил личной гигиены, защита пищевых продуктов от загрязнения
105. Трихомонады, трипаносомы, лямблии. Систематика, морфология, циклы развития, пути заражения, методы лабораторной диагностики.Трихомонады. Разделяется на кишечную и влагалищную. Кишечная обитает в кишечнике, размножаясь в кишечнике человека, трихомонада может вызывать недолгие поносы. В половых органах человека обитает влагалищная трихомонада Trichomonas vaginalis (кл. Жгутиковые) — возбудитель трихомоноза. Длина этого паразита 14—30 мкм. Форма тела грушевидная. На переднем конце находятся четыре жгутика. До середины клетки доходит также небольшая ундулирующая мембрана. По середине тела тянется аксостилъ, выступающий из клетки на ее заднем конце. Характерна форма ядра, овального, заостренного с двух концов и напоминающего косточку сливы. В пищеварительных вакуолях располагаются лейкоциты, эритроциты и бактерии, которыми этот паразит питается. Цист не образует. Эта трихомонада обитает у женщин во влагалище и в шейке матки, а у мужчин — в мочеиспускательном канале, мочевом пузыре и в предстательной железе. Зараженность женщин достигает 20—40%, мужчин — 15%. Серьезных повреждений хозяину | эта трихомонада не наносит, но, тесно контактируя с эпителием мочеполовой системы, она вызывает возникновение мелких воспалительных очагов под эпителиальным слоем и слущивание поверхностных клеток слизистой оболочки. Через нарушенную эпителиальную выстилку в просвет органа поступают лейкоциты. У мужчин заболевание обычно завершается спонтанным выздоровлением примерно через 1 мес. У женщин трихомоноз может протекать несколько лет. Лабораторная диагностика — обнаружение живых подвижных трихомонад в мазке из выделений мочеполовых путей. Профилактика—соблюдение правил личной гигиены при половых контактах.Трипаносомы — род паразитических одноклеточных простейших семейства трипаносомовые, которые паразитируют на различных хозяевах и вызывают многие заболевания, среди которых сонная болезнь и болезни Чагаса Естественным резервуаром трипаносом в основном являются млекопитающие, переносчиком — насекомые. Муха цеце является переносчиком Trypanosoma brucei - возбудителя сонной болезни. Триатомовые клопы являются переносчиками Trypanosoma cruzi — возбудителя болезни Чагаса.Ля́мблии, или жиа́рдии (лат. Giardia) — паразитические жгутиковые протисты из отряда дипломонадид (Diplomonadida), паразитирует в тонком кишечнике человека и многих других млекопитающих, а также птиц. Один из видов этого рода — кишечная лямблия (Giardia intestinalis) (синонимы — Giardia lamblia и Giardia duodenalis) — возбудители лямблиоза человека.Размножаются в активном состоянии (на стадии трофозоита) путем продольного деления надвое. Во внешнюю среду с фекалиями хозяина попадают как трофозоиты, так и образующиеся в кишечнике цисты. Выживают во внешней среде только цисты, попадающие в организм новых хозяев фекально-оральным путем (с зараженной водой или пищей).
106. Лейшмании. Систематическое положение, морфология, циклы развития. Пути заражения, профилактика, лабораторная диагностика. Лейшмании (от имени У. Лейшмана) — род паразитических простейших, вызывающих лейшманиозы. Лейшмании, как и все трипаносоматиды, являются облигатными паразитами. Жизненный цикл лейшманий включает двух хозяев: млекопитающее и насекомое (москит).Москиты заражаются лейшманиями, когда пьют кровь заражённого млекопитающего. Кровью питаются только самки москитов. Лейшмании вместе с проглатываемой кровью проникают в пищеварительный канал москита. В задней части средней кишки вокруг проглоченной крови у москита (как и у некоторых других кровососущих насекомых, например, комаров) образуется так называемая перитрофическая матрица.Промастиготы лейшманий размножаются в пищеварительном канале самок москитов. Приблизительно через неделю инфекция распространяется до верхних отделов пищеварительного канала и паразиты блокируют просвет канала своими телами и секретируемым им гелем. Когда самка кусает потенциального хозяина, она выделяет в кожу свою слюну. Самка с блокированным пищеварительным каналом не может глотать, и у неё возникают спастические движения, в результате которых она отрыгивает промастиготы в ранку на коже хозяина.В среднем во время укуса инфицированным москитом в кожу попадает 102 — 103 промастигот.Первыми на место повреждения прибывают полиморфоядерные нейтрофилы, которые фагоциритуют паразитов. Внутри нейтрофилов лейшмании не размножаются и не превращаются в амастиготы. Затем, когда нейтрофилы переходят в фазу апоптоза, они уничтожаются макрофагами, и лейшмании проникают в макрофаги, не вызывая иммунного ответа.[15] Макрофаги являются основными клетками-хозяевами лейшманий в организме млекопитающих. Внутри макрофагов лейшмании трансформируются во внутриклеточную морфологическую форму — амастиготы.Внутри макрофага лейшмании заключены в так называемые "паразитифорные вакуоли", которые образуются от слияния первичной фагосомы c лизосомами и эндосомами. В них промастиготы транформируются в амастиготы. При этом происходят изменения в морфологии — продолговатые промастиготы с длинным жгутиком превращаются в овальные амастиготы с коротким жгутиком, в метаболизме, чтобы приспособиться к кислой среде, и в биохимическом составе мембраны. Трансформация занимает от двух до пяти дней. Амастиготы способны выживать в кислой среде этих вакуолей и питаться их содержимым. Внутри вакуоли амастиготы медленно размножаются, и каждый цикл размножения занимает около 24 часов.При кожном лейшманиозе инфильтрат образуется в коже и содержит преимущественно макрофаги, а также лимфоидные клетки и немного плазматических клеток. При висцеральном лейшманиозе очаги инфекции образуются в органах ретикуло-эндотелиальной системы.[16]Для обнаружения возбудителя при кожном лейшманиозе готовят мазки из содержимого бугорков, при висцеральном лейшманиозе — из пунктата костного мозга; кроме того, делают посев крови. Профилактика. Уничтожение грызунов в радиусе до 1,5 км от жилья, ликвидация мест выплода москитов. Защита населения от нападения москитов (сетки, пологи,репелленты). Уничтожение собак, заболевших лейшманиозом.Выявление, госпитализация и лечение больных.В очагах кожного лейшманиоза целесообразна вакцинация вновь прибывших лиц живой культурой лейшманий. 107. Виды малярийных плазмодиев и вызываемые ими заболевания. Систематическое положение, циклы развития. Борьба с малярией. Малярийные плазмодии (Тип Простейшие, Класс Споровики) —возбудители малярии. Известны следующие виды малярийных плазмодиев, паразитирующие у человека: возбудитель трехдневной малярии, возбудитель тропической малярии, возбудитель четырехдневной малярии, возбудитель овале-малярии, близкой к трехдневной. Жизненный цикл малярийных плазмодиев типичен для споровиков, включая стадии бесполого размножения в виде шизогонии, полового процесса и спорогонии. Окончательным хозяином паразитов является комар, а промежуточным — только человек. Комар является одновременно и переносчиком. Поэтому малярия — типичное антропонозное трансмиссивное заболевание.Со слюной зараженного комара при укусе плазмодии попадают в кровь человека. С током крови они разносятся по организму и поселяются в клетках печени. Здесь они растут и размножаются. Клетки печени при этом разрушаются и паразиты, называющиеся на этой стадии мерозоитами, поступают в кровь и внедряются в эритроциты. С этого момента начинается эритроцитарная часть цикла развития плазмодия. Паразит питается гемоглобином, растет и размножается шизогонией. Следующая стадия называется амебовидным шизонтом. У паразита появляются ложноножки, а вакуоль увеличивается. Наконец плазмодий занимает почти весь эритроцит. Следующая стадия развития паразита — фрагментация шизонта. После разрушения эритроцита мерозоиты попадают в плазму крови и оттуда в новые эритроциты, после чего весь цикл эритроцитарной шизогонии повторяется.Таким образом, в организме человека плазмодий размножается только бесполым путем — шизогонией, человек является его промежуточным хозяином. В организме комара проходят две другие стадии цикла развития паразита: половой процесс — гаметогония и образование спорозоитов за счет деления под оболочкой ооцисты — спорогония. Поэтому малярийный комар является окончательным хозяином этого паразита.При четырехдневной малярии приступы повторяются через 72 ч. Часто встречается и бессимптомное носительство. При тропической малярии вначале приступы развиваются через разные промежутки времени, а позже — через 24 ч. От осложнений со стороны центральной нервной системы или почек возможна смерть больного. Шизонты в клетках печени не сохраняются, а заболевание может продолжаться до 18 мес.Лабораторный диагноз малярии можно поставить только в период, соответствующий стадии эритроцитарной шизогонии, когда в крови удается обнаружить паразитов.Профилактика малярии — раннее выявление и лечение больных, профилактическое лечение в зонах широкого распространения малярии. Как и при любых трансмиссивных заболеваниях, необходима прицельная борьба с переносчиками (осушение болот, опрыскивание химическими веществами место размножения переносчиков, средства индивидуальной защиты).
108. Токсоплазма. Систематическое положение, морфология, цикл развития, пути заражения, диагностика, профилактикаТоксоплазма — возбудитель токсоплазмоза. Имеет форму полумесяца, один конец которого заострен более другого. В центре располагается крупное ядро. Дина паразита 4-7 мкм. Жизненный цикл токсоплазмы типичен для споровиков: в нем чередуются стадии шизогонии, гаметогонии и спорогонии. Основные хозяева паразита — домашние кошки и дикие виды семейства Кошачьи. Они заражаются, поедая больных грызунов, птиц или инвазированное мясо крупных животных. Паразиты у них сосредотачиваются в клетках кишечника, размножаются шизогонией, а затем образуют гаметы. После копуляции гамет формируются ооцисты, которые выделяются во внешнюю среду. В них происходит спорогония, т.е. деление зиготы под оболочкой.Своеобразной особенностью цикла развития токсоплазм является то, что промежуточные хозяева могут заражаться ими не только от основного хозяина, но и при поедании друг друга. Возможно и внутриутробное заражение плода от больной беременной самки, когда паразиты проникают через плаценту.В соответствии с этим и человек как промежуточный хозяин может заразиться токсоплазмозом разными путями: 1) при поедании мяса инвазированных животных; 2) с молоком и молочными продуктами; 3) через кожу и слизистые оболочки; 4) внутриутробно через плаценту; 5) при медицинских манипуляциях переливания крови и лейкоцитарной массы, при пересадках органов, сопровождающихся приемом иммунодепрессивных препаратов. Наиболее опасным является трансплацентарное заражение. При этом возможно рождение детей с множественными врожденными пороками развития, в первую очередь головного мозга. Для исследования используют плаценту, печень, кровь, лимфатические узлы, головной мозг.Профилактика — термическая обработка животных продуктов питания, санитарный контроль на бойнях и мясокомбинатах, предотвращение тесных контактов детей и беременных женщин с домашними животными.
109. Балантидий. Систематическое положение, морфология, цикл развития; пути заражения, диагностика, профилактика.Балантидии (Balantidium coli), простейшее из отряда равноресничных инфузорий. Паразитирует в организме человека и свиней. Б. впервые был обнаружен у свиней в 1862 немецким протозоологами Р. Лейкартом и Ф. Штейном. Несимметричное овальное тело Б. (длиной 30 — 150 мкм, шириной 20—110 мкм) покрыто короткими ресничками, расположенными продольными рядами. Размножается Б. поперечным делением. Паразитирует в толстых кишках, преимущественно в слепой кишке. Питается крахмалом и остатками пищи в каловых массах, форменными элементами крови и др. Иногда Б. пожирают один другого. Б. в кишечнике могут округляться и одеваться плотной оболочкой, образуя цисты, которые выходят наружу с фекалиями хозяина. Б. могут быть причиной заболевания — балантадиаза.Жизненный циклБалантидий обитают в кишечнике свиней, для которых малопатогенны. С испражнениями свиней цисты паразита выделяются в окружающую среду, где могут сохраняться несколько недель. Попадая с загрязненной водой или пищей в рот цисты в толстом кишечнике человека дают начало вегетативной стадии с последующим их размножением. Человек, больной или носитель, только в редких случаях может быть источником распространения балантидиев, так как у человека цисты образуются редко и в незначительном количестве, а вегетативными стадиями заразиться почти невозможно. Клиническая картина. Балантидий могут внедряться в слизистую оболочку толстого кишечника и вызывать воспалительно-язвенный процесс. В результате развивается балантидиаз. У больного наблюдаются понос, боли в животе, интоксикация, рвота, головные боли, в испражнениях — слизь, кровь. Болезнь может протекать в субклиничеекой, острой или хронической формах, в ряде случаев приводя к летальному исходу.ДиагнозДля обнаружения балантидиев каплю свежевыде-ленных испражнений помещают в изотонический раствор хлорида натрия на предметном стекле и исследуют под малым увеличением микроскопа (см. 9.4). Балантидий хорошо видны благодаря крупным размерам и активному движению. Выделяются они периодически, поэтому исследование при отрицательном результате необходимо повторять несколько раз. В некоторых случаях назначают солевое слабительное. У носителей обнаруживают только единичное цисты. Профилактика. Соблюдение правил личной гигиены, особенно при уходе за свиньями. Охрана от загрязнения воды и пиши. Балантидиаз чаще регистрируется в южных районах, хотя спорадически он выявляется повсеместно, особенно там, где развито свиноводство.
110. Тип Плоские черви. Основы систематики, характерные черты организации, медицинское значение.Тип плоские черви включает три основных класса: ресничные (молочная планария), сосальщики (печеночный сосальщик) и класс ленточные черви (бычий цепень). Два последних класса — паразитические формы.1. Трехслойные многоклеточные животные.2. Двусторонняя симметрия тела.3. Тело их вытянуто в длину и сплющено в спинно-брюшном направлении.4. Передвижение с помощью сокращений кожно-мускульного мешка. 5. Не имеют полости тела.6. Пищеварительный канал состоит из эктодермальной передней кишки, или глотки и эндодермальной средней, замкнутой слепо. Анального отверстия нет. У паразитов пищеварительная система полностью редуцирована.7. Нервная система состоит из парного мозгового ганглия и идущих от него в заднюю часть тела нервных стволов, соединенных кольцевыми перемычками.8. Кровеносная и дыхательная системы отсутствуют.9. специальные органы выделения.10. Половая система гомафродитна и очень сложна.11. Обычно имеются личиночные стадии.
111. Класс Сосальщики. Систематическое положение, характерные черты организации, медицинское значение.Печеночный сосальщик. Сосальщики - это паразитические черви с плоским листовидным или удлиненным нерасчлененным телом. Оно покрыто плотной защитной оболочкой; ресничного эпителия нет. К телу хозяина черви прикрепляются с помощью присосок. Тип плоские черви, класс сосальщики.Печеночный сосальщик - листовидное тело от 3 до 5 см с двумя присосками - ротовой и брюшной. На дне ротовой присоски находится рот, ведущий в глотку, от которой отходят 2 ветви средней кишки с многочисленными боковыми выростами. В жизненном цикле наблюдается смена хозяев. Взрослая гермафродитная особь живет в печени окончательного (основного) хозяина (овцы, крупного рогатого скота, свиньи, лошади, изредка человека). Питается в основном желчью, вызывая истощение или даже гибель хозяина. Оплодотворенные яйца паразита вместе с желчью поступает в кишечник хозяина, выводятся во внешнюю среду. В пресном водоеме, где через 15-25 дней из яйца выходит личинка, с помощью которых она плавает. В теле промежуточного хозяина - моллюска малого прудовика личинка внедряется с помощью буравящего стилета, проникает в печень. Образуется спороциста, в ней образуется личинка. Личинка в теле промежуточного хозяина дважды размножается партеногенетически. Покинув тело моллюска, личинка свободно плавает, прикрепляется к растениям, теряет хвост, одевается плотной оболочкой и превращается в цисту, внутри которой сохраняет свою жизненную способность. С зеленым кормом цисты попадают в организм домашних животных, первоначально в желудок, оболочка цисты растворяется и паразит проникает в кишечник, из него в желчные протоки печени, вырастает во взрослого сосальщика. Человек заражается ими при употреблении сырой воды из водоема, овощей и фруктов, вымытых в этой воде.Ряд приспособлений: форма тела, размеры, покров (покрыт плотной оболочкой, выполняющей защитную роль от действия пищеварительных соков хозяина); присоски (ими червь прикрепляется к печени хозяина), продукты жизнедеятельности (ядовитые жидкие вещества) выделяются в организм хозяина, вызывая страшные страдания животных и человека от заболевания; наличие в жизненном цикле паразита кроме размножающегося половым путем гермафродитного поколения еще и партеногенетического размножения (без оплодотворения) у личинок можно рассматривать как приспособительный признак, обеспечивающий резкое повышение интенсивного размера. Профилактика - уничтожение в местных водоемах малого прудовика (промежуточного хозяина) химическими средствами, лечебными препаратами - с взрослой формой этого паразита.Диагностика — обнаружение яиц в фекалиях, моче или мокроте, аллергические пробы.
112. Описторхис, клонорхис, дикроцелиум, нанофиетус, метагонимус, парагонимус, шистосомы. Систематическое положение, морфология, циклы развития, диагностика, пути заражения, профилактика, географическое распространение.Описторхис[источник не указан 611 дней] (лат. Opisthorchis) — род дигенетических сосальщиков из отряда Opisthorchida. В роли окончательного хозяина представителей выступают хищные млекопитающие и человек, в которых черви проникают через пищу. Вызываемое представителями этого рода заболевание — описторхоз. Основную роль в распространении описторхоза играют люди и кошки, употребляющие зараженную рыбу. Дети болеют реже взрослых, поскольку употребляют в пищу меньше рыбы, не прошедшей термическую обработку. В механизме передачи описторхоза наибольшее эпидемиологическое значение имеет загрязнение водоемов фекалиями. Методы диагностики: Микроскопия. Выявление яиц описторхиса в дуоденальном содержимом и в фекалиях.Анализ крови. Кровь исследуется на количество эозинофилов и признаки анемии.Клонорхоз — гельминтоз, возбудитель которого паразитирует в желчных протоках, желчном пузыре и поджелудочной железе. В России основные очаги клонорхоза регистрируются в Приамурье и Приморье. Клонорхоз широко распространен в Китае, Японии, Корее и Вьетнаме. Возбудителем болезни является двуустка китайская. Развитие паразита происходит со сменой трех хозяев. Окончательным хозяином гельминта являются человек, собаки и другие млекопитающие. Промежуточный хозяин — моллюск, дополнительный — карповые рыбы. Источником инвазии являются больные люди и животные, выделяющие в огромном количестве яйца паразита. Заражение человека происходит при употреблении в пищу недостаточно прожаренной, вареной или сырой, пораженной клонорхозом рыбы. Возбудитель дикроцелиоза - гельминт-трематода ланцетовиднаяDicrocoelium lanceatum - двуустка ланцетовидная. Длина паразита 5-15 мм, ширина 1-2, 5 мм. Яйца темно-коричневые, имеют толстую оболочку; размер яиц 0, 038- 0, 045 х 0, 022-0, 030 мм. Паразитирует у овец, коз, крупного рогатого скота, буйволов, верблюдов, лошадей, ослов, мулов, маралов, зайцев, кроликов, сусликов и человека. Возбудитель откладывает яйца с уже развитым мирацидием, который вылупляется после того, как яйцо будет заглочено наземными моллюсками. В организме моллюска происходит развитие спороцист, после чего сформировавшиеся церкарии выделяются в окружающую среду. Затем они попадают в организм муравьев, где развиваются метацеркарии. В эпизоотологии дикроцелиоза большое значение имеют условия активности моллюсков. Активность максимально проявляется при 67 - 100%-ной влажности и температуре воздуха 3,5 18 'С. Поэтому церкарии выходят из моллюсков чаще после дождей. Яйца дикроцелий устойчивы во внешней среде. Высушивание при температуре 18 - 20 'С не убивает их в течение недели. Они переносят также сильный холод (до - 23, а иногда до -50 'С). Таким образом, первый промежуточный хозяин паразита наземные моллюски, второй - муравьи, окончательные хозяева - сельскохозяйственные и дикие животные, а также человек. Диагноз подтверждает обнаружение в дуоденальном содержимом и фекалиях яиц гельминта. Профилактика заключается в предупреждении заползания на продукты питания муравьев, особенно в экспедициях, туристских походах; в выявлении и лечении домашних животных, больных дикроцелиозом.Нанофиетоз — гельминтоз с преимущественным поражением кишечника. В России встречается в Приморском крае (бассейны рек Амура и Уссури, на Сахалине), большая зараженность отмечается в удэгейских селах. Заболевание регистрируется также в Северной Америке. Окончательными хозяевами паразита являются человек, собака, норка, кошка, барсук, волк, енот, медведь. Промежуточным хозяином служит моллюск, дополнительным — рыба (кета, кунджа, амурский сиг, таймень, хариус). Человек заражается при употреблении в пищу сырой, плохо проваренной или недожаренной рыбы. Местом локализации паразита у человека является тонкий кишечник. Клинические проявления болезни могут практически отсутствовать при слабой инвазии или быть незначительными. Чаще всего у больных отмечаются боли в животе, неустойчивый стул (чередование запоров и поносов), тошнота, возможна рвота и другие признаки энтерита. Лабораторная диагностика основана на обнаружении яиц в кале больных (не ранее 5—7-го дня после заражения). Метагонимоз — гельминтоз, протекающий преимущественно с кишечными расстройствами.Этиология. Возбудителем является трематода Metagonimus yokogawi (Yokogawa, 1911) длиной 1-2,5 мм, шириной 0,4-0,7 мм. Тело покрыто маленькими шипиками, брюшная присоска слита с ротовой.Яйца имеют размеры 0,023—0,029 х 0,014—0,018 мм с крышечкой на одном и утолщением скорлупы на другом конце. Взрослые паразиты локализуются в тонком кишечнике человека, собаки, кошки. Развитие происходит со сменой двух хозяев — пресноводных моллюсков и различных видов рыб (форель, уссурийский сиг, амурский лещ, сазан, карась, верхогляд и др.).Эпидемиология. Источником инвазии являются больные люди, собаки, кошки, выделяющие с фекалиями яйца гельминта. Выходящая из яиц личинка активно проникает в моллюска, где размножается и развивается в церкариев. Церкарии выходят в воду и проникают в рыб, инцистируются и превращаются в метацеркарии. Заражение человека происходит при употреблении в пищу сырой рыбы и случайном заглатывании ее чешуек. Заболевание распространено в Китае, Корее, Японии, в бассейне Амура.Патогенез. Из проникших в кишечник человека цист (метацеркарии) вылупляются личинки, внедряющиеся в слизистую оболочку тонкой кишки. Через 2 недели они достигают половой зрелости и выходят в просвет кишечника. В основе проявлений заболевания лежат механическое и токсико-аллергическое воздействие паразита на слизистую оболочку кишечника, раздражение энтерорецепторов.Симптомы и течение. На 2-й неделе после заражения возникают лихорадка, головная боль, тошнота, диарея, уртикарная и папулезная экзантема, эозинофильный лейкоцитоз. Острые проявления сохраняются в течение 2—4 суток. В поздних стадиях метагонимоз протекает по типу хронического рецидивирующего энтерита, проявляющегося тошнотой, слюнотечением, болью в животе без определенной локализации, упорной диареей. Редко заболевание протекает субклинически.Диагноз и дифференциальный диагноз. Чаще всего приходится дифференцировать метагонимоз с диареями бактериальной и паразитарной природы, новообразованиями кишечника. Решающее значение имеет обнаружение в кале яиц метагонимуса, сходных с яйцами возбудителей описторхозаи клонорхозаПарагонимоз (paragonimoses) — гельминтоз из группы трематодозов, характеризующийся преимущественным поражением легких, подкожной клетчатки и скелетных мышц, реже — головного мозга. Возбудитель лёгочного парагонимоза — Paragonimus westermani (Paragonimus ringeri) — Лёгочный сосальщик. Тело Paragonimus westermani яйцевидной формы, красно-коричневого цвета, длиной 7,5-13 мм, шириной 4-8 мм, толщиной 3,5-5 мм. Поверхность тела покрыта шипиками. Брюшная присоска около середины тела. Яйца золотисто-коричневые, овальные, с крышечкой. Размеры яиц - 63 - 84 x 45 - 54 мкм. Яйца имеют золотисто-коричневый оттенок. Симптомы: лихорадка, одышка, кашель с гнойной мокротой (иногда с примесью крови). Через 2-3 мес проявления переходят в хроническую стадию; отмечают периоды обострений и ремиссий. У некоторых больных развивается серозный экссудативный плеврит. При заносе гельминтов в головной мозг развиваются симптомы поражения ЦНС.Диагноз легочного парагонимоза устанавливают на основании клинической картины, обнаружения яиц гельминта в мокроте, иногда и в фекалиях. Диагноз личиночного парогонимоза подтверждается серологическими реакциями со специфическим антигеном. При заражении P. szechuanensis диагноз устанавливают на основании результатов биопсии узла. В биоптате обнаруживают личинки паразита.Шистосомоз - паразитарное заболевание, вызванное плоскими червями шистосомами и считающееся присущим преимущественно тропическим странам. Последние, попадая в кровь человека, мигрируют в вены брюшной полости, после чего сквозь сосуды их яйца проникают в кишечник и мочевой пузырь, которые впоследствии разрушаются за счёт сильнейшего яда, который выделяется шистосомами. Путь заражения: Купание в стоячих водоёмах, где могут быть продукты жизнедеятельности заражённого шистосомозом человека или животного. Диагностика: анализ кала и мочи, эмпирический осмотр печени.
113. Класс Ленточные черви. Систематическое положение, характерные черты организации, медицинское значение.Ленточные черви (лентецы, цестоды) (лат. Eucestoda) — класс паразитических плоских червей (Platyhelmithes). Описано около 3500 видов. Представители этого таксона полностью утратили пищеварительную систему. Некоторые виды — опасные паразиты человека; вызываемые ими заболевания носят название цестодозы.В традиционной трактовке жизненный цикл цестод представляет собой онтогенез одной особи, сменяющей нескольких хозяев. Дефинитивные (способные к половому размножению) стадии паразитируют в кишечнике водных и наземных позвоночных. Стадии, обитающие в промежуточных хозяевах (позвоночных и беспозвоночных), могут локализоваться в тканях и полостях тела.Жизненный циклЖизненный цикл может состоять из 3—4 этапов. На первом этапе взрослые черви обитают в кишечнике окончательного хозяина, размножаются и продуцируют яйца. На втором этапе яйца попадают во внешнюю среду: в почву или в воду. На суше в яйцах формируется личинка или зародыш, представляющий фазу внедрения в промежуточного хозяина. У некоторых видов, яйца которых развиваются в воде, из яйца выходит свободноплавающая личинка, покрытая ресничками, а в ней формируется вторая личиночная фаза. На третьем этапе происходит развитие личинок в промежуточном хозяине, где развиваются в пузырчатую глисту — финну. Финна для дальнейшего развития должна попасть в кишечник основного хозяина, где головка из финны выворачивается, присасывается к стенке кишечника, после чего начинается процесс роста червя.Свиной солитер (Taenia solium) в коллекции Зоологического музея ЗИН РАН[править]СтроениеВ связи с паразитизмом у большинства ленточных червей редуцирована пищеварительная система, слабо развиты нервная система и органы чувств. С другой стороны, у них сильно развита половая система, метамерно повторяющаяся в члениках, что обеспечивает их высокую плодовитость как паразитов. Это повышает возможность выживания ленточных червей, развивающихся со сменой хозяев.Данный класс характеризуется длинным лентовидным телом (от долей милимметра до 10 метров), обычно подразделённым на членики (числом от двух до нескольких тысяч), а на переднем конце тела имеется «головка» — сколекс с органами прикрепления (присосками, иногда дополеннными хитиновыми крючками). Но имеются черви и с нерасчленённым телом.За головкой расположена шейка, являющаяся зоной роста ленточного червя. От шейки постоянно отшнуровываются новые членики. На заднем конце тела зрелые членики с яйцами отрываются и выносятся с фекалиями хозяина во внешнюю среду, а за счёт образования новых члеников в области шейки тело паразита восстанавливается.Тело червей покрыто тегументом, состоящим из наружного цитоплазматического слоя клеток, которые имеют вытянутую форму, благодаря которой ядра находятся в погружённом слое. Поверхность тегумента образует микротрихии, имеющие значение при всасывании пищи из кишечника хозяина. Наружный слой тегумента подстилает мембрана, под которой располагаются кольцевые и продольные мышцы. Как и у других плоских червей, у ленточных имеются пучки дорсовентральных мышц.Нервная система ортогональна. В сколексе имеется парный ганглий, от которого отходит несколько пар нервных тяжей. Наиболее развиты два боковых ствола. В коже червей располагаются осязательные и рецепторные клетки.Половая система червей гермафродитная и повторяется в каждом членике. У видов с нерасчлененным телом может быть одиночный половой аппарат или метамерный ряд половых аппаратов. У цестод небольших размеров оплодотворение перекрёстное. Крупные формы (солитеры) встречаются в кишечнике хозяина поодиночке, что делает перекрёстное оплодотворение невозможным. При половом размножении у них происходит копуляция разных члеников одной особи. Самооплодотворение в пределах одного членика очень редко. Плодовитость цестод чрезвычайно велика, например, солитёр в год продуцирует около 600 млн яиц, а за всю жизнь (18—20 лет) он может производить около 11 млрд яиц.
114. Свиной и бычий цепни. Систематическое положение, особенности морфологии, жизненные циклы, пути заражения, диагностика, профилактика.Бычий цепень (Тип Плоские черви; Класс Ленточные черви; ленточные черви, жизненный цикл развития которых не связан с водной средой; ленточные черви, жизненный цикл которых не связан с водной средой). Бычий цепень (Taeniarrhyncnus saginatus) - возбудитель тениаринхоза, достигает в длину 4—10 м. На головке имеет только четыре присоски. Гермафродитные членики квадратной формы, матка в них не разветвляется, а яичник состоит из двух долей. Зрелые членики сильно вытянуты. Матка очень разветвлена, число ее боковых ветвей достигает 17—34 пар. Яйца содержат онкосферы, расположенные под тонкой прозрачной оболочкой, которая быстро разрушается. Онкосферы имеют три пары крючьев и толстую, радиально исчерченную оболочку. Диаметр онкосфер около 10 мкм. Тениаринхоз распространен повсеместно, где население употребляет в пищу сырое или недостаточно обработанное говяжье мясо.Жизненный цикл бычьего цепня типичен. Основной хозяин только человек, промежуточный — крупный рогатый скот. Характерной особенностью является способность члеников активно выползать из заднепроходного отверстия поодиночке. Корова, проглотив такие членики, становится промежуточным хозяином паразита. В ее мышцах формируются финны, называемые цистицер-ками. Финна представляет собой пузырек, заполненный жидкостью, в котором находится сколекс. В мышцах финны могут сохранять жизнеспособность долгие годы. При поедании мяса такой коровы в желудке под действием кислой среды желудочного сока головка вывертывается, прикрепляется к стенке кишки и развивается новый цепень.Диагностика проводится несложно — при обнаружении зрелых члеников в фекалиях, так как членики имеют характерное строение.Свино́й це́пень, или свино́й солитёр, или вооружённый це́пень (лат. Taenia solium) — вид ленточных червей из отряда Cyclophyllidea. Паразит млекопитающих. В качестве промежуточных хозяев использует свиней, собак, зайцев, кроликов, верблюдов и человека. Окончательный хозяин — только человек. В зависимости от стадии жизненного цикла, на которой происходит заражение, свиной цепень вызывает у хозяина тениоз (заражение на стадии личинки) или цистицеркоз (заражение на стадии яйца). В отличие от бычьего невооружённого цепня, у свиного на сколексе имеется в дополнение к четырём присоскам двойной венчик крючьев, служащий для лучшего закрепления червя в кишечнике хозяина[1]. Стробила меньшего размера, чем у бычьего цепня, достигает в длину 2—3 м[2]. Зрелые членики отделяются по 5—6 штук и выходят с испражнениями, не проявляя активных движений[3]. Промежуточный хозяин свиного цепня — домашние или дикие свиньи, также им может быть и человек.
115. Цистицеркоз. Пути заражения. Профилактика.Цистицеркоз (лат. Cysticercosis) — гельминтоз из группы цестодозов, характеризующийся в зависимости от локализации паразита поражением кожи, подкожной клетчатки, мышц, головного и спинного мозга, глаз, внутренних органов, костей. Возбудитель — личинки свиного цепня (Taenia solium).Цистицеркоз развивается в результате попадания в желудок яиц свиного цепня с загрязненными продуктами, через грязные руки, при забрасывании зрелых члеников из кишечника в желудок при рвоте у лиц, зараженных половозрелой формой свиного цепня (то есть как осложнение тениоза).Цистицерк представляет собой наполненный прозрачной жидкостью пузырь размером от горошины до грецкого ореха (от 3 до 15 мм в диаметре). На внутренней поверхности пузыря располагается головка финны — сколекс с крючьями и присосками.Для развития цистицеркоза необходимо попадание яиц гельминта в желудок и кишечник, где под влиянием желудочного сока плотная оболочка яиц растворяется и освободившиеся зародыши по кровеносным сосудам разносятся по тканям и органам человека.Источником возбудителя инвазии является больной человек, выделяющий с фекалиями зрелые яйца и членики цепня. Заражение происходит через загрязненные руки, пищу, воду; помимо заражения яйцами цепня из окружающей среды больной тениозом человек может заразиться цистицеркозом при антиперистальтике, рвоте с последующим заглатыванием онкосфер (аутоинвазия).При проглатывании онкосферы в тонкой кишке личинка освобождается от оболочки, головка личинки выворачивается наружу, и личинка активно внедряется в стенку кишки, попадает в кровеносные сосуды и током крови разносится по организму. Помимо токсического и сенсибилизирующего действия ферментов и метаболитов личинок при миграции в дальнейшем важным фактором патогенеза является механическое давление растущего паразита. Токсико-аллергическое воздействие паразита, например при гибели и лизисе паразита, может вызватьанафилактический шок.Диагноз устанавливают на основании обнаружения округлых образований в коже и мягких тканях туловища, конечностей. На рентгенограмме выявляются овальные или веретенообразные образования с четко контурированной оболочкой, нередко кальцифицированные. Распознаванию болезни способствуют данные эпидемиологического анамнеза, наличие у больного тениоза. Диагноз подтверждают положительные результаты серологических реакции (связывание комплемента, непрямая гемагглютинация, иммуноферментный анализ) с антигенами цистицерка, положительная реакция с ними цереброспинальной жидкости, обнаружение в ликворе цитоза с высоким процентом лимфоцитов и умеренной эозинофилией.
116. Карликовый цепень. Систематическое положение, особенности морфология жизненный цикл, пути заражения, диагностика, профилактика.Карликовый цепень (лат. Hymenolepis nana) — вид ленточных червей отряда циклофиллид (Cyclophyllidea). Половозрелые особи паразитируют в кишечнике человека, намного реже у мышей и крыс. Является одним из немногих представителей ленточных червей, которые могут осуществлять все стадии жизненного цикла, не выходя из окончательного хозяина. Биология карликового цепня довольно своеобразна, ибо развитие гельминта происходит в организме одного хозяина, который сначала служит для паразита промежуточным хозяином, а затем становится окончательным. Однако не исключается возможность развития карликового цепня и с участием промежуточного хозяина в качестве некоторых насекомых - блох и мучных жуков. Основным хозяином карликового цепня является человек. Человек заражается карликовым цепнем при заглатывании яиц Н. nаnа, которые проходят желудок и попадают в верхний отдел тонкой кишки Здесь онкосфера активными движениями эмбриональных крючьев самостоятельно освобождается от яйца и внедряется в ворсинку или в толщу солитарного фолликула нижней трети тонкого кишечника, где осуществляется тканевая фаза инвазии, когда человек является промежуточным хозяином. Онкосфера через 6-8 суток превращается в цистицеркоид ( зародыш, имеющий вздутую переднюю часть с ввернутой головкой, на заднец конце тела - хвостовидный придаток с крючьями). Цистицеркоиды могут развиваться также в лимфоидных фолликулах, в подслизистом слое, т. е. там, куда проникают онкосферы, а также в других органах: печени, брыжеечных лимфоузлах. Через 5-8 суток в результате разрушения ворсинок цистицеркоиды выпадают в просвет кишки. Нельзя исключить также возможного активного освобождения цистицеркоидов карликового цепня из ворсинок. Этим заканчивается тканевая фаза развития и начинается кишечная, когда человек является окончательным для карликового цепня хозяином. Цистицеркоид, имея готовый сколекс, с помощью присосок и крючьев фиксируется к слизистой оболочке тонкой кишки. Начинается процесс стробиляции, который длится в среднем до 2 недель. Таким образом, формирование взрослой особи карликового цепня от момента заражения завершается за 3 недели. Диагностика карликового цепня основана на обнаружении яиц в фекалиях.
117. Эхинококк и альвеококк. Систематическое положение, особенности морфологии, жизненные циклы диагностика, профилактика, особенности географического распространения.Эхинококк — возбудитель эхинококкоза. Половозрелая форма имеет головку с крючьями и 3-4 членика разной степени зрелости. Последний из них зрелый, он содержит около 800яиц. Общая длина тела до 5 мм. Яйца по форме и размерам сходны с яйцами свиного и бычьего цепней. Эхинококкоз у человека распространен во всех географических и климатических зонах, преимущественно в регионах с развитым отгонным животноводством. Жизненный цикл эхинококка связан с хищными животными семейства Псовых (волками, шакалами, собаками), которые являются его окончательными хозяевами. Взрослые членики способны активно ползать, распространяя яйца по шерсти хозяина и в окружающей среде. Их могут проглотить травоядные животные — коровы, овцы, олени или человек, становясь промежуточными хозяевами. Финна (промежуточная форма паразита) эхинококка — пузырь, нередко достигающий 20 см. в диаметре. Он заполнен жидкостью с ограниченным количеством молодых сколексов, постоянно почкующихся от внутренней поверхности стенки финны. Окончательный хозяин заражается, поедая пораженные органы промежуточного. Растущая финна сдавливает органы, вызывает их атрофию. Постоянное поступление продуктов диссимиляции в организм хозяина вызывает его истощения. Очень опасен разрыв эхинококкового пузыря: жидкость, заключенная в нем, может вызвать токсический шок. При этом мелкие зародышевые сколексы могут распространяться по организму, поражая другие органы. Множественный эхинококкоз заканчивается смертью хозяина. Личная профилактика заражения — мытье рук после контактов с пастушьими собаками. Общественная профилактика — обследование и дегельминтизация собак, недопущение скармливания им органов больных животных. Альвеококк — возбудитель альвеококкоза. Половозрелая форма отличается от эхинококка меньшими размерами тела (до 2 мм) и деталями строения крючьев и матки. Финнозная стадия состоит из множества мелких пузырьков, постоянно почкующихся друг от друга наружу, благодаря чему финна представляет собой постоянно растущий узел. В каждом пузырьке находится зародышевая головка паразита. Рост финны разрушает окружающие ткани наподобие злокачественной опухоли. Возможен и отрыв отдельных пузырьков с распространением патологического процесса по организму. Природные очаги альвеококкоза имеются в Сибири, Средней Азии, на Урале и Дальнем Востоке, а также в Северной Америке, в Центральной и Южной Европе.Жизненный цикл — принципиально не отличается от эхинококка, но окончательными хозяевами этого паразита являются дикие хищники — лисы, песцы, волки, реже домашние — собаки, промежуточными — мышевидные грызуны, иногда человек. Альвеококкоз — более тяжелое заболевание, чем эхинококкоз, в связи с инвазивным характером роста финны. Личная профилактика — как при эхинококкозе, общественная — соблюдение правил гигиены при обработки шкур промысловых животных, а также запрещение скармливания собакам тушек грызунов
118. Возбудители дифиллоботриоза - систематическое положение, особенности морфологии, жизненные циклы, пути заражения, диагностика, профилактика. Возбудитель дифиллоботриоза на ДВ.Дифиллоботриоз — гельминтоз, протекающий с признаками преимущественного поражения желудочно-кишечного тракта и часто сопровождающийся развитием мегалобластной анемии.Этиология. Возбудители дифиллоботриоза относятся к отряду Pseudophylideaи насчитывают 12 видов лентецов, из которых лентец широкий (Diphyllobothrium latum) является наиболее распространенным и изученным (Unney, 1758, Lube, 1910). Длина стробилы, состоящей из большого числа члеников (до 4000), достигает 2-9 м. Сколекс длиной 3—5 мм имеет продолговато-овальную форму, сплющен с боков, на боковых поверхностях — две щели (ботрии), посредством которых паразит прикрепляется к слизистой оболочке кишечника. Яйца лентеца широкоовальные, крупные (70 х 45 мкм), с двухконтурной оболочкой, имеют на одном полюсе крышечку, на другом —бугорок. Паразитируя в органах окончательного хозяина, лентецы выделяют незрелые яйца, развитие которых происходит в пресноводных водоемах.Распространение дифиллоботриоза связано с крупными пресноводными водоемами. Очаги его преобладают в Северной Европе, Восточном Средиземноморье, в районе Великих озер США, в Канаде и на Аляске. В России заболевание регистрируется преимущественно в Карелии, Красноярском крае, на Кольском полуострове. Заражение человека происходит при употреблении свежей, недостаточно просоленной икры и сырой рыбы. Окончательными хозяевами лентеца широкого являются человек, собаки, кошки, медведи, лисицы, свиньи. Промежуточные хозяева — пресноводные рачки (циклопы, диаптомусы), дополнительные — пресноводные рыбы. Продолжительность жизни лентеца широкого в организме человека может достигать 25 лет, в организме собаки 1,5-2 года, кошки — 3-4 недели. В диагностике информативен эпиданамнез (пребывание в эндемичной области, употребление сырой рыбы, недосоленной икры). Часто больные сообщают о выделении с калом частей гельминтов. Для дифиллоботриоза характерно выделение обрывков стробилы, что отличает его от инвазии цепнями (бычьим и свиным).
119. Тип круглые черви. Классификация. Характерные черты организации, медицинское значение.Круглые черви — первичнополостные животные. У них развивается первичная полость тела, заполненная жидкостью и выполняющая роль гидроскелета. В первичной полсти помещаются все внутренние органы. Пищеварительная система имеет сложное строение (задняя кишка и заднее отверстие) и выполняет моторную и железистую функции. Нервная система представлена в передней части нервным окологлоточным кольцом, от которого отходят нервные стволы, соединенные комиссурами. Отсутствует кольцевой мускулатуры и мезенхимной ткани, постоянство клеточного состава, отсутствие способности восстанавливать утраченные части тела. Более 25 тыс. видов (свободный и паразитический образ жизни). - брюхоресничные;- собственно круглые черви;- волосатиковые;- коловратки;- скребниАскариды - аскаридоз.
120. Аскарида, власоглав, острица, анкилостомиды, угрица кишечная, трихинелла. Систематическое положение, особенности морфологии, жизненные циклы, пути зараражения, диагностика, профилактика. Аскарида человеческая — возбудитель аскаридоза — крупный гельминт, самки которого достигают 40 см длины, а самцы — 20 см. Тип и класс круглые черви. Зрелые яйца овальны и бугристы, оболочка их толстая и многослойная. Цвет желтовато — коричневый, длина до 60 мкм.Паразитирует в кишечнике человека. Оплодотворенное яйцо начинает развиваться в матке червя. Попадая с фекалиями в окружающую среду, яйца при доступе кислорода и достаточно высокой температуре развиваются, и под оболочкой яйца образуется личинка. С загрязненной водой , овощами, фруктами яйца попадают в кишечник человека, где из них выходят личинки, которые внедряются в стенки кишечника и проникают в кровь, по крови в легкие. Пробуравливают стенки капилляров и альвеол, проходят в бронхи, трахею, ротовую полость и вторично заглатываются. В кишечнике образуется взрослая аскарида. Аскариды отравляют организм токсичными продуктами обмена, а также воздействуют механически: при большом количестве могут вызвать непроходимость кишечника.Диагностика — обнаружение яиц в фекалиях больного.Профилактика — личная гигиена и гигиена питания, выявление больных аскаридозомВласоглав — круглый червь, вызывающий заболевание трихоцефалёз. Своё название червь получил за оригинальную форму тела — нитевидную переднюю часть (через неё проходит только пищевод) и значительно более широкую заднюю, в которой находятся все остальные внутренние органы. Размеры взрослой особи составляют 35—50 мм. Эти беловатого цвета гельминты обитают в начальном отделе толстой кишки, причём паразитируют только у человека. Власоглавы глубоко внедряются своим утончённым передним концом тела в стенку кишки и питаются тканевой жидкостью и кровью. Самка власоглава выделяет в течение суток около 60 000 яиц, которые попадают в просвет кишки и с фекалиями выносятся в окружающую среду. Личиночное развитие паразита также идёт без промежуточного хозяина (то есть он является геогельминтом). Во влажной почве при температуре 25—30 °C в яйце развивается личинка, через три-четыре недели яйца становятся инвазионными и могут быть проглочены. Попав в толстую кишку человека, личинки выходят из яиц, продолжают своё развитие и через несколько недель становятся половозрелыми.Возбудитель энтеробиоза - острица - мелкая нематода белого цвета. Длина самца 2-3 мм, самки - 9-12 мм. Хвостовой конец самца спирально загнут сторону брюшной поверхности, а у самки шиповидно заострен (поэтому гельминт получил название "острица"). Яйца остриц продолговаты, несколько асимметричны, как правило они обнаруживаются в кожных складках вокруг анального отверстия, в фекалиях встречаются редко. Однако взрослые самки при пассивном выхождении могут попадать в фекалии. Половозрелые острицы паразитируют в нижних отделах тонкой и верхних отделах толстой кишки. Самки, наполненные яйцами в количестве 5000-15000 шт каждая, неспособны удерживаться на слизистой оболочке кишок, спускаются до прямой кишки и выползают из анального отверстия для откладывания яиц в перианальных складках. После отложения яиц самка погибает, длительность жизни ее не превышает одного месяца. При проникновении острицчервеобразный отросток, они могут стать причиной аппендицита. Острицы живут в прямой кишке ребенка, но выползают на кожу, чтобы откладывать яйца. Обычно они делают это ночью, что объясняет то, что зудит в основном ночью.Анкилостомиды (Ancylostomatidae), семейство круглых червей - кишечных паразитов человека и млекопитающих животных (диких и домашних). Распространены на всех материках, преимущественно в тропиках и субтропиках. У человека паразитируют анкилостома (Аnсуlostoma duodenale, длина тела 8-13 мм) . Из яиц паразита, выделяемых зараженными животными или человеком, во внешней среде (в почве) развиваются личинки. Заражение возможно двумя путями: через кожу (личинка внедряется активно) и через рот (личинка попадает с пищей). Заболевания, вызываемые паразитированием анкилостом, называются анкилостомозом,СТРОНГИЛОИДОЗ - это заболевание, вызываемое стронгилоидой, мелким нитевидным паразитом, длиной около 2 мм. Этого гельминта называют еще угрицей кишечной. Стронгилоиды распространены на территории Закавказья, Украины, средней полосы России, в Средней Азии. Возбудители стронгилоидоза паразитируют в организме человека одновременно с анкилостомами в верхних отделах тонкой кишки, в двенадцатиперстной кишке. Источником данного заболевания являются зараженные люди. Пути заражения стронгилоидозом :Если личинка возбудителя стронгилоидоза попадает в организм человека через рот с загрязненной пищей или водой, то миграции, как правило, не происходит, а сразу развивается взрослая форма. Однако этот способ проникновения личинок - пассивное попадание - встречается гораздо реже. Основной путь заражения стронгилоидозом - активное внедрение через кожу при ходьбе босиком, работе с землей.В цикле развития стронгилоидиды происходит смена свободноживущих (в почве) и паразитирующих (в организме человека) поколений. Жизненный цикл этого гельминта очень специфический - он может проходить в теле человека без выхода во внешнюю среду. Попадая в кишечник, самка возбудителя стронгилоидоза откладывает яйца, из них развиваются личинки, которые внедряются в кровеносные сосуды, сердце, легочные артерии, альвеолы, бронхи, трахею, глотку, откуда снова попадают в кишечник и завершают свое развитие, превращаясь в половозрелые формы.В кишечнике взрослые особи живут 5-6 лет. Трихинеллы (лат. Trichinella) — род паразитических круглых червей класса Enoplea. В роли хозяев выступают плотоядные млекопитающие, в том числе — человек. Первая личиночная стадия паразитирует в поперечно-полосатой мускулатуре, три последующих личиночных стадии и взрослые особи — в просвете тонкого кишечника. Трихинеллы вызывают смертельно опасное заболевание — трихинеллёз. Копуляция раздельнополых червей происходит в просвете тонкого кишечника окончательного хозяина. Эмбриональное развитие и вылупление личинок из яйца происходит в половых путях самки (яйцеживорождение). Самки трихинелл внедряют передний конец тела в кишечный эпителий и рождают 1-2 тысячи личинок, которые разносятся через кровеносные и лимфатические сосуды по всему телу хозяина. Выживают лишь те личинки, которые попадали в поперечно-полосатые мышцы с хорошим кровоснабжением (жевательные, глазодвигательные мышцы, мышцы диафрагмы). Там они функционирующим лишь на этой стадии стилетом разрушают мышечную ткань и вызывают формирование хозяином капсулы веретеновидной формы. Впоследствии капсула пропитывается известью, однако обмен веществом между паразитом и хозяином не прекращается. Эта стадия (мышечная трихина) может существовать несколько лет. Заражённость мышц личинками трихинелл может достигать 15 тысяч на килограмм ткани.Для замыкания жизненного цикла необходимо, чтобы мышцы хозяина съело другое млекопитающее. При попадании в тонкий кишечник в течение нескольких дней трихинелла претерпевает четыре линьки, достигая половой зрелости. Таким образом, для развития одного поколения необходима смена хозяина, который последовательно выступает в роли окончательного для родительских форм и промежуточного — для дочерних.
121. Ришта. Систематическое положение, морфология, цикл развития, пути заражения, диагностика, профилактика. Работы Л.М. Исаева по ликвидации дракункулеза. Дракункулёз, или ришта (от тадж. нить) — гельминтоз из группы нематодозов, вызываемый самками круглых червей Dracunculus medinensis (англ.)русск.. Распространено в тропиках и субтропиках Африки и Азии. Паразит попадает в организм человека перорально: при заглатывании воды, содержащей веслоногих раков, заражённых личинками ришты. При попадании в кишечник ришта пробуравливает его стенку и попадает влимфатические сосуды, а оттуда проникает в полость тела, где претерпевает две последовательные линьки и достигает половой зрелости. После спаривания самцы гибнут, а самки мигрируют в кожу, где локализуются в подкожной клетчатке. Там самки продолжают расти и достигают длины 80 см. При контакте заражённого участка кожи с водой самка высовывает наружу передний конец тела и выбрасывает в воду многочисленных личинок, которые для замыкания цикла должны заразить веслоногого рака.Л. М. Исаев (1934) полагал, что первичным очагом дракункулеза была Африка, откуда это заболевание распространилось и в другие страны. В дальнейшем Л. М. Исаев (1923 — 1932) уточнил детали биологии и эпидемиологии ришты и разработал комплекс мероприятий по ликвидации этого гельминтоза. В результате целенаправленных оздоровительных мероприятий, проводимых под руководством и при непосредственном участии Л. М. Исаева, самый крупный очаг ришты (г. Бухара и ее окрестности) был ликвидирован в 1932 г. Очаги в других городах прекратили свое существование еще ранее в результате изменившихся социальных и природных условий, сделавших невозможным поддержание эпидемического процесса при дракункулезе.
122. Методы овогельминтоскопии. Овогельминтоскопия – это анализ, выявляющий наличие яиц гельминтов в исследуемом материале. Исследовать можно фекалии, мочу, соскобы перианальных складок, кровь, мочу или содержимое желудка. Чаще исследованию подвергаются фекалии.Яйцо печеночного сосальщика (Fasciola hepatica). Яйца печеночного сосальщика одни из самых крупных яиц гельминтов (130-150×70-90 мкм). Они имеют овальную форму и коричневую окраску. На одном из полюсов есть крышечка. Зарисуйте яйцо фасциолы в альбом, на рисунке обозначьте: 1 – крышечка.Яйцо широкого лентеца (Diphyllobothrium latum). Яйца широкого лентеца похожи на яйца печеночного сосальщика, но имеют меньшие размеры (70× 45 мкм). Яйца имеют желтоватый цвет и крышечку на одном из полюсов. Зарисуйте яйцо широкого лентеца в альбом, на рисунке обозначьте: 1 – крышечка.тениид (Taenia solium и Taeniarhynchus saginatus). Яйцо округлое, размером 0,031 - 0,038 мм. Под оболочкой яйца видна онкосфера с шестью крючьями. Яйца свиного и бычьего цепней практически не различимы. Зарисуйте яйца тениид в альбом, на рисунке обозначьте: 2 – онкосфера.Яйца шистосом(Schistosoma haematobium и Schistosoma mansoni). Яйца Schistosoma haematobium и Schistosoma mansoni овальные, размером 0,12 0,16 – 0,05-0,07 мм. Яйца Schistosoma haematobium имеют шип на одном из полюсов, яйца Schistosoma mansoni - сбоку. Зарисуйте яйца шистосом в альбом, на рисунке обозначьте: 3 – шип. Яйцо аскариды. Инвазионное яйцо аскариды желтого цвета, имеет плотную неровную (бугристую) наружную оболочку и гладкую полупрозрачную внутреннюю. Размеры 30-40×50-60 мкм. Зарисуйте яйцо аскариды в альбом, на рисунке обозначьте: 4 – бугристая оболочка.Яйцо острицы. Яйца остриц бесцветные, прозрачные, с тонкой двухслойной оболочкой. Имеют овальную форму, срезаны с одной стороны. Размер яиц – 0,05-0,06×0,02-0,03 мм. Зарисуйте яйцо острицы в альбом. Яйцо власоглава. Яйца власоглава имеет форму бочонка, на полюсах яйца имеются пробочки. Размер яйца – 0,047-0,052×0,022-0,023 мм. Зарисуйте яйцо власоглава в альбом, на рисунке обозначьте: 5 – пробочки.
123. Тип Членистоногие. Классификация. Характерные черты организации. Медицинское значение.Членистоногие - самая многочисленная группа животных населяющих нашу планету, число видов превышает 2 миллионов. В состав типа входит несколько классов, из которых основными являются ракообразные, паукообразные и насекомые. Членистоногие обладают членистым телом, в котором можно выделить 3 основные отдела: голову, грудь, брюшко. На голове располагаются органы чувств и ротовые придатки, грудь несет органы движения — членистые конечности, а у насекомых еще и крылья. Брюшко чаще всего снабжено половыми придатками. Наружный покров членистоногих — кутикула — пропитан твердым азотсодержащим полисахаридом — хитином. Образуется наружный скелет. Это создает затруднения при росте. Поэтому членистоногие линяют: старая кутикула лопается и животное выползает из нее. Рост происходит, пока новая кутикула не пропитается хитином, животное растет, увеличиваясь в размерах. Тело типичного членистоногого состоит из нескольких сегментов, покрытых твердым наружным скелетом (экзоскелетом). Мышечная система очень сложна и образована большим количеством специализированных мышц. Пищеварительная система сложно дифференцирована в продольном направлении. Дыхательная система в зависимости от образа жизни, представлена жабрами, легкими или трахеями. Кровеносная система всегда незамкнутая. Нервная система состоит из сложноустроенного мозга и типичной брюшной нервной цепочки. Все членистоногие раздельнополые. Развитие членистоногих происходит как с превращением, так и прямым путем. Прямое развитие свойственно всем паукообразным. Насекомые преимущественно развиваются с превращением, причем встречаются различные формы этого процесса. Трансмиссивные болезни, инфекционные и паразитарные заболевания человека и животных, возбудители которых передаются членистоногими. Передача возбудителя происходит при укусе комарами, блохами, москитами, клещами и др., при попадании на кожу и слизистые оболочки инфицированных выделений переносчика и др. путями. У человека различают облигатные Т. б., возбудители которых передаются исключительно переносчиками (малярия, жёлтая лихорадка, клещевой возвратный тиф и др.), и факультативные Т. б., передача возбудителей которых осуществляется воздушно-капельным путём, через пищеварительный тракт, непосредственно от человека к человеку (туляремия, чума, сибирская язва и др.). Т. б. относится к болезням с выраженной природной очаговостью.
124. Класс Паукообразные. Классификация. Характерные черты организации. Медицинское значение.Паукообра́зные (лат. Arachnida) — класс членистоногих из подтипа хелицеровых (Chelicerata). Наиболее известные представители: пауки, скорпионы, клещи́. Размеры паукообразных варьируют от сотен микрон (некоторые клещи) до нескольких сантиметров. Длина тела аранеоморфных пауков и сенокосцев обычно не превышает 2—3 см. Наиболее крупные представители класса (скорпионы, сольпуги и жгутоногие) могут достигать 20 см в длину. Ещё большими размерами обладают некоторые пауки-птицееды. Традиционно в теле паукообразных выделяют два отдела — головогрудь и брюшко. Головогруди состоит из 6 сегментов, несущих по паре конечностей: хелицеры, педипальпы и четыре пары ходных ног. У паукообразных они несут относительно тонкую хитиновую кутикулу, Кутикула предохраняет организм от потери влаги при испарении.Органами дыхания служат трахеи (у фаланг, лжескорпионов, сенокосцев и некоторых клещей) или так называемые легочные мешки (у скорпионов и жгутоногих), иногда те и другие вместе (у пауков);Пищеварительная система приспособлена к питанию полужидкой пищей.Кишечник состоит из узкого пищевода, принимающего слюнные железы, желудка, снабженного парными и непарными отростками, и задней кишки, обыкновенно с расширенной клоакой, впереди которой впадают выделительные, так называемые мальпигиевы железы.Существуют и другие выделительные органы, так называемые коксальные железы.Органы выделения —мальпигиевы сосуды.Органы чувств разнообразны: простые глаза, органы обоняния, вкуса, осязания, химического чувства и др.Все паукообразные раздельнополы и в большинстве случаев демонстрируют ярко выраженный половой диморфизм. Половые отверстия располагаются на втором сегменте брюшка (VIII сегмент тела). Большинство откладывает яйца, но некоторые отрядыживородящи (скорпионы, бихорхи, жгутоногие).Некоторые отряды имеют специальные органы.ядоносный аппарат — скорпионы и пауки.прядильный аппарат — пауки и лжескорпионы.Скорпионы — крупные хищники (до 15 см длиной), напоминающие по внешнему виду речного рака и имеющие клешневидную вторую пару ротовых конечностей. Брюшко вытянуто и подвижно. В позе угрозы скорпион заносит его над головой и вонзает ядовитую иглу, расположенную на конце последнего сегмента брюшка, в тело жертвы, удерживая ее клешнями. Питаясь насекомыми, пауки и скорпионы снижают численность вредителей. Сами они, в свою очередь, служат пищей для мелких ящериц, птиц и млекопитающих.Клещи — мелкие, в том числе микроскопические, наземные членистоногие. Многие из них являются паразитами животных и растений. Для клещей характерно слияние отделов тела. Ротовые конечности у паразитических форм видоизменены в колющий хоботок, с помощью которого клещи внедряются в тело хозяина. Так, мелкие паутинные клещи высасывают соки из листьев картофеля, малины, земляники, огурцов. Амбарные клещи —- серьезные вредители зерна и муки. Чесоточный клещ просверливает ходы в коже человека и вызывает мучительную болезнь — чесотку. Сотни видов клещей — паразиты птиц и млекопитающих.
125. Клещи. Систематическое положение, семейства, морфология, особенности развития, медицинское значение. Отряд Клещи Acari Представители этого отряда имеют несегментированное тело. Ротовой аппарат представлен сложно устроенным хоботком, в состав которого входят хелицеры и дополнительное образование — гипостом. Педипальты выполняют обонятельную осязательную функции. Развитие клещей происходит с метаморфозой. Из яйца вылупляется личинка, имеющая три пары ног. За ней следует стадия нимфы. У нее по четыре пары конечностей, но половая система не развита. Нимфа превращается во взрослую стадию — имаго, отличающая более крупными размерами и развитой половой системой. Некоторые виды имеют несколько стадий нимфы.У клещей - временно кровососущих эктопаразитов ротовой аппарат предназначен для прокалывания кожи и всасывания крови, пищеварительная система и покровы тела сильно растяжимы, что позволяет им питаться редко, иногда только один раз в жизни. Выделяемая в ранку слюна клещей обладает местным раздражающим и общетоксическим действием. Примеры: иксодовый (таежный, собачий, пастбищный), аргазовые, гамазовые клещи.Один из путей борьбы — использование животных, паразитирующих на клещах и на яйцах, вызывающих их гибель или снижающих жизнеспособность. У клещей — обитателей человеческого жилища ротовой аппарат грызущего типа. Способны к активным передвижениям в поисках пищи. Питаются пищевыми запасами — зерно, мукой, копченым мясом и рыбой, сушеными овощами и фруктами, а также эпидермальными чешуйками, сливающимися с поверхности кожи человека, и спорами плесневых грибов. Опасность для здоровья человека: 1 портят пищевые продукты, 2 кусают человека, вызывая зерновую чесотку, чесотку продавцов бакалейных товаров и др. С продуктами питания могут попадать в пищевую систему человека, вызывая тошноту, рвоту, понос. Некоторые виды клещей способны переходить в анаэробных условиях кишечника и даже размножаться там. При попаданию с пылью в дыхательные пути и легкие эти клещи вызывают акаридоз дыхательной системы. Примеры: домашний клещ.Борьба с домовыми клещами состоит в первую очередь в частых влажных уборках помещений, использование мебели, подушек и матрацев из синтетических материалов, в которых эти животные не могут поселяться. У клещей — постоянных паразитов человека крайне мелкие размеры тела — 0,1 — 0,4 мм, сильная редукция конечностей, малая подвижность и цикл развития, полностью проходящей на хозяине. К ним относятся чесоточный зудень - возбудитель чесотки человека. Чесотка может поражать чесоточные зудни лошадей, коз, верблюдов, собак и др. животных. Они вызывают характерные поражения кожи, но на человеке живут не долго и почти не размножаются. Перемещаясь в толщи кожи, клещи раздражают нервные окончания и вызывают нестерпимый зуд, усиливающийся ночью. При расчесывания ходов клещей они переносятся на др. участки кожи или на др. человека. Железница угревая — возбудитель демодикоза. Эти клещи обитают в сальных железах и волосяных фолликулах кожи лица, шеи и плеч, располагаясь головным концом вниз, обычно группами по 4 особи. Массовые нападения клещей могут быть причиной не только поражений кожи — клещевых дерматитов, но и тяжелых лихорадочных состояний и нервных расстройств. Особенную опасность представляет не сколько питание кровью, способность быть переносчиком возбудителей пригодно — очаговых протозойных, бактериальных, вирусных болезней, а также заболеваний, вызываемых риккетсиями.
126. Клacc насекомые. Классификация. Характерные черты организации. На сегодняшний день известно около 1 млн видов насекомых, но истинное их количество может быть не менее 2 млн.Кратко класс можно охарактеризовать следующим образом. Это наземные членистоногие, у которых тело явно разделено на голову, грудь и брюшко, а основные, служащие для передвижения, конечности находятся в числе 3 пар на грудном отделе. Брюшко у взрослых особей несёт видоизменённые остатки конечностей — грифельки и др., лишь у отряда бессяжковых сохранились слабые брюшные ноги. Дыхание осуществляется с помощью трахейной системы или кожное — всей поверхностью тела. Те насекомые, которые дышат с помощью всей поверхности тела и имеют проницаемые для дыхательных газов и испарения покровы, ограничены в своём распространении только влажными средами. Они встречаются в основном в почве и в гниющих остатках организмов. Те высшие насекомые, у которых покровы непроницаемы и развита трахейная система, могут обитать и в сухих средах. Эти насекомые широко расселились по нашей планете. Именно такие, ведущие открытый образ жизни и часто ярко окрашенные высшие насекомые и известны большинству читателей. Размеры тела 0,2—330 мм (у современных представителей), обычно в пределах 1—50 мм. Покрывающее тело кутикула является частью кожи, образуя плотный наружный панцирь или экзоскелет, но в ряде случаев она мягкая и тонкая. Наружная кутикула подразделена на отдельные щитки — склериты (sclerites) и вследствие своей плотности благоприятна для развития на ней различных образований: вдавлений, бороздок, бугорков, рёбрышек, мелких волосков -хетоидов и др. К кутикуле причленяются также подвижные кожные волоски — хеты, имеющие иногда характер щетинок или чешуек. Тело насекомых подразделяется на три сегментированных отдела: голова, грудь и брюшко. Каждый сегмент подразделяется на четыре части — верхнее полукольцо называется тергит, нижнее полукольцо называется стернит, боковые стенки — плейриты. Усики сидят по бокам лба, между глазами или впереди них, нередко в хорошо обособленной усиковой впадине. Они очень разнообразны, характерны для разных групп насекомых. В своей основе усики состоят из утолщённого основного членика, называемого рукояткой (scapus), за которым следует ножка (pedicellus), и, начиная с третьего членика, располагается основная часть — жгутик (flagellum). Различают несколько типов усиков. Крылья насекомых (alae) представляют собой пластинчатые выросты покровов, связанные с комплексом обслуживающих их мышц и преобразованных склеритов груди. В типичном случае их две пары: передняя, прикреплённая к среднегруди, и задняя — на заднегруди. Крылья состоят из тонкой крыловой пластинки (membrana), укреплённой на твёрдых жилках (venae), играющих роль прочной основы крыла. Дыхательная система большинства насекомых представлена множеством трахей, пронизывающих всё тело. Трахейная система насекомых выполняет не только роль снабжения организма кислородом, но и транспортную функцию по доставке кислорода к тканям. Необходимость наличия трахейной системы, пронизывающей всё тело, не позволяет насекомым достигать крупных размеров. Кровеносная система насекомых незамкнутая. Гемолимфа свободно циркулирует по полости тела, её приводит в движение мускулистая трубка, разделённая на камеры, — сердце. Число камер — до восьми, каждая камера имеет пару боковых отверстий с клапанами, обеспечивающих односторонний ток крови из полости в сердце. Нервная система построена по типу брюшной цепочки. У высших отрядов насекомых нервная система достигает высокого уровня дифференциации, имеется относительно развитый головной мозг.
127. Комнатная муха, муха це-це, Вольфартова муха. Систематическое положение; морфология; эпидемиологическое значение, меры борьбы.Мухи (лат. Musca) — род насекомых семейства Настоящие мухи (Muscidae). Наиболее распространённые виды: комнатная муха (Musca domestica Linnaeus, 1758), домовая муха (Muscina stabulans), Зелёная падальная муха (Lucilia caezar L.) и Серая мясная муха (Sarkophaga carnaria L.)Мухи являются переносчиками некоторых видов опасных заболеваний, например сибирской язвы и сепсиса (переносчик Осенняя жигалка (Stomoxys calcitrans Linnaeus, 1758)).Мухи - одни из самых известных насекомых, часто сталкиваются с человеком. Мухи - одна из неприятностей "летних" месяцев.Мухи всех видов рассматриваются в символическом аспекте негативно. Зачастую господствовали представления о демонической мощи, воплощенной в неистребимых полчищах мух. Изображения мух на античных геммах должны были защитить от сглаза. Её греческое название символически означает паразита.Мухи настоящие (Muscidae) чрезвычайно разнообразны и населяют почти всю Землю от полярных стран до экватора. Многие из них широко распространены, как, например, комнатная муха, мясная муха, золотисто-зелёная муха, которые встречаются летом во всех тех местах, где есть органические гниющие вещества и вообще нечистоты.Известно около 4 тыс. видов настоящих мух. Питаются соком растений, иногда – кровососы. Личинки развиваются в органических остатках. Многие настоящие мухи – переносчики опасных заболеваний.Мухи комнатные (Musca domestika) относится к отряду двукрылых. Она не может переваривать твёрдые куски, поэтому питается в основном жидкой пищей. Однако на хоботке мухи имеются крошечные зубы, которыми она может отделять мельчайшие частицы пищи, которые потом размягчаются жидкостью, отрыгиваемой через хоботок.У большинства мух имеется пара так называемых настоящих крыльев, с помощью которых она может летать. Они чрезвычайно быстры в полёте, манёвренны и неуловимы. Кроме них у мухи есть ещё пара задних, или ложных, крыльев, именуемых жужжальцами, которые помогают мухе удерживать равновесие в полёте, крутится в воздухе на одном месте и даже летать задом наперёд.Комнатная муха распространилась по всему миру вместе с человеком, и не только досаждает ему, но и прямо угрожает его здоровью. Она размножается в органическом веществе, в т.ч. и в экскрементах, а взрослые мухи питаются без разбора как этим, так и сладкой пищей. Мухи приносят на своих конечностях возбудителей болезней, заражая ими пищу. Кроме того, во время питания они отрыгивают слюну и частично переваренную пищу, чтобы растворить сахар и другую плотную пищу. Эта отрыгнутая жидкость содержит множество вредных для человека микроорганизмов, которых муха заглотила вместе с загрязнённой пищей.Мухи падальные зелёные (Lucilia) – род падальных, или мясных, мух. Типичные обитатели трупов. Откладывают единовременно до 200 яиц, поглубже в тело трупа, в защищённые от света места, складки. Через сутки из них вылупляются личинки. Они питаются мёртвым телом и постепенно развиваются. Продолжительность развития зависит от температуры воздуха и составляет от 10 до 150 суток.Мухи падальные синие (Calliphora vicina)- в длину достигают 8-13 мм. Подобно другим мухам, она очень плодовита и в течение 15 мин может отложить 60-70 яичек, обыкновенно кучками, а иногда – до 100 вместе. Их яйца способны развиваться как на падали, так и в экскрементах животных и людей. Синие падальные мухи стремятся залететь в наши дома и заселить своими личинками незакрытое нами мясо.Мухи серые мясные (Sarcophagidae) – это пепельно-серая муха с бурыми полосками на спине и серебристым отливом, украшенном шашечным рисунком. Её научное имя«саркофага» обозначает «пожирательница мяса», но обычно её называют серой падальной мухой. Длинна тела 3-18 мм. Известно около 2,5 тысяч видов. Распространены широко. Для этих мух характерно живорождение или яйцеживорождение. Стоит ей на несколько секунд оказаться на куске мяса на вашей кухне или у раны не теле домашнего скота, и раз-два, кучка крохотных личинок уже копошится и расползается, проникая внутрь.Запас яиц «саркофаги» достигает двадцати тысяч. В один приём появляется до 30 личинок, но саркофага рождает их много раз. Через 4-8недель готова новая поросль мух.Мушки плодовые, дрозофилы(Drosophilidae) – одно из семейств двукрылых. Быстро развиваются, возможно, развитие в запасах фруктов и овощей, комнатных вазонах, кадках и горшках; через 7-10 дней появляется молодая, назойливая поросль.
128. Вши, блохи, систематическое положение, морфология, особенности развития, эпидемиологическое значение, методы борьбы.Вши (Класс Насекомые). К постоянным кровососущим паразитам человека из класса насекомых относятся только вши. Человек для них - единственный хозяин, поэтому и трансмиссивные заболевания, возбудителей которых переносят вши, являются типичными антропонозами.Вши характеризуются выраженными адаптациями к эктопаразитизму: размеры их невелики, конечности снабжены аппаратом фиксации к коже, волосам и одежде, ротовой аппарат колюще-сосущего типа, цикл развития упрощен (развитие с неполным метаморфозом), все стадии жизненного цикла обитают и питаются на хозяине. Человеческая и лобковая. Вид человеческая вошь представлен двумя подвидами: головная, платяная вошь. Зараженность лобковыми вшами - фтириаз.Вши отличаются друг от друга морфологическими и физиологическими признаками и особенностями жизненного цикла. Самая крупная вошь — платяная, размеры до 4,7 мм. Головная достигает длины 3 мм, а лобковая—не более 1,5 мм. Платяная и головная вши имеют четко отграниченные друг от друга головку, грудь и брюшко, а у лобковой грудь и брюшко слиты. Платяная вошь живет около 50, головная — около 40, а лобковая —до 30 сут. Головная и платяная вши питаются кровью по 2—3 раза в сутки, а лобковая —почти постоянно малыми порциями. Самка платяной и головной вшей откладывает до 300 яиц за всю жизнь, лобковой —до 50. Яйца приклеиваются к волосам или нитям одежды- гниды, очень устойчивы.Слюна вшей обладает токсическими свойствами, вызывает ощущение жжения и зуда. Лобковая вошь является только паразитом, а головная и платяная — еще и специфическими переносчиками спирохет - возбудителей возвратного тифа, риккетсий - возбудителей эпидемического сыпного тифа, возбудителей волынской лихорадки.Профилактика - борьба с педикулезом, соблюдение правил личной гигиены, особенно в местах массового скопления людей. Из химических средств используют мази и шампуни, содержащие инсектициды, лекарственные препараты, применяемые внутрь и действующие через кровь, которой питаются эти паразиты. Наиболее эффективны короткая стрижка для уничтожения гнид и обработка белья в дезинфекционных камерах.Блохи (Класс Насекомые, Отряд Блохи)— это временно сосущие паразиты, мелкие насекомые длиной от 1 до 5 мм. Паразитирование блох облегчается сплющенностью тела с боков, наличием на поверхности его большого количества щетинок, направленных остриями назад, и колюще-сосущим ротовым аппаратом. Задние конечности удлинены и служат для передвижения прыжками. Признак дегенерации- рудиментарные глаза и отсутствие крыльев. Развитие блох идет с полным метаморфозом.Человеческая блоха и крысиная блоха предпочитают питаться кровью соответственно человека и крыс, легко переходят на другие виды животных. Крысиная блоха живет в норах крыс, человеческая — в трещинах пола, за плинтусами и обоями. Здесь самки откладывают яйца, из которых развиваются червеобразные личинки, питающиеся разлагающимися органическими веществами, фекалиями взрослых блох. Через 3—4 недели они окукливаются и превращаются в половозрелых насекомых.Человека блохи посещают ночью. Укусы болезненны, вызывают сильный зуд. Блохи- переносчики бактерий — возбудителей чумы. Борьба с блохами — содержание жилых помещений и хозяйственных построек в чистоте, применение инсектицидов и различных средств борьбы с грызунами. Меры индивидуальной защиты, например репелленты, которыми пропитывают одежду и постельное белье.
129. Комары. Систематическое положение, строение, развитие, медицинское значение, меры борьбы. Комары малярийные и немалярийные. Комары — насекомые с тонким телом (длиной 4—11 мм), длинными ногами и узкими прозрачными крыльями. Окраска тела жёлтая, коричневая или серая. Брюшко удлинённое, состоящее из 10 сегментов. Грудь шире брюшка. Лапки заканчиваются парой коготков. Крылья покрыты чешуйками, скопления которых иногда образуют пятна. Антенны длинные, состоят из 15 члеников. Ротовой аппарат колюще-сосущего типа. У самок хоботок длинный и состоит из колющих щетинок, у самцов — без них. Как и у всех прочих двукрылых насекомых, у комаров 4 фазы развития: яйцо, личинка, куколка, имаго. При этом все фазы, кроме имаго, живут в водоёмах. Живущие в воде личинки и куколки комаров дышат атмосферным воздухом через дыхательные трубки, выставляя их на поверхность. Летом взрослые самки кровососущих комаров встречаются как в природе в заболоченных и сырых местах, так и в помещениях для животных, в жилище человека на стенах, окнах, в затенённых местах. Зимой их можно встретить в помещениях для скота, тёплых подвалах, других постройках, где они пребывают в малоактивном состоянии, либо в оцепенении (если температура ниже 0°C). При выборе жертвы самка кровососущего комара ориентируется на углекислый газ, выдыхаемый человеком (радиус обнаружения до 12 метров[источник не указан 128 дней]), на тепловое излучение, а также на запах молочной кислоты, содержащейся в поте. Также самка комара реагирует на свет, предпочитая освещённые помещения.
130. Москиты. Систематическое положение, строение, развитие, медицинское значение, меры борьбы. Моски́ты (Phlebotomidae)кровососущие двукрылые насекомые семейства Phlebotomidae; входят в состав гнуса (Гнусавость).Семейство насчитывает свыше 400 видов, сгруппированных в 4 рода: Phlebotomus, Sergentomyia, Lutzomyia, Brumptomyia. Москиты родов Phlebotomus и Sergentomyia распространены на юге Украины, в Молдове, на Кавказе, в республиках Средней Азии; условная северная граница их ареала — 47—48° с. ш.Москиты наносят большой вред человеку, т.к. являются активными кровососами и переносчиками возбудителей некоторых заразных болезней: флеботомной лихорадки (Флеботомная лихорадка), кожного и висцерального лейшманиозов (Лейшманиозы), бартонеллеза. Основной переносчик москитной лихорадки на территории СССР — Ph. papatasii, переносчики кожного лейшманиоза — Ph. papatasii, Ph. sergenti, Ph. caucasicus, переносчики висцерального лейшманиоза — Ph. chinensis и Ph. kandelakii.Укусы М. болезненны. В месте укуса образуется папула или волдырь, появляются зуд и жжение. В отдельных случаях отмечается повышение температуры тела.Взрослые особи имеют длину 1,3—3,5 мм (рис.); окраска их варьирует от светло-желтой до темно-серой. М. — насекомые с полным превращением (см. Насекомые).Преимагинальные фазы (яйцо, личинка, куколка) развиваются во влажном разлагающемся органическом субстрате. Местами выплода М. в природных биотопах являются колонии большой и краснохвостой песчанок, норы сусликов, дикобразов и др. В районах с относительно высокой влажностью выплод М. происходит также в гнездах некоторых видов птиц (сизоворонки, золотистой щурки); в населенных пунктах — в норах домовых грызунов, в подпольях, в кучах мусора и др.После выхода из яйца личинка питается органическими остатками субстрата, на котором были отложены яйца. Личинка проходит в своем развитии четыре стадии и после линьки превращается в куколку. Окукливание осуществляется на поверхности субстрата. Продолжительность развития от яйца до имаго при температуре около 25° в зависимости от вида — 45—60 дней. На 2—3-й день после вылета из куколки самки ищут объекты для кровососания. В отличие от большинства других кровососущих двукрылых М. не разлетаются далеко от места выплода. Питание продолжается 1—2 мин, однако вспугнутая самка может прервать кровососание и перелететь на другой объект для продолжения питания. Эта способность к прерывистому питанию на различных объектах имеет большое эпидемиологическое значение, т.к. увеличивает вероятность передачи возбудителей.Москиты — временные свободноживущие кровососы. Кровь пьют только самки, подчиняющиеся закону гонотрофической гармонии, согласно которому яйцекладке предшествует акт кровососания. Лет и нападение на добычу происходят перед заходом солнца и в первые часы после заката. В Средней Азии и Закавказье М. появляются в апреле — мае и исчезают в сентябре — октябре. За сезон дают 1—3, чаще 2 генерации.Меры борьбы с М. направлены главным образом на уничтожение взрослых особей. В населенных пунктах обработку помещений и предполагаемых мест выплода проводят контактными инсектицидами (см. Дезинсекция). Если в данной местности бо́льшее эпидемиологическое значение имеют москиты экзофильных видов, то обрабатывают наружные поверхности стен, кучи мусора, органических субстратов и пр. При наличии М. эндофильных видов основное внимание обращают на обработку внутренних поверхностей жилых и подсобных помещений. Одновременно следует проводить санитарное благоустройство поселков и окружающей их территории (истребление грызунов, заделывание нор, очистка). Защита людей от укусов М. состоит в засетчивании окон и дверей, применении пологов, а также репеллентов
131. Класс Ракообразные, иx участие в качестве промежуточных хозяев гельминтов.Общая характеристика класса. Класс включает более 40 тыс. водных членистоногих, большинство которых обитает в морях, реже в пресных водоемах и лишь немногие из них освоили влажные наземные местообитания. Образ жизни плавающий, ползающий, реже прикрепленный; имеются паразитические формы. Несмотря на различное количество сегментов тела (от 10 до 50) и разнообразие внешней формы, организация ракообразных имеет следующие сходные черты: 1) дыхание с помощью жабр; 2) слияние головного и грудного отделов с образованием головогруди; 3) наличие двух пар усиков, выполняющих осязательную и обонятельную функции, пары сложных, или фасеточных, глаз, и трех пар ротовых конечностей (пара верхних и две пары нижних челюстей, которые захватывают и измельчают пищу); 4) разнообразное строение грудных конечностей, которые выполняют функции удержания и перемещения пищи ко рту, движения организма, дыхания; 5) брюшные конечности служат для плавания, а у самок и для прикрепления оплодотворенных яиц; 6) ракообразные всех возрастных групп линяют, но молодь чаще, чем взрослые.Не все ракообразные являются промежуточными хозяевами гельминтов. Веслоногие являются промежуточными хозяевами гельминтов:1. промежуточными хозяевами ришты (Dracunculus medinensis) служат различные циклопы (род Cyclops и другие представители Cyclopoidea). В настоящее время ришта на территории России почти истреблена (проф. Л. М. Исаев);2. первыми промежуточными хозяевами широкого лентеца (Diphyllo-bothrium latum (L.), а также, по-видимому, и других видов лентецов человека, распространенных в Сибири и на Дальнем Востоке, является целый ряд различных представителей циклопов (роды Acanthocyclops, Cyclops, Eucy clops, Macrocyclops, Mesocyclops, Tropocyclops), а также некоторые виды рода Diaptomus (Calanoidea): D. gracillis, D. graciloides и др.